
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Муниципальное образовательное учреждение «Средняя школа с углубленным изучением отдельных предметов №49 Краснооктябрьского района Волгограда»
Исследование полиморфизма листоеда ильмового (Galerucella luteola Mull)
Волгоград, 2016 |
СОДЕРЖАНИЕ
ВВЕДЕНИЕ………………………………………………………………….……3
ГЛАВА 1. ПОЛИМОРФИЗМ ПОПУЛЯЦИЙ……………………………….….5
ГЛАВА 2. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ………………….….9
2.1. Материал исследования..................................................................................9
2.2. Листоед ильмовый как объект исследования……………………………..10
ГЛАВА 3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ………………………………...15
3.1. Особенности морфологии листоеда ильмового по рисунку переднеспинки…………………………………………………………….……..15
3.2. Особенности полиморфизма исследуемых популяций листоеда ильмового………………………………………………………………………...19
ВЫВОДЫ ………………………………………………………………………..24
ЛИТЕРАТУРА…………………………………………………………………...25
ПРИЛОЖЕНИЕ………………………………………………………………….28
ВВЕДЕНИЕ
Стратегия выживания любого организма выражается в проявлении приспособительных процессов, которые возникают из-за постоянно меняющихся характеристик онтогенеза и под влиянием изменения экологических факторов. Организм, как открытая система, объективно реагирует на изменение условий обитания. Вариабельность отдельных признаков организмов отражает результат взаимодействия генетических детерминантов и условий существования (Яблоков, 1987).
На популяционном уровне одним из проявлений приспособительных процессов является полиморфизм популяций. Анализ популяционно-фенетической структуры видов и ее пространственно-временной динамики представляет собой одно из важных направлений в исследовании микроэволюционных процессов.
Начальный этап эволюционного процесса заключается в необратимом преобразовании популяции. Отсюда следует, что оценка межпопуляционных различий дает материал для суждения о том, в чем конкретно проявляется первый шаг эволюционной дифференциации вида, дает представление о масштабе внутривидовых преобразований и о формах их проявлений. Путем сравнения характера изменчивости возможно установление различий между разными популяциями, выявление особенностей внутривидовой изменчивости и закономерностей формирования адаптивных реакций на популяционном уровне. Изучение диапазона изменчивости признаков в популяциях способствует выбору критериев для выявления естественных популяционных группировок на сплошном участке видового ареала, позволяет определить размер популяций, судить о характере внутрипопуляционных группировок. Накопление и анализ данных по морфологической дивергенции популяций, находящихся в разных экологических условиях, в перспективе помогут понять адаптивную ценность тех или иных признаков для конкретной популяции, а также направления формирования популяционной структуры вида.
Кроме теоретического значения исследование характера изменчивости и полиморфизма популяций может иметь и практический интерес. Полиморфная структура популяции имеет адаптивный характер и позволяет оценить состояние популяции и перспективы ее развития – динамику численности и структуры. Это особенно важно при изучении видов, влияющих на жизнедеятельность человека и среду обитания человека, видов, причиняющих вред сельскохозяйственным культурам и животным и др. Одним из таких видов является листоед ильмовый – жук, регулярно дающий массовые вспышки численности и повреждающий ильмовые породы, широко распространенные в Нижнем Поволжье. Рекогносцировочный надзор и мониторинг, осуществляемый за данным видом в г. Волгограде и области, может использовать и фенетический подход. Это определило тему нашего исследования.
Цель исследования: изучение полиморфизма популяций листоеда ильмового (Galerucella luteola Mull)
Задачи:
1. Изучить спектр изменчивости рисунка переднеспинки листоеда ильмового.
2.Описать полиморфную структуру исследуемых популяций листоеда ильмового.
3.Провести сравнительный анализ полиморфной структуры исследуемых популяций листоеда ильмового.
Объект исследования: полиморфизм популяций листоеда ильмового (Galerucella luteola Mull)
Предмет исследования: особи листоеда ильмового.
Гипотеза: Рисунок переднеспинки листоеда ильмового – полиморфный признак, который свидетельствует о характере действии естественного отбора.
ГЛАВА 1. ПОЛИМОРФИЗМ ПОПУЛЯЦИЙ
Полиморфизм (от греч. Polymorphos – многообразный) – наличие в пределах одного вида резко отличных по облику особей, не имеющих переходных форм.
Полиморфизм – категория внутрипопуляционная. Полиморфизм можно определить как дискретную внутрипопуляционную изменчивость наследственного характера (Новоженов,1979).
Полиморфизм носит адаптивный характер и его значение связано с наличием в популяции отличающихся форм, которые делают ее пластичной (Кайданов, 1996). Эти формы насыщают популяцию генетическим материалом, который при изменении давления отбора (его направленности и интенсивности) позволяет ей быстро и успешно изменить свою структуру и приспособиться к новым условиям среды. Большая изменчивость у многих насекомых с разными типами покровительственной окраски имеет индивидуальную адаптивность, т. к. не позволяет хищнику выработать условный рефлекс на определенный раздражитель (Новоженов, 1997).
В отличие от мутаций полиморфизм – это отобранная изменчивость. В популяции даже редкие формы встречаются с определённой частотой, более или менее стабильной из поколения в поколение. Это является доказательством того, что они включены отбором в гомеостатическую генетическую систему популяций, а частота их соответствует степени необходимости. В пределах всех популяций имеется достаточно богатое разнообразие вариации. Однако в каждой из них, как правило, преобладает одна типичная форма, сопровождаемая двумя, тремя дополнительными и серией редких форм (Новоженов,1979).
Полиморфизм популяций – это понятие было определено в 1940 году английским генетиком и экологом Э. Фордом как существование в популяции одновременно двух или более генетически различающихся форм, причём частота наиболее редкой формы всё же достаточно велика, чтобы её поддержание можно было объяснить мутационным давлением. В популяционно-генетической литературе принято считать критерием полиморфизма присутствие в популяции наиболее редкой формы частотой >5%. В некоторых случаях этот порог снижают до 1%.
Отличие генетической гетерогенности от генетического полиморфизма популяций состоит в том, что генетическая гетерогенность – это любое, пусть даже самое минимальное генетическое разнообразие в популяции, вызываемое мутационным процессом или рекомбинантным процессом (Береговой, 1977). Полиморфизм, как следует из того, что у данного определения есть устойчивое поддержание в популяции достаточно многих генотипических классов. Его поддержание может быть лишь при участии отбора.
Различают полиморфизм по разным признакам: морфологическим, физиологическим, биохимическим. Выделяют адаптивный, геномный, хромосомный, гетерозиготный, инверсионный и др. полиморфизм.
Неоднородность генотипа и фенотипа особей, входящих в популяцию имеет большое экологическое значение. В зависимости от конкретных условий больше шансов выжить то у одних, то у других особей, которые затем снова восстанавливают всю популяцию. Так, например колорадский жук способен быстро наращивать свою численность даже после воздействия очень неблагоприятных условий: сплошных химических обработок или временного отсутствия своего кормового растения, благодаря разной продолжительности диапаузы у разных особей. Диапаузирующие жуки могут находиться в почве от одного года до трех лет.
Иногда эта изменчивость может регулироваться внешними факторами, как бы направляющими особь по тому или иному пути развития. Так под влиянием большой плотности популяции саранча Locusta migratoria L. существенно меняет свой облик и физиологию, переходя из одиночной фазы в мигрирующую. Мигрирующая саранча охотно образует скопления – кулиги личинок и стаи имаго, передвигающиеся в поисках корма. Плодовитость мигрирующей саранчи в 3-4 раза ниже, чем одиночной.
Некоторую аналогию с мигрирующей саранчой представляют крылатые особи тлей, возникающие, в частности, при высокой скученности тлей на растении или ухудшении питания. Правда, крылатые тли, не способные к активному полету из-за своих малых размеров, мигрируют одиночно, не образуя скоплений. Под влиянием условий меняется соотношение длинно и короткокрылых особей одного вида клопов-водомерок. Жаркое и сухое лето способствует появлению большого количества длиннокрылых мигрирующих особей, что необходимо в связи с массовым пересыханием мелких водоемов. Короткокрылые же особи оказываются более устойчивыми к неблагоприятным условиям и могут вытеснять длиннокрылых.
Полиморфизм может быть обусловлен и чисто генетическими причинами. Так -Ресовский в течение 11 лет (1930-1940) исследовал полиморфизм по окраске надкрыльев в популяции божьей коровки Adelia bipunctata в предместье Берлина-Бухе. Дважды в год, весной и осенью, он определял частоту встречаемости двух морф – красной и чёрной. В результате проведенных исследований -Ресовский пришёл к выводу, то популяционная динамика двух форм, очевидно, зависит от различий в их адаптивных качествах в разные сезоны года. Чёрные коровки плохо переносят зимовку, что, по-видимому, связано с повышенной отдачей тепла при такой окраске, но зато интенсивнее, чем красные, размножаются в летние месяцы, благодаря чему в природных популяциях поддерживается относительно стабильное равновесие. Таким образом, особи – обладатели разных фенов – всегда обнаруживают те или иные адаптивные различия (Яблоков, 1980). Так, разнонаправленное давление отбора в отдельные периоды жизни популяции приводит к выработке устойчивого адаптационного полиморфизма (Яблоков, 1998). В результате такой стратегии отбора выигрывает вся популяция в целом.
Проявление индустриального меланизма на протяжении полутора веков удалось наблюдать в популяциях бабочек березовых пядениц (Biston betularia) в Англии. К середине 19 в. частота чёрных пядениц в районе Манчестера не превышала 1 %, но уже к концу века эта форма в значительной степени вытеснила светлую форму в промышленно загрязненных районах. За последнюю четверть 20 в. наблюдался обратный процесс – постепенное уменьшение доли меланистов и возрастание светлых форм в популяции бабочек. Этот процесс обусловлен, очевидно, эффективными мерами по охране окружающей среды в Западной Европе.
Собранные данные и их противоречия по модельным объектам: Adalia bipunctata, Harmonia axyridis позволили выявить эволюционное значение полиморфизма, как фактора популяционной адаптивности, пластичности и полифункциональности (Тимофеев-Ресовский, 1979).
Изменчивость в популяции может быть и не так очевидна, но иметь большое приспособительное значение. В любой популяции можно найти более и менее склонных к миграции особей. В зависимости от запасов пищи и погоды преимущественно выживает та или иная часть популяции, сохраняя ее общий генофонд. Кроме того, в популяции всегда выделяются менее разборчивые к питанию или другим условиям особи и особи, более специализированные по своим потребностям и возможностям (Креславский, 1987).
Итак, полиморфизм есть та смешанная эволюционная стратегия популяции, которая делает ее оптимально приспособленной. Реакция популяции характеризуется двумя тактиками: устойчивостью и изменчивостью. Возможности и той, и другой определяются популяционным гомеостазом. Даже значительные колебания в частоте составляющих форм не нарушают сложившуюся систему гомеостаза (Новоженов, 1989).
ГЛАВА 2. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
2.1. Материал исследования
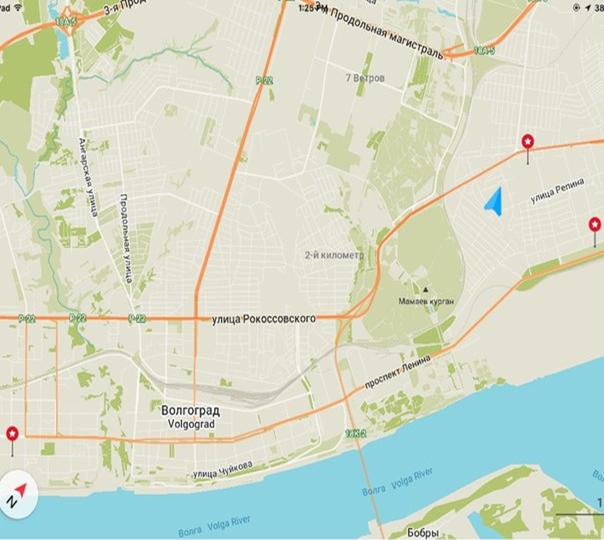
Материалом для работы стали сборы листоеда ильмового, проведенные в 2015 г. на трех участках, два из которых расположены в Краснооктябрьском районе (пр. Ленина рядом с заводом Красный Октябрь и ул. Маршала Еременко рядом с крупной автомагистралью) и один в Ворошиловском районе (ул. Баррикадная) г. Волгограда.
При выборе мест сбора учитывалась удалённость их друг от друга, что, вероятно, позволяет говорить о сборе жуков в разных популяциях.
Сбор насекомых на разных участках производился с интервалом в 3-4 дня, то есть жуки принадлежат к одному сроку вылета. Сбор проводился руками путём случайной выборки жуков из популяции. Всего было собрано 120 жуков.
При описании фенов мы использовали метод непозиционного кодирования (Яблоков, 1981). Описывали вариации каждого элемента рисунка. Пятна на переднеспинке очень изменчивы, поэтому подобные фены объединяли в комплексы – морфы (Яблоков, 1985).
При характеристике фенооблика популяций и межпопуляционном сравнительном анализе учитывали число и набор морф, частоту встречаемости каждой из них. Установленное соотношение морф позволило выделить доминантные, сопутствующие, фоновые и редкие формы (Степанчук, 2009).
Под доминатной морфой понимают такую морфу, частота встречаемости которой в данной популяции наибольшая по сравнению с другими морфами.
Сопутствующими считают морфы, частота которых менее чем в 2 раза ниже доминантной.
Фоновые морфы, имеют частоту меньше сопутствующих, но больше редких. Редкие (единичные) морфы – морфы, частота которых более чем в 10 раз меньше доминантной.
2.2. Листоед ильмовый как объект исследования
Систематика вида
Класс: Насекомые (Insécta)
Отряд: Жесткокрылые (Coleoptera)
Семейство: Листоеды (Chrysomelidae)
Подсемейство: Козявки (Galerucinae)
Триба: Galerucini
Род: Galerucella
Вид: Листоед ильмовый (Galerucella luteola Mull)

Рисунок 1. Имаго листоеда ильмового
Морфология вида
Жуки буровато - или светло-желтые, часто рыжевато-желтые. На дорзальной стороне тела расположены темные пятна: одно на темени и три на переднеспинке (рис. 2.1). Вдоль бокового края проходит продольная полоса, а близ щитка расположено пятно. Внутренний край шва, заднегрудь и брюшко жука имеют черный цвет. Тело продолговато-овальное, слабовыпуклое, покрыто негустыми желтовато-серыми волосками, умерено блестящее. Длина жука 5-8 мм.

Рисунок 2. Личинки листоеда ильмового
Биология вида
Распространен в Евразии, Северной Африке, завезен в Северную Америку, в России – в лесостепной и степной зонах европейской части, в Крыму, на Кавказе, в Юго-Западной Сибири (Маслов, 1970).
Зимуют жуки в подстилке и трещинах коры. Они появляются в апреле-мае с распусканием листьев у ильмовых деревьев и приступают к питанию. Жуки выгрызают на листьях круглые дыры, не трогая крупных жилок. Жуки многократно спариваются и откладывают яйца в несколько приемов, обычно 400-700 шт., иногда до 1100. Генерация у листоеда ильмового обычно двойная, в Средней Азии – тройная. Наиболее вредоносна первая генерация.
Яички длиной 1 мм, бутылковидной формы, желтые или оранжево-желтые, мелкоячеистой структуры. Откладываются главным образом с нижней стороны листьев беспорядочными кучками по 10-30 шт. вместе, в стоячем и лежачем положении (рис. 2 ).
Яйца развиваются 5-9 дней. Личинка интенсивно желтого цвета, вдоль боков тела продольная черная полоса, на переднеспинке черное пятно, тергит 9-го сегмента брюшка черный, ноги красновато-бурые или черные, голова черная (рис. 2). Тело покрыто бородавками, несущими щетинки; длина личинки до 8 мм.
Личинки скелетируют листья снизу, не трогая жилок и верхней кожицы; живут они группами, их развитие длится около месяца. В конце мая в июне личинки окукливаются в трещинах почвы около ствола или неровностях коры.
Куколка длиной 5 мм и шириной 3 мм, светло-желтая; спина усеяна мелкими острыми бородавками. Стадия куколки продолжается около недели.
Молодые жуки питаются, как и весной, и также откладывают яйца. Личинки развиваются в июле, а в августе появляются жуки второго поколения. В степной зоне в отдельные годы наблюдается три поколения, на севере ареала – не более одного (Старк, 1931; Оглоблин, 1936; Померанцев, 1949; Вайнштейн, 1953; Стебаев и Поливанова, 1959).
Листоед ильмовый – фитофаг, кормится листьями различных видов ильмовых, отмечено повреждение конского каштана. Литературные данные (Померанцев, 1949; Вайнштейн, 1953; Бельговский, 1955) о предпочитаемости отдельных видов ильмовых несколько противоречат друг другу, но большинство сходится на том, что берест, ильм и вяз мелколистный сильнее повреждаются, вяз обыкновенный повреждается слабее. Поврежденные жуками и личинками листья скручиваются и засыхают. Наиболее опасной для деревьев считается первая генерация, развивающаяся в период, когда листья еще слабо развиты. Повторные объедания листвы сильно ослабляют деревья и способствуют их заселению стволовыми вредителями, поражению болезнями и усыханию.
В Волгоградской области периодически наблюдается массовое размножение листоеда ильмового в различных типах древесно-кустарниковых насаждений. Листоед повреждает прежде всего деревья, растущие в лесополосах, изреженных лесах, по опушкам.
обобщил информацию о вспышках численности листоеда ильмового в Волгоградской области и сопредельных регионов (2009, 2013).
На территории Волгоградской области очаги массового размножения листоеда ильмового регистрировались в 1956 г. на площади 1515 га. В 1957 г. их площадь увеличилась до 4045 га, и эти очаги оказались одними из крупнейших за период наблюдений. Однако к 1959 г. площадь очагов уменьшилась до 692 га. В течение 1960-1966 гг. и 1981–1992 гг. очаги листоеда продолжали действовать, однако их площади не были столь большими. В период 1969-1983 гг. вспышки массового размножения охватывали небольшие площади, причиной уменьшения масштабного распространения был контроль численности с использованием инсектицидов.
В 1996-1998 гг. повышенная численность листоеда наблюдалась в полезащитных и придорожных лесных полосах с участием ильмовых пород Быковского, Дубовского, Городищенского, Калачевского, Котельниковского, Октябрьского муниципальных районов области, а так же в городских озеленительных посадках Волгограда и Волжского. Повреждениям средней и сильной степени подверглись 37 % насаждений.
В 2004 г. бóльшая часть вязовых насаждений области была охвачена новой вспышкой массового размножения листоеда. В 2006-2007 гг. повреждения насаждений листоедом наблюдались на территории пяти муниципальных районов, а также Волгограда и Волжского.
Наиболее крупные очаги в Волгоградской области действовали в 1957, 1961, 1986, 1987, 2008 гг. За полувековой период наблюдений (1956–2013 гг.) средняя ежегодная площадь очагов распространения листоеда составила 484 га, максимальная площадь очагов составляла 6,4 тыс. га – это практически пятая часть площади вязовых насаждений области (31,7 тыс. га).
Методы исследования:
1. Ручной сбор особей имаго ильмового листоеда.
2. Фенетический анализ имаго.
Его проводили на высушенном материале, после фиксации в морилках с помощью ацетона, который лучше эфира или хлороформа сохраняет окраску насекомых (, 2009).
3. Определения морф рисунка переднеспинки насекомых с помощью микроскопа.
ГЛАВА 3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
3.1. Особенности морфологии листоеда ильмового по рисунку переднеспинки
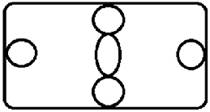
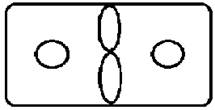
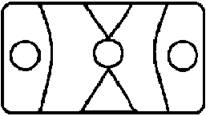
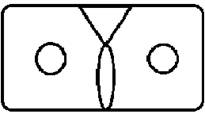
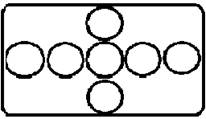
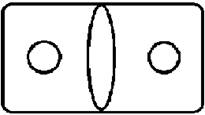
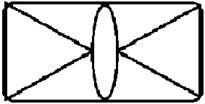
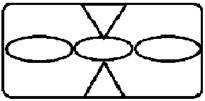
Рисунок на переднеспинке листоеда ильмового состоит из трех элементов – центрального и двух боковых (рис. 3). Форма и размер этих элементов, степень их обособления изменчивы.

Рисунок 3. Элементы рисунка переднеспинки листоеда ильмового
Центральный элемент может быть представлен единым пятном – овальной, круглой или треугольной формы, а может быть разбит на отдельные сегменты.
Боковые элементы чаще имеют форму круга или овала, которые не соединяются с центральным элементом. У отдельных особей отмечено слияние или соприкосновение центрального и боковых элементов.
В исследованных нами популяциях листоеда ильмового было выделено 11 морф (табл. 1) по характеру рисунка на переднеспинке. Наибольшая частота встречаемости отмечена для морфы 4 и 1 (18,33 % и 17,59 %), единично выявлена морфа 9 (0,83 %).
Таблица 1.
Морфы листоеда ильмового
Морфы | Описание морфы | Рисунок |
Морфа 1 (Mv 1) | Центральный элемент состоит из трех соприкасающихся пятен – овального в центре и двух круглых по краям, боковые пятна имеют округлую форму. |
|
Морфа 2 (Mv 2) | Центральный элемент представлен двумя овальными соприкасающимися пятнами, боковые пятна округлые. |
|
Морфа 3 (Mv 3) | Центральный элемент состоит из трех элементов – округлого центрального, соединенного с двумя треугольными пятнами по бокам, боковые пятна округлые. Центральный и боковые элементы обособлены между собой овальными линиями. |
|
Морфа 4 (Mv 4) | Центральный элемент состоит из двух частей – вертикального овального и треугольного пятен, боковые пятна округлые. |
|
Морфа 5 (Mv 5) | Центральный элемент состоит из трех округлых пятен, каждый из боковых – из двух пятен. Все компоненты рисунка соприкасаются между собой. |
|
Морфа 6 (Mv 6) | Большое центральное пятно овальной формы, вертикально расположено, боковые пятна округлые. |
|
Морфа 7 (Mv 7) | Центральное пятно в виде вертикально расположенного овала, боковые пятна – треугольные. Боковые пятна вершинами соприкасаются с центральным. |
|
Морфа 8 (Mv 8) | Центральный элемент состоит из горизонтально расположенного овального пятна и примыкающих к нему сверху и снизу треугольных. Боковые пятна крупные, овальные. Все компоненты рисунка соприкасаются между собой. |
|
Морфа 9 (Mv 9) | Центральный элемент состоит из двух треугольных пятен, не соединенных между собой. Каждый боковой элемент представлен двумя треугольными пятнами. Вершины всех пятен обращены к центру переднеспинки. |
|
Морфа 10 (Mv 10) | Центральный и каждый боковой элементы состоят из двух круглой формы пятен, одинакового размера. |
|
Морфа 11 (Mv 11) | центральный элемент состоит из трех элементов – крупного и мелкого круглых пятен, а также одного треугольного, основанием обращенного к центру переднеспинки. Боковые пятна округлые. Все компоненты рисунка обособлены друг от друга. |
|
3.2. Особенности полиморфизма исследуемых популяций листоеда ильмового
Сопоставление характера изменчивости рисунка переднеспинки листоеда ильмового в разных биотопах выявило сходные признаки и отличия (табл. 2).
В совокупности жуков, собранных с вязов на улице Баррикадной, было выделено три морфы, среди которых Mv 9 – единична. Морфы Mv 8 и Mv 9 отмечены только в этой популяции. Для популяции данного участка характерно минимальное разнообразие вариаций рисунка, преобладание двух морф и незначительное смещение равновесия в сторону одного из вариантов. 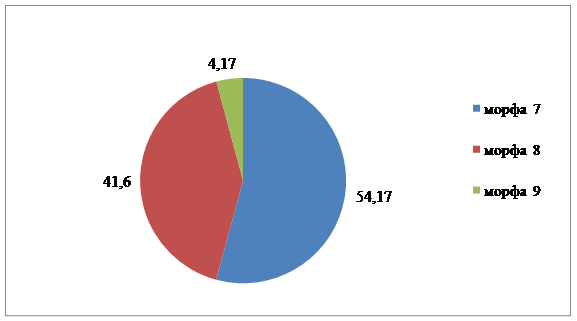
Рис. 3.2. Соотношение морф в популяциях листоеда ильмового на улице Баррикадной
Полиморфная структура популяций Краснооктябрьского района сходная. В каждой из популяций выделено по шесть морф, среди которых три общие (табл. 2). На пр. Ленина доминирует Mv 2, на ул. Маршала Еременко – Mv 1. Доминирующие морфы в этих популяциях составляют около 30 %.
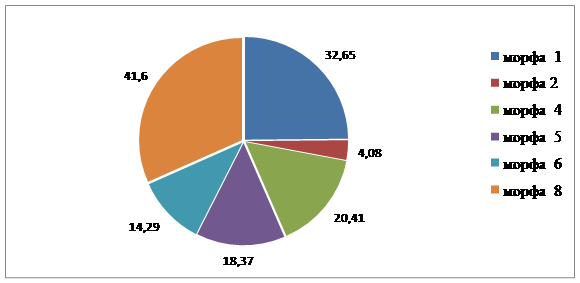
Рис. 3.3. Соотношение морф в популяциях листоеда ильмового на улице им. маршала С. Еременко
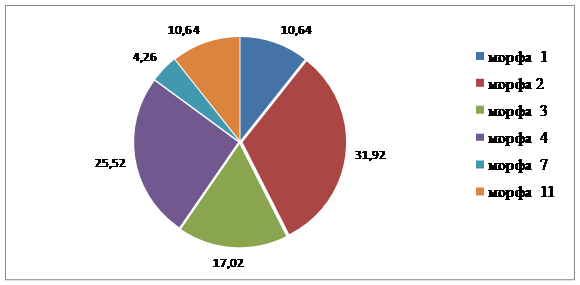
Рис. 3.4. Соотношение морф в популяциях листоеда ильмового на улице проспекта им.
Сопоставление характера изменчивости рисунка переднеспинки листоеда ильмового в разных биотопах выявило сходные признаки и отличия (табл. 2).
Таблица 2.
Полиморфная структура популяций листоеда ильмового
Морфа | Встречаемость, % | |||
Ул. Баррикадная | Ул. Маршала Ерёменко | Пр. Ленина | Общая | |
1 | - | 32,65 | 10,64 | 17,59 |
2 | - | 4,08 | 31,92 | 14,17 |
3 | - | - | 17,02 | 6,67 |
4 | - | 20,41 | 25,52 | 18,33 |
5 | - | 18,37 | - | 7,50 |
6 | - | 14,29 | - | 5,83 |
7 | 54,17 | - | 4,26 | 12,50 |
8 | 41,6 | - | - | 8,33 |
9 | 4,17 | - | - | 0,83 |
10 | - | 10,20 | - | 4,17 |
11 | - | - | 10,64 | 4,17 |
По соотношению сопутствующих и фоновых групп также наблюдается довольно высокая степень сходства (табл. 3)
Таблица 3.
Сравнительная характеристика полиморфной структуры исследуемых популяций
Показатели | Места исследования | ||
ул. Баррикадная | ул. Маршала Еременко | пр. Ленина | |
1. Количество морф | 3 | 6 | 6 |
2.Число и доля доминантных морф | 1 (54,16 %) Mv 7 | 1 (32,65 %) Mv 1 | 1 (31,92 %) Mv 2 |
3.Число и доля сопутствующих морф | 1 (41,67 %) Mv 8 | 2 (38,78 %) Mv 4, Mv 5 | 2 (42,54 %) Mv 4, Mv 3 |
4.Число и доля фоновых морф | 3 (28,57 %) Mv 6, Mv 10, Mv 2 | 3 (25,54 %) Mv 1, Mv 11, Mv 7 | |
5.Число и доля редких морф | 1 (4,16 %) Mv 9 | - | - |
Анализируя характер полиморфизма в исследуемых популяциях, можно отметить незначительное разнообразие вариаций – 3 или 6, при общем выявленном разнообразии 11 (рис. 3.5). Обращает внимание и значительная степень доминирования 2-3 морф. На них в популяциях приходится от 71 до 96 % особей. Подобные особенности характерны для популяций, испытывающих неблагоприятные воздействия или восстанавливающих свою численность после депрессии (Северцов, 2005).
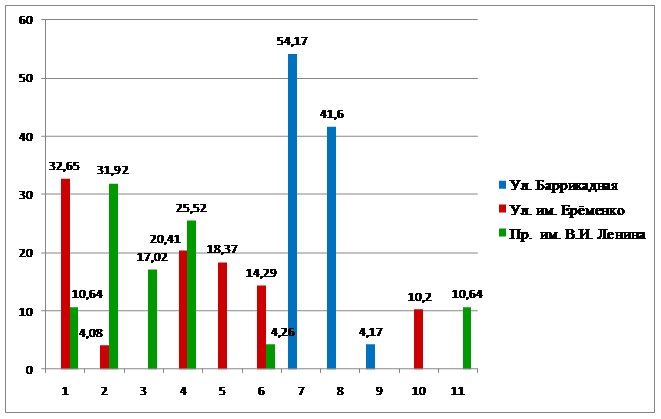
Рис. 3.5. Соотношение морф в популяциях листоеда ильмового
В Волгограде пик численности листоеда ильмового наблюдался в 2008 г., после которого последовал спад (Серый, 2013). В начале подъема численности (2014-2015 гг.) естественный отбор благоприятствует генотипам и морфам, определяющим высокую плодовитость. Возможно, поэтому на фоне невысокой численности листоеда наблюдается незначительное разнообразие в каждой популяции и высокая степень доминирования.
ВЫВОДЫ
1. Рисунок переднеспинки листоеда ильмового – полиморфный признак:
· каждый элемент рисунка варьирует по форме, размеру, расположению и степени обособленности;
· в исследуемых популяциях выделено 11 морф, различающихся по характеру изменчивости элементов рисунка.
2. Полиморфная структура каждой исследуемой популяции представлена небольшим набором морф, среди которых 1 доминантная и 1-2 сопутствующие, что свидетельствует о благоприятствовании естественного отбора морфам с интенсивным размножением и характерно для популяций, восстанавливающих численность после депрессии.
3. Сравнительный анализ полиморфной структуры исследуемых популяций выявил:
· значительные межпопуляционные различия полиморфной структуры по набору и соотношению морф;
· большее отличие популяции Ворошиловского района;
· болшее сходство популяций Краснооктябрьского района: выявлены три общие морфы, среди которых морфа 4 встречается с частотой выше 20 % и является сопутствующей;
· значительное преобладание по численности 2-3 морф в каждой из популяций.
4. В исследуемых популяциях можно отметить незначительное разнообразие вариаций – 3 или 6, при общем выявленном разнообразии 11 морф. Значительная степень доминирования характерна для 2-3 морф, что в популяциях составляет от 71 до 96 % особей. Подобные особенности характерны для популяций, испытывающих неблагоприятные воздействия или восстанавливающих свою численность после депрессии.
ЛИТЕРАТУРА
1. Береговой, гетерогенности в популяциях полиморфного вида / Журн. общ. биол. -1977. Т. 38, №2. - С. 182-184.
2. Вайнштейн, Б. А. К биологии ильмового листоеда в Южном Казахстане [Текст] //Труды республиканской станции защиты растений Казахского филиала ВАСХНИЛ, т. 1. Алма-Ата, изд-во АН Казахской ССР, 1953.
3. Калюжная, Н. С., Горбачева, О. В., Дидык листоед Galerucella luteola Müll. (Coleoptera. Crysomelidae) как вредитель зеленых насаждений на юге Ергеней (Калмыкия)// Энтомол. обоз. -1995. №1. - С. 45-51.
4. Кайданов, популяций: учеб. для вузов по направлению "Биология" и специальностям "Биология" и "Генетика" / Под ред. -Вечтомова ; Прил. -Борисова 319, [ 1 ]с. ил. 22 см. М. Высш. шк. -1996 – С. 320
5. Креславский, миграция. Последствия для изменчивости количественных признаков. [Текст]: / А. Г Креславский., , A. B. Михеев, .// Журн. общ. биол.- 1987. Т. 48, вып. 5. - С. 602-613.
6. Маслов, ильмовых пород и меры борьбы с ними. [Текст]: / // М.: Лесная промышленность -1970. С. 52-56.
7. Новоженов, и микроэволюция [Текст]: / // Онтогенез, эволюция, биосфера. М.: Наука, 1989. – С. 144-160..
8. Новоженов изменчивость популяции [Текст]: / // Журн. общ. биол. 1989. Т.50. № 2. - С. 171-183.
9. Новоженов диморфизм полиморфизма окраски у некоторых видов насекомых [Текст]: / // Журн. общ. биол. - 1997. Т. 58. № 1. - С. 26-43.
10. Орлова листоед (Galerucella luteola Müll.) вредитель городских насаждений [Текст]: / // Альманах-2008 /под ред. д-ра хим. наук, проф. ; МААНОИ. ВО, РЭА, РАЕН, ВолГУ. Волгоград: Изд-во Волгогр. гос. ун-та - 2008. - С.127-131.
11. Померанцев насекомые и борьба с ними в лесах и лесных полосах юго-востока европейской части СССР [Текст]:/ // М-Л.: Гослесбумиздат - 1949. – С. 212.
12. Северцов, эволюции. [Текст]: / учеб. для студентов вузов, обучающихся по направлению "Биология". / Северцов, А. С. - М.: Гуманитар. изд. центр ВЛАДОС - 2005 - С. 118-122.
13. Серый, размножение Ильмового листоеда в Волгоградской области [Текст]:/ // Известия Санкт-Петербургской лесотехнической академии. СПб: СПбГЛТА- 2009. - Вып.187. - С. 304-310.
14. Серый, состояние массовых размножений ильмового листоеда на юге России. [Текст]:/ Г. А Серый// Исследования природного комплекса окрестностей озера Баскунчак: сб. науч. ст. ФГБУ «Государственный природный заповедник «Богдинско-Баскунчакский». Волгоград: Волгоградское научное изд-во - 2013. - С. 121-126.
15. Старк, ильмовых. [Текст]:/ В. Н Старк // Вредители и болезни полезащитных лесных насаждений и меры борьбы с ними. М-Л.: Сельхозгиз-1951.
16. Степанчук, Н. М., Прилипко, по общей экологии. 9 класс [Текст]:/ , - Волгоград: УчиС. 78-85.
17. Яблоков, . Эволюция, популяция, признак [Текст]:/ А. В Яблоков - М.: Просвещение - 1980.-С. 136.
18. , Юсуфов учение./ , - М.: Высш. шк. - 1998. – С.336.
Приложение 1
Места сбора материала для исследования
Приложение 2
Собранный материал на улице Еременко

Собранный материал на проспекте им.

Приложение 3
Собранный материал на улице Баррикадной

