

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Примечание. Даны оценки статистической значимости отличия минимальных значении к кди) и ккмакс (оценки одинаковы) от соседних значений в данном столбце: * -р < 0.05, ** -р < 0.01. В левой колонке - номера электродов продольной цепочки, составивших пары ЭЭГ-отведений.
Таблица 2.
Зависимость величины КК(0) и ККмакс от расстояния в продольной цепочке от затылка ко лбу (з-л) и наоборот (л-з)
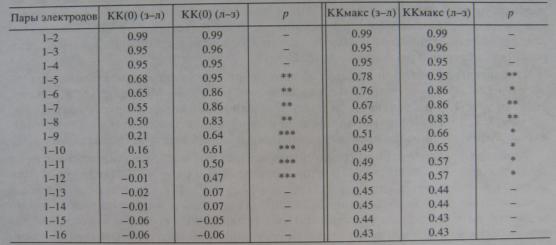
Примечание, В левой колонке - номера электродов продольной цепочки, составивших пары ЭЭГ - отведений; электродом № 1 является крайний задний или передний электрод, относительно которого вычисляются КК. Звездочки - случаи статистически значимых различий для КК, взятых на равных расстояниях от краевых электродов цепочки: * — р < 0.05; ** - р < 0.01; *** — р < 0.001.
Сходным с данными по индексу ОС оказался и ход кривых зависимости КК от межэлектродного расстояния в продольной цепочке: в направлении от затылка ко лбу падение значений КК было гораздо более крутым, чем в обратном направлении (табл. 2). При этом фазовый сдвиг не является здесь главной причиной снижения КК при увеличении межэлектродного расстояния, поскольку это в равной мере касалось как КК(0), так и ККмакс (табл. 2).
Наконец, динамика КК(0) для пар соседних ЭЭГ-отведений в поперечных Цепочках (табл. 3) достаточно точно повторяла аналогичные зависимости для индекса ОС (рис. 2).
Можно полагать, что отмеченное, по крайней мере для изучавшихся альфа-ритмичных ЭЭГ, совпадение пространственных распределений индексов ОС, с одной стороны, и КК - с другой, отражает приуроченность обеих оценок к какой-то базовой структурной особенности ЭЭГ. Поскольку индексы ОС более контрастно, чем КК, демонстрируют неоднородность биопотенциального поля, например в продоль ном направлении (см. рис. 1 и табл. 1), весьма вероятно, что в основном именно временная согласованность сегментной структуры двух ЭЭГ определяет расчетное значение ККмакс между ними.
Таблица 3
Значения КК(О) и ККмакс для ЭЭГ в соседних парах отведений
в передней (лоб) и задней (затылок) цепочках электродов
Примечание. Звездочки - случаи статистически значимых различий между КК, взятых для одноименных пар электродов в обеих цепочках: * - p < 0.05; ** -р < 0.01; *** - p < 0,001.
Таблица 4
Зависимость КК(0) и ККмакс от межэлектродного расстояния в поперечных передней (лоб) и задней (затылок) цепочках
Примечание. Статистически значимый уровень КК при p < 0.05 для указанных данных равен 0.21; сред-неквадратическое отклонение для средних значений КК не превышает значения 0.007.
Действительно, при выраженной пачечной альфа-активности ЭЭГ наибольшая фазовая стабилизация наблюдается в пределах альфа-веретен [23], а моменты резких изменений билатеральных фазовых отношений в ЭЭГ с наибольшей вероятностью приходятся на начало и конец альфа-веретен [14, 20]. Следовательно, чем чаще и более точно эти веретена, или другого рода квазистационарные фрагменты ЭЭГ, совпадают по времени в паре ЭЭГ-записей, тем выше будет максимальное значение кросскорреляционной функции, усредненной по всему фрагменту записи ЭЭГ.
Таким образом, именно исходная согласованность сегментной структуры двух альфа-ритмичных ЭЭГ может обусловливать высокие значения КК между ними. При меньшей структурированности и выраженности основного ритма ЭЭГ, например в височных или нижнелобных отведениях, динамические характеристики индексов ОС и КК могут существенно расходиться. Возможно, этим объясняются различия в зависимости значений КК и индексов ОС от межэлектродного расстояния в поперечных цепочках, когда первые претерпевают монотонное падение на всем своем протяжении (табл. 4), а вторые заметно возрастают для пары ЭЭГ наиболее далеко расположенных, но гомологичных отведений (рис. 2).
ЗАКЛЮЧЕНИЕ
Полученные данные о закономерностях изменения индекса ОС в парах ЭЭГ-отве-дений по линиям продольного и поперечного "сечений" коркового биоэлектрического поля свидетельствуют о значительной неоднородности этого поля в отношении временнбй согласованности его региональной сегментной структуры. В многократных записях ЭЭГ (п = 20-24) у каждого из двух испытуемых в условиях спокойного бодрствования отмечены хорошо локализованные корковые зоны, для которых показано статистически значимое по сравнению с другими корковыми регионами снижение индексов ОС. При этом конкретные значения индексов ОС ЭЭГ зависят как от межэлектродного расстояния, так и от степени морфофункционального подобия соответствующих корковых образований. Однако представить полную пространственную картину операциональной синхронности корковых структур можно только при оценке индекса ОС не просто в парах ЭЭГ-отведений, но и во всех из возможных комбинациях по два, по три и т. д. Весьма вероятно, что в зависимости от конкретной функциональной нагрузки различные корковые регионы будут выстраиваться в специфические пространственные модули, проявляющиеся на уровне ЭЭГ во взаимной стабилизации сегментной структуры соответствующих ЭЭГ-отведений [18]. Проверке этого предположения будет посвящено следующее исследование.
Работа выполнена при частичной поддержке фонда "Университеты России - фундаментальные исследования" (проект ).
СПИСОК ЛИТЕРАТУРЫ
1. "Алфавит" ЭЭГ: типология стационарных сегментов ЭЭГ человека // Индивидуально-психологические различия и биоэлектрическая активность мозга человека / Под ред. . М.: Наука, 1988. С. 56-70.
2. , , Фингелъкурц Ал. А. и др. Топологическое картирование процесса синхронизации моментов резких перестроек в мультиканальной ЭЭГ у человека // Журн. высш. нерв. деят. 1997. Т. 4. № 1. С 32-37.
3. Каплан А Я., Фингелъкурц Ан. А., Фингелъкурц Ал А. и др. Временная согласованность физических перестроек основных частотных компонентов ЭЭГ-сигнала // Журн. высш. нерв. деят. 1998. Т. 48. № 5. С. 816-826.
4. Нестационарность ЭЭГ: методологический и экспериментальный анализ//Успехи физиол. наук. 1998. Т. 29. № з. С. 35-55.
5. Синхронная электрическая активность мозга и психические процессы. М.: Наука, 1987. 156 с.
6. Методы корреляционного и регрессионного анализа. М.: Финансы и статистика, 1983. 302 с.
7. , , Апанасионок B.C. Формирование биопотенциального поля человека, Л.: Наука, 1979. 163 с.
8. Barlow J. S. Methods of analysis of nonstationary EEGs, with emphasis on segmentation techniques: a comparative review //J. Clin. Neurophysiol. 1985. V. 2. P. 267-304.
9. Basar E. Brain natural frequencies are causal factors for resonances and induced rhythms // Induced Rhythms in the Brain / Eds E. Basar, Т. Н. Bullock. Boston: Birkauser, 1992. P. 425-467.
10. Bodenstein G. t Praetorius HM. Feature extraction from the electroencephalogram by adaptive segmentation // Proc. IEEE. 1977. V. 65. P. 642-652.
11. Bullock Т.Н., McClune M. C., Achimowicz JZ. et al. EEG coherence has structure in the millimeter domain: subdural and hippocampal recordings from epileptic patients // EEG a. Clin. Neurophysiol. 1995. V. 95. P. 161-177.
12. Burgess A. P., GruzelierJ. How reproducible is the topographical distribution of EEG amplitude?// Intern. J. Psychophysiol. 1997. V. 26. P. 113-119.
13. Homan R. W., Herman J., Purdy P. Cerebral location of international 10-20 system electrode placement // EEG a. Clin. Neurophysiol. 1987. V. 66. P. 376-382.
14. HooveyZ. V., Heinemann U„ Creutzfeldt O. D. Inter-hemispheric "synchrony" of alpha waves //EEG a Clin Neurophysiol. 1972. V. 32. № 4. P. 337-347.
15. Hori H. f Hayasaka K., Sato K. et at. A study on phase relationship in human alpha activity. Correlation of different regions // EEG a. Clin. Neurophysiol. 1969. V. 26. P. 19-24. 6
16. Jansen В.И. Quantitative analysis of the electroecephalograms: is there chaos in the future // Intern. J. put. 1991. V. 27. P. 95-123.
17. Kaplan A. Y., Fingelkurts ALA., Fingelkurts An. A. et al. Topological mapping of sharp reorganization synchrony in multichannel EEG //Amer. J. Electroneurodiagnostic Technol. 1997. V. 37. p. 265-275.
18. Kaplan A. Ya. Spatial EEG Moduli: Change-Point Analysis // Proceedings of 5lh Congress of the International Organization of Societies for Electrophysiological Technology (OSET). Washington D. C., 1995-P. 12.
19. Kaplan A. Y., Kadr I. The multiple combinatorial analysis of segmental EEG structure // Proceedings of the International Sympos. Mathematical approaches to brain functioning diagnostics. Prague, 1990. P. 69.
20. Lehmann D. Multichannel topography of human alpha EEG fields // EEG a. Clin. Neurophysiol.
1971. V. 31. P. 439-449.
21. Lehmann D., Ozaki H., Pal I. EEG alpha map series: brain micro-states by space-oriented adaptive
segmentation//EEG a. Clin. Neurophysiol. 1987. V. 67. P. 271-288.
22. Lehmann D. Brain electric field mapping and map analysis in psychiatry: The "atoms of thought" //
Biol. Psychiatry. 1991. V. 2. P. 391-394.
23. Levine R. B., Smith R. P., Hawkes G. R. On synchrony of the alpha rhythms // Aerospace Med. 1963.
V. 34. №4. P. 349-352.
24. McEwen J A., Anderson G. B. Modeling the stationary and gaussianity of spontaneous electroenceph-alographic activity // IEEG Transactions on Biomed. Engng. 1975. V. 22. № 5. P. 361-369.
25. Ozaki H„ Suzuki H. Transverse relationships of the alpha rhythm on the scalp // EEG a. Clin. Neurophysiol. 1986. V. 66. P. 191-195.
26. Thatcher B. W., Krause P. J., Hryhyk M. Cortico-cortical associations and EEG coherence: a two-compartmental model // EEG a. Clin. Neurophysiol. 1986. V. 64. P. 123-143.
Кафедра физиологии человека и
животных Московского государственного
университета им.
Поступила в редакцию
6.Х. 1998
Принята в печать
21.XII. 1998
SPATIAL SYNCHRONY OF HUMAN EEG SEGMENTAL STRUCTURE
KAPLAN A. Ya., FINGELKURTS An. A., FINGELKURTS ALA., SHISHKIN S. L, IVASHKO R. M.
Lomonosov State University, Moscow
Topographic features of spatial synchronization of sharp changes, or rapid transition processes (RTP), were studied in human EEG recorded from longitudinal and transversal electrode arrays. A new algorithm, the EEG Threshold Scanning, was proposed for the detection of the RTP. Synchronization of the RTP was estimated by Operational Synchrony Index (OSI) based on the difference between the actual and stochastic frequency of RTP coincidence in a pair of EEG channels. The relationship between the OSI and interelectrode distance was not monotonous. The OSI depended also on the extent of morpho-functional similarity between two cortical areas. Similar results were obtained for crosscorrelation calculated for the same pairs of the EEG derivations. The existence of dynamic spatial modules which incorporate different brain areas by complementary stabilization of their functional states is discussed.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
