

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Публикация доступна для обсуждения в рамках функционирования постоянно
действующей интернет-конференции “Бутлеровские чтения”. http:///readings/
УДК 541.49.183:546.562.’723:547.854.5. Поступила в редакцию 27 декабря 2014 г.
Влияние солей одно - и двух - зарядных катионов металлов
на сорбцию Н+ и ОН - ионов на мицелии базидиомицетов
© Чухно* Александр Сергеевич, ,
, Банкина+ Анастасия Николаевна,
и
Кафедра физической и коллоидной химии. Санкт-Петербургская государственная химико-фармацевтическая академия. . г. Санкт-Петербург, 197376. Россия.
Тел.: (812) 234-11-38. E-mail: *****@***ru , *****@***ru
___________________________________________
*Ведущий направление; +Поддерживающий переписку
Ключевые слова: базидиомицет, потенциометрическое титрование, точка нулевого заряда.
Аннотация
В работе представлены исследования коллоидных свойств мицелия базидиомицетов. Целью работы является изучение адсорбции на мицелии базидиомицетов Abortiporus biennis и Poliporus ciliatus, а так же определение точки нулевого заряда мицелия, и исследование влияния электролитов на ее значение. Исследовеия проводились методом потенциометрического титрования. Точка нулевого заряда мицелия Poliporus ciliatuиs равна 5.0, а точка нулевого заряда мицелия Abortiporus biennis равна 6,5. Установлено, что на мицелии специфически сорбируются катионы простых электролитов, о чем свидетельствует сдвиг точки нулевого заряда в растворах электролитов в кислую область. С увели-чением концентрации либо заряда катиона влияние на точку нулевого заряда возрастает. Это говорит об избирательности сорбции на поверхности мицелия. Мицелий базидиомицетов Abortiporus biennis и Poliporus ciliatus можно использовать как в качестве носителя, так и сорбента катионов металлов.
Введение
Базидиальные грибы являются перспективным источником биологически активных со-единений, перспективных для использования в фармации и медицине. В настоящее время изу-чены лишь отдельные представители данного отдела грибов.
Известны их иммуномодулирующее, противоопухолевое, противовирусное, антибакте-риальное, гиполипидемическое, антиоксидантное, противовоспалительное и другие свойства. Выделены и охарактеризованы метаболиты некоторых базидиомицетов, а также их мицелий. В частности, полисахаридные компоненты, выделенные из плодовых тел или мицелия бази-диомицетов, относятся к перспективным иммуномодуляторам, способным регулировать функции иммунной системы.
Ранее основным направлением исследований базидиальных грибов было выращивание и изучение их плодовых тел. Исследования показали, что источником биологически активных веществ служат не только плодовые тела базидиомицетов, но и вегетативный мицелий.
Благодаря развитию биотехнологии стало возможным искусственное выращивание бази-диомицетов в виде вегетативного мицелия методом глубинного культивирования в жидких питательных средах, обеспечивающим высокий выход биомассы и позволяющим выделять биологически активные метаболиты, применяемые для создания биопрепаратов.
Глубинное культивирование позволяет осуществлять регулируемый синтез нужных метаболитов, сократить время процесса и получать стандартный целевой продукт.
Биологическая активность базидиомицетов определяется рядом компонентов их мице-лия, среди которых наибольшее значение имеют полисахариды, терпеноиды и иммуномоду-лирующие протеины (лектины), витамины, антибиотические вещества. По сравнению с про-дуктами химического синтеза, они менее токсичны и более эффективны при профилактике и лечении многих болезней человека и животных. Полисахариды базидиомицетов обладают иммуномодулирующей, противоопухолевой, противовоспалительной активностью. В настоя-щие время известны антивирусные, антибактериальные и противогрибковые свойства грибов базидиомицетов [1].
В связи с возможностью использования мицелия, как носителя и сорбента лекарствен-ных препаратов необходимо изучать коллоидные свойства мицелия [2-5]. Данные исследова-ния позволят подобрать правильные условия сорбции, удобную лекарственную форму и нужную форму сорбируемого лекарственного вещества. В перспективе планируется создание новых лекарственных форм, на основе уже существующих субстанций [6-9].
Больший интерес представляет группа гименомицетов. Это самая большая группа по числу видов среди базидиальных грибов и самая известная. Она объединяет два порядка: порядок пластинчатых, или агариковых (Agaricales) грибов и порядок не пластинчатых, или афиллофоровых грибов (Aphyllophorales). Исследуемые в работе Abortiporus biennis и Polipo-rus ciliatus являются представителями этой группы.
В последние время широко исследуются физикохимические и коллоидные свойства сис-тем содержащих сложные биологически активные соединения, а также устойчивость их дис-персий [10-17].
Известно, что мицелий грибов в основном состоит из полисахаридов и белков, адсорб-ционные свойства которых были исследованы в более ранних работах [18-24]. Природные высокомолекулярные вещества (ВМВ) характеризуются постоянным значением молекуляр-ной массы. Они являются полидисперсными системами. Специфические свойства полимеров обусловлены существованием двух типов связей – химических и межмолекулярных, удержи-вающих макромолекулярные цепи около друг друга, а так же гибкостью цепей, связанной с внутренним вращением звеньев [25].
Целью работы является изучение коллоидных свойств мицелия базидиомицета, опреде-ление точки нулевого заряда мицелия, и исследование влияния электролитов на ее значение.
Экспериментальная часть
В качестве объектов исследования выбран мицелий базидиомицетов Abortiporus biennis и Polipo-rus ciliatus. Штамм базидиомицета Polyporus ciliatus (0626) был получен из лаборатории микологии БИН РАН. На плотной питательной среде гриб Polyporus ciliatus образовывал колонии, состоящие из скопления мицелия, белого цвета, ватообразные, опушенные, с неровным краем, с хорошо развитым воздушным мицелием белого цвета. Субстратный мицелий при выращивании на суслоагаре не пигментирован (рис. 1). Бурокоричневый пигмент образовывался по внешней кромке при выращива-нии на жидкой глюкозопептонной среде (рис. 2). Отмечался запах прелой земли. При старении куль-туры более 3-х недель образовывались зачатки плодового тела. При микроскопии культуры, выра-щенной на суслоагаре 4% в течение 5-7 суток, были обнаружены пряжки на септированном мицелии, которые характерны для базидиомицетов (рис. 3). При культивировании на жидких питательных сре-дах в динамических условиях культура росла в виде пеллет, крупных, гладких, шарообразных скоп-лений биомассы мицелия, размером от 0.6 до 1.2 см. Выращивание культуры проводили методом глубинного культивирования. Культуру выращивали 10 суток при температуре 23-24 оС в колбах Эрленмейера вместимостью 750 мл (объем питательной среды 150 мл) на лабораторной качалке (n = 220 об/мин) при непрерывном перемешивании.
В качестве базовой питательной среды использовали полусинтетическую глюкозопептонную среду (ГПС) следующего состава: пептон – 2.5 г/л; глюкоза – 10 г/л; натрия хлорид (NaCl) – 0.5 г/л; кальция хлорид (СаСl2) – 0.5 г/л; магния сульфат (MgS04) – 0.5 г/л; дигидрофосфат калия (КН2Р04) – 0.6 г/л; гидрофосфат калия (К2НР04) – 0.4 г/л; мальт-экстракт – 1.6 г/л; рН = 6.0. Стерилизацию сред проводили в автоклаве в течение 30 минут при избыточном давлении 0.5 атм [26-27].
В качестве исходного посевного материала использовали культуру, выращенную на скошенном сусло-агаре, откуда асептично брали около 1 см2 пленки гриба, для посева в жидкую питательную среду. Для получения посевного материала культуру выращивали в статических условиях 10-14 дней на ГПС при температуре 24 °С. При этом в среду добавляли стерильные бусы. Перед внесением посевного материала в колбы для ферментации его встряхивали до образования мелких фрагментов мицелия. Суспензию посевного материала вносили в ферментационные колбы в количестве 10 мл.
Мицелий гриба от культуральной жидкости после ферментации отделяли фильтрованием под вакуумом на воронке Бюхнера через бумажный фильтр. Полученную биомассу обрабатывали этило-вым спиртом для обезвоживания и сушили на воздухе при комнатной температуре. Далее высушенную биомассу измельчали в ступке до порошкообразного состояния. Выход определяли по сухому весу. Мицелий Abortiporus biennis получен аналогичным путем.
Рис. 1. Polyporus ciliatus (0626) на сусло-агаре |
Рис. 2. Polyporus ciliatus (0626) на глюкозопептонной среде |
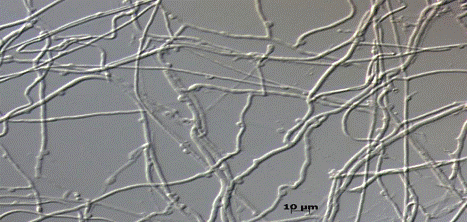
Рис. 3. Микроскопия Polyporus ciliatus (0626) на сусло-агаре (увеличение 1.5х1000)
В качестве простых электролитов (источников одно-, двухзарядных ионов) использовали водные растворы КСl (0.01 моль/л), СаСl2 (0.01 моль/л), K2SO4 (0.01 моль/л).
Точку нулевого заряда (рНтнз) находили методом непрерывного потенциометрического титро-вания [28-29]. В ячейку для потенциометрического титрования помещали 25 мл исследуемой диспер-сии, 30 минут пропускали воздух, освобожденный от СО2 и титровали раствором КOH (0.0129 моль/л) или НС1 (0.010 моль/л). Титрант вводили порциями по 0.1 мл с помощью микробюретки с интервалом в 1 минуту. Растворы КOH готовили на дистиллированной воде, предварительно освобожденной от СО2 воздуха кипячением в течение 30 минут. После каждой добавленной порции титранта измеряли значения рН раствора. Регистрация рН осуществляется на рН-метре – милливольтметре рН-673 с точностью измерения рН ±0.05 единиц рН.
По результатам потенциометрического титрования были построены зависимости рН иссле-дуемого раствора от объема добавленного титранта (V, мл) для растворов без адсорбента и с адсор-бентом. Расчет величины адсорбции проводился по следующему алгоритму. В кислой области преоб-ладает адсорбция ионов водорода, а в щелочной области ионов ОН-. Значение рН точки пересечения кривых титрования с адсорбентом и без него соответствует условию равенства адсорбированных Н+ и ОН--ионов – это соответствует рНТНЗ. Слева от рНТНЗ преобладает адсорбция ионов Н+ в результате чего поверхность заряжается положительно, а справа преобладает адсорбция ОН - – поверхность мице-лия заряжается отрицательно. Таким образом, методом потенциометрического титрования опреде-ляется разница между адсорбцией ионов Н+ и ОН-. Кривые титрования с адсорбентом и без срав-нивают и рассчитывают разницу между количеством адсорбированных ионов Н+ – (ОН-) отнесенное к единице массы или поверхности адсорбента:
![]() (ОН - - H+) =
(ОН - - H+) = 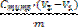 (1)
(1)
где (H+ – ОН-) – разница между количеством адсорбированных ионов Н+ и ОН - отнесенная к единице массы адсорбента, моль/г; Ститр – концентрация титранта КOH (HCl), моль/л;
V1 – объем раствора титранта, пошедший на титрования раствора без адсорбента, мл;
V2 – объем раствора титранта, пошедший на титрование раствора с адсорбентом, мл;
m – масса адсорбента, г.
Важно отметить, что:
![]() (ОН - - H+)= -
(ОН - - H+)= - ![]() (Н+- ОН-) (2)
(Н+- ОН-) (2)
Расчет – (Н+ – ОН-) производят для одинаковых значений рН раствора с адсорбентом и без, в нашей работе при значениях рН от 3.0 до 8.0. Через заданное значение рН проводят линию параллель-ную оси абсцисс и находят V1 и V2.
Погрешность измерения значений рН составила 0.05 единиц рН.
Результаты и их обсуждения
Обычно для электролитов различают специфическую и ионообменную адсорбцию. Ионообменная адсорбция реализуется за счет простого электростатического взаимодействия, в ней участвуют противоионы, количество адсорбированных ионов зависит от состава и кон-центрации различных ионов в растворе, их заряда. Любая другая адсорбция является специ-фической. Причем механизмы специфической адсорбции, виды взаимодействий адсорбата с поверхностью могут быть различны. Например, поверхностно-активные веществ адсорбиру-ются вследствие гидрофобного эффекта, располагаясь на границе раздела фаз так, чтобы умень-шить их разность полярностей, а следовательно и межфазное натяжение. На поверхности могут происходить различные химические взаимодействия с образованием ковалентных, до-норно-акцепторных, ионных и водородных связей [30-32]. Удельная адсорбция (Г) – избыток растворенного вещества в поверхностном слое по сравнению с объемом раствора, отнесенный либо к единице поверхности вещества – моль/м2, либо к единице массы вещества – моль/кг. Количество адсорбированного вещества определяют по убыли концентрации растворенного вещества в объеме водной фазы.
Для нерастворимых систем (в нашем случае мицелия) заряд поверхности (σо) опреде-ляется количеством адсорбированных Н+ и ОН--ионов, которые для мицелия являются потен-циалопределяющими ионами. Величина рН растворов, при которой σ = σо = 0, называется рНтнз. В рНтнз адсорбция Н+ и ОН--ионов одинакова (Гн+ = Гон-).
Существуют два метода изучения адсорбционных процессов: электрокинетические ис-следования и прямые адсорбционные измерения. Прямое измерение количества адсорбиро-ванного вещества позволяет оценить адсорбционную и буферную емкость сорбентов. Для использования мицелия в качестве носителя необходимо определить его адсорбционную ём-кость. Количество активных центров на поверхности мицелия можно оценить методом непре-рывного потенциометрического титрования. Для определения рНтнз для мицелия был прове-ден ряд экспериментов потенциометрического титрования с различными неорганическими электролитами. По полученным результатам были построены кривые представленные на рис. 4 для мицелия Polyporus ciliates и рис. 5 для мицелия Abortiporus biennis.
Точка пересечения кривых титрования с адсорбентом и без адсорбента соответствует равенству количества адсорбированных протонов и гидроксид ионов. В области рН < рНТНЗ мицелий заряжен положительно, а рН > рНТНЗ отрицательно, что согласуется с результатами электрокинетических измерений [4]. Отметим, что величина отрицательного заряд больше, чем положительного. Изоэлектрическая точка и точка нулевого заряда в отсутствии специфи-ческой сорбции совпадают. Их расхождение связано с наличием в водной фазе ионов, способ-ных к специфическому взаимодействию.
На рис. 5 приведены кривые потенциометрического титрования мицелия водных раст-ворах различных электролитов (вода, хлорид калия 0.01 моль/л, сульфат калия 0.01 моль/л, хлорид кальция 0.01моль/л). Точка пересечения кривой с адсорбентом (2) и без адсорбента (1) соответствует рНтнз.
В области рН меньше рНтнз кривая с мицелием идет выше, чем кривая без мицелия, вследствие адсорбции Н+ ионов. В области рН больше рНтнз кривая с мицелием идет ниже, вследствие адсорбции ОН--ионов, что эквивалентно десорбции Н+. Из зависимостей видно, что с ростом ионов калия в растворе кривая смещается в кислую область, что свидетельствует о незначительной специфической сорбции катионов калия, при чем данная тенденция на-блюдается для мицелия как Polyporus ciliates, так и для мицелия Abortiporus biennis. Это смещение увеличивается с ростом концентраций ионов калия (раствор K2SO4) в соответствие с теоретическими представлениями о специфической адсорбции [33-34].
а) |
б) |
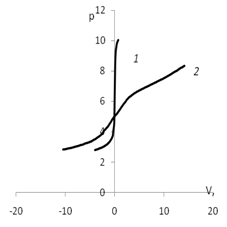
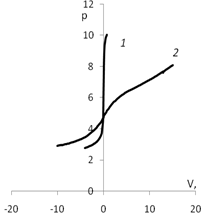
Рис. 4. График потенциометрического титрования: а) 1 – кривая титрования воды; 2 – кривая титрования дисперсии мицелия Polyporus ciliatus; б) 1 – кривая титрования раствора КСl
(0.01 моль/л); 2 – кривая титрования дисперсии мицелия Polyporus ciliatus КСl (0.01 моль/л)
а) |
б) |
в) |
г) |
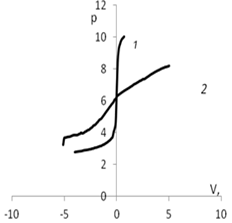
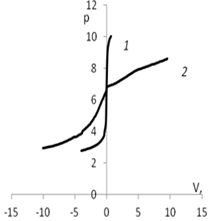
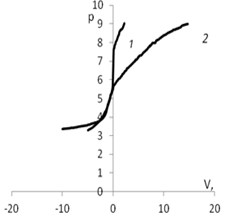
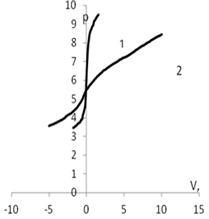
Рис. 5. Изменение рН в зависимости от количества добавленного титранта: а) 1 – кривая титрования раствора КСl (0.01 моль/л); 2 – кривая титрования дисперсии мицелия Abortiporus biennis c КСl
(0.01 моль/л); б) 1 – кривая титрования воды; 2 – кривая титрования дисперсии мицелия Abortiporus biennis в воде; в) 1 – кривая титрования раствора СаСl2 (0.01 моль/л); 2 – кривая титрования дисперсии мицелия Abortiporus biennis СаСl2 (0.01 моль/л); г) 1 – кривая титрования раствора K2SO4 (0.01 моль/л); 2 – кривая титрования дисперсии мицелия Abortiporus biennis c K2SO4 (0.01 моль/л)
Из кривых потенциометрического титрования рассчитывались количества адсорбиро-ванных ионов водорода и гидроксил ионов при разных значениях рН. Результаты расчетов представления в виде графиков на рис. 6 и 7.
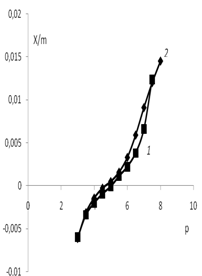
Рис. 6. Зависимость адсорбции ионов мицелия Poliporus ciliatus от рН: 1 – водная
дисперсия мицелия; 2 – водная дисперсия мицелия с добавлением KCl (0.01 моль/л)
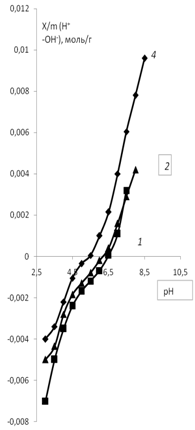
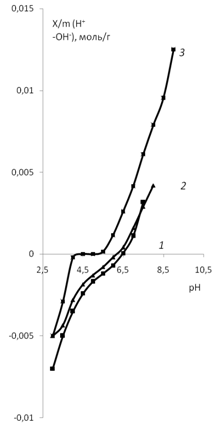
а) б)
Рис. 7. Зависимость адсорбции ионов мицелия Abortiporus biennis от рН: 1 – водная
дисперсия мицелия; 2 – водная дисперсия мицелия с добавлением KCl (0.01 моль/л);
3 – водная дисперсия мицелия с добавлением СаCl2 (0.01 моль/л);
4 – водная дисперсия мицелия с добавлением K2SO4 (0.01 моль/л).
Сравнение кривых зависимостей (рис. 7а и б) показывает, что влияние катиона кальция выше, чем катиона калия. Кривая для хлорида кальция имеет ступенчатый характер, что по-видимому связано с тем, что ион кальция двухзарядный, и кривая титрования для него носит ступенчатый характер, как для любых многозарядных ионов.
Максимальные значения адсорбции в области высоких значений рН соответствуют ко-личеству отрицательных активных центров на поверхности мицелия. Минимальные значения в области низких рН-количеству положительных активных центров.
Важно отметить, что для биполярных сорбентов, коим является мицелий или белки, количество адсорбированного вещества зависит не только от концентрации адсорбата, но и от значений рН водной среды, при которых идет процесс адсорбции. В работе определены зна-чения рНтнз для мицелия базидиомицетов Poliporus ciliatuиs и Abortiporus biennis. Определены области наибольшей устойчивости водных дисперсий мицелия Poliporus ciliatuиs (рН < 4.0 и рН > 5.5) и Abortiporus biennis (рН < 5.5 и рН > 7.0).
Выводы
Установлено, что точка нулевого заряда мицелия Poliporus ciliatuиs равна 5.0, а точка нулевого заряда мицелия Abortiporus biennis равна 6.5. Установлено, что на мицелии специи-фически сорбируются катионы простых электролитов, о чем свидетельствует сдвиг точки ну-левого заряда в растворах электролитов в кислую область, при этом с увеличением концент-рации, либо заряда катиона влияние на точку нулевого заряда возрастает. Это говорит об избирательности сорбции на поверхности мицелия. Мицелий базидиомицетов Abortiporus biennis и Poliporus ciliatus можно использовать, как в качестве носителя, так и сорбента ка-тионов металлов.
Литература
[1] , , Пучкова базидиальные грибы – продуценты антивирусных соединений. Современная микология в России. Т.2. М.: Национальная академия микологии. 2008. 18с.
[2] , , Банкина адсорб-ционных свойств мицелия базидиомицета Poliporus ciliatus. Молодой ученый. 2014. №18. С.116-120.
[3] , , Бриллиантова адсорбционных свойств мицелия базидиомицета Abortiporus biennis. Молодой ученый. 2014. №19. С.56-60.
[4] , , Дмитриева коллоидных свойств мицелия базидиомицетов с целью использования его как носителя БАВ. Бутлеровские сообщения. 2014. Т.39. №9. С.76-82 .
[5] , , Дмитриева электроповерхностных свойств мицелия базидиомицета Abortiporus biennis в зависимости от рН. Известия вузов. Прикладная химия и биотехнология. 2014. №5 (10). С.32-38.
[6] , , Зарембо переход пар – кристалл при синтезе пленок парацетамола методом вакуумного испарения и конденсации. Журнал технической физики. 2014. Т.84. №3. С.141-143.
[7] , , Беляева парацетамола ромбической формы. Бутлеровские сообщения. 2014. Т.37. №3. С.113-119.
[8] , , Бордей превращения при формировании кристаллов парацетамола из паровой фазы. Журнал технической физики.2014. Т.84. №7.С.156-158.
[9] , , О механизме начальной стадии образования наноструктур в условиях сверхнизких температур. Наносистемы: физика, химия, математика. 2012. Т.3. №5. С.103-110.
[10] , , Голикова устойчивость дисперсных систем. Часть 1. Исследование электроповерхностных свойств и кинетики коагуляции монодисперсных полимерных частиц с карбоксилированной поверхностью. Бутлеровские сообщения. 2014. Т.37. №2. С.29-38.
[11] , , Чухно свойства гемоглобина в водных растворах НСl и KCl. Бутлеровские сообщения. 2012. Т.30. №6. С.103-107.
[12] , , Чухно свойства гемоглобина в вод-ных растворах 1-, 2- и 3-зарядных ионов. Бутлеровские сообщения. 2013. Т.34. №6. С.135-140.
[13] , , Чухно свойства хлорофилла в вод-ных растворах 1-, 2- и 3-зарядных катионов. Бутлеровские сообщения. 2013. Т.34. №6. С.130-134.
[14] , , Назипова ионов Н+ и ОН - на хлорофилле, влияние рН на устойчивость водных дисперсий хлорофилла. Бутлеровские сообщения. 2013. Т.34. №5. С.124-130.
[15] , , Чухно состава дисперсионной среды на устойчивость и электрокинетические свойства билирубина. Бутлеровские сообщения. 2014. Т.37. №1. С.55-61.
[16] , , Тефц оценка действия поверхностно-активных веществ на изменение кинетических параметров агрегации тромбоцитов. Институт стоматологии. 2010. Т.3. № 48. С.80-81.
[17] , , Кучук физико-химических свойств водной дисперсии энтеросорбента полисорба мп. Бутлеровские сообщения. 2013. Т.35. №8. С.94-99.
[18] , , Мартынов солей одно-, двух - и трехзарядных катионов металлов на сорбцию Н+ и ОН—ионов на декстране. Бутлеровские сообщения. 2011. Т.27. №14. С.47-54.
[19] , , Мартынов точка белков в водных растворах азолов. Вестник СПбГУ. 2011. Серия 4: Физика. Химия. Вып.2. С.124-133.
[20] , , Мартынов ионов Н+ и ОН - на коллагене. Вестник СПбГУ. 2011. Серия 4: Физика. Химия. Вып.3. С. 87–95.
[21] , , . Изучение взаимодействия белков с биологически активными азотсодержащими гетероциклическими соединениями при различных значениях pH. Бутлеровские сообщения. 2013. Т.34. №5. С.91-99.
[22] , , . Кинетика процесса набухания желатины в водных растворах азолов. Бутлеровские сообщения. 2014. Т.38. №5. С.84-88.
[23] Men'shikova A. Yu., Skurkis Yu. O., Kuchuk V. I., Dmitrieva I. B., Evseeva T. G., Shabsel's B. M. Effect of the surface structure of poly(styrenecoacrolein) microspheres and its modification by protein on electrosurface properties. Коллоидный журнал. 2001. Т.63. №5. С.629-636.
[24] , , Чухно точки нулевого заряда молекул альбумина в водных дисперсиях при различных концентрациях хлорида калия. Молодой ученый. 2014. №18. С.123-126.
[25] , Кучук и коллоидная химия. Под. ред. . 2-ое изд., переработанное и дополненное. М.: ГЭОТАР-Меди. 2012. 752с.
[26] , , Алексеева условий культивирования и антиоксидантной активности мицелия базидиомицета Phallus impudicus (веселка обыкновенная). Проблемы медицинской микологии. 2014. Т.16. №3. С.80-82.
[27] , , Галынкин культивирования, состав и биологическая активность мицелия flammulina velutipes (fr.). P. Karst. Прикладная биохимия и микробиология. 2010. Т.46. №5. С.583-586.
[28] Григоров свойства капиллярных систем. , , М.-Л.: Изд-во АН СССР. 1956. 336с.
[29] , , . Физическая и коллоидная химия. Руководство к практическим занятиям: учебное пособие. Под ред. проф. . М.: ГЭОТАР-Меди. 2012. 320с.
[30] , , . Взаимодействие глицина с катионами железа(III) и никеля(II) в водных растворах и на поверхности их оксидов. Бутлеровские сообщения. 2013. Т.35. №8. С.133-137.
[31] Dmitriyeva I. B., Tikhomolova K. P., Chukhno A. S., Prokopovich P. P., Starov V. M. Investigation of the electrosurface properties of NiO and Fe2O3 in azole solutions. Colloids and Surfaces A: Physicochemical and Engineering Aspects. 2007. Т.300. №3 SPEC. ISS. P.315-320.
[32] , , Кергенцев свойства оксида кремния(IV) в водных растворах азолов. Бутлеровские сообщения.2014. Т.38. №5. С.78-83.
[33] , , Чухно адсорбции 1,3- диазола на поверхности оксидов NiО и Fe2О3. Журн. прикл. химии. 2005. Т.78. Вып.5. С.741-746.
[34] , , Чухно тетразола на оксидах Ni(II) и Fe(III). Журн. прикл. химии. 2006. Т.79. Вып.1. С.51-56.
