
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
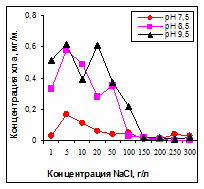 Рис. 1. Зависимость содержания
Рис. 1. Зависимость содержания
хлорофилла а от концентрации
NaCl и рН
Смена видов в сообществе происходила следующим образом: при минерализации 1-50 г/л (рН 7,5-9,5) доминировала Nodularia sp. Leptolyngbya sp. и D. salina встречались реже. В интервале 50-100 г/л происходила качественная и количественная смена сообщества с цианобактерий на эукариотические водоросли, сопровождающаяся снижением содержания хлорофилла а (рис. 1).
Наиболее благоприятными для роста цианобактерий являлись значения рН 8,5-9,5. рН 7,5 частично ограничивал рост, а в совокупности с высокой соленостью делал его невозможным.
Диапазон развития цианобактерий находился в пределах 1-50 г/л NaCl с оптимумом при 1-10 г/л. Уменьшение содержания хлорофилла а при солености более 100 г/л NaCl происходило на фоне высокой численности, но низкой, по сравнению с цианобактериями, биомассы эукариотных водорослей.
4.3.2. Анализ видового состава цианобактерий с помощью коэффициента флористической общности Жаккара
С помощью коэффициента флористической общности Жаккара (k) был проанализирован состав цианобактерий солоноватых и соленых озер Южного Забайкалья на видовом и родовом уровнях.
Анализ данных показал, что коэффициенты сходства варьируют в широких пределах (k = 0-0,75). Самые низкие коэффициенты сходства (k = 0-0,063) получены при сопоставлении видового и родового спектров цианобактерий, развивающихся в низких и высоко минерализованных озерах (солоноватые озера и рассолы, соответственно) (рис. 2 А, Б). Это свидетельствует о значительном морфологическом отличии цианобактериальных комплексов озер с пограничными значениями минерализации. Наибольшие коэффициенты сходства (0,75 для родов, 0,44 для видов) получены при сопоставлении таксономического спектра озер с близкими значениями минерализации, что объясняется присутствием значительного количества общих видов цианобактерий.
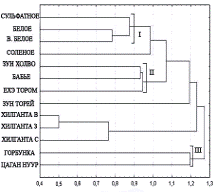
А
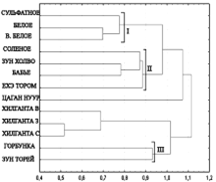
Б
Рис. 2. Дендрограмма сходства комплексов цианобактерий различных озер, построенная методом невзвешенного попарного арифметического среднего на основании коэффициента Жаккара, рассчитанного для видовых таксонов (А) и для родовых таксонов (Б). Расстояние Эвклидовое.
4.4. Функциональная активность продуцентов исследуемых озер
Скорости продукционных процессов в цианобактериальных матах
Скорость общего фотосинтеза в цианобактериальных матах озер Южного Забайкалья составляла 18,4-336,0 мг С/м2∙сут, темновой фиксации – 2,9-19,4 мг С/м2∙сут (табл. 4).
Таблица 4
Скорость продукционных процессов
в цианобактериальных матах озер (мг С/м2∙сут)
Озеро | Дата | М, г/л | Общий фотосинтез | Аноксигенный фотосинтез | Темновая фиксация | |
мг С/м2∙сут | % от продукции фотосинтеза | |||||
Борзинское | 08.2002 | 322 | 201,7-336,0 | 0 | 0 | 0 |
Хилганта | 07.2003 | 164 | 63,5 | 0 | 0 | 0 |
Бабье | 08.2002 | 85,1 | 50,7-123,7 | 0 | 0 | 0 |
Хилганта | 08.2001 | 82,0 | 756 | 538 | 71 | 41,4 |
Хилганта | 04.2004 | 30,0 | 18,4-22,9 | 0,46-0,52 | 2,0-2,8 | 5,02-8,56 |
Соленое | 09.2002 | 11,4 | 72,5-262,5 | 31,2-47,0 | 11,8-64,8 | н. о. |
Верхнее Белое | 06.2002 | 9,4 | 541,9 | 49,6 | 9,15 | 2,9 |
Соленое | 06.2004 | 8,0 | 78,4-81,5 | 2,4-3,3 | 3,06-4,04 | 11,4-19,4 |
Скорость аноксигенного фотосинтеза находилась в пределах 0,46-538,0 мг С/м2∙сут. Доля аноксигенного фотосинтеза в продукции органического вещества (ОВ) варьировала от 2 до 71%. Продуктивность матов исследованных озер Южного Забайкалья значительно уступает продуктивности матов озер Восточно-Африканской рифтовой зоны (11600 мг С/м2∙сут) (Grant et al., 1990). Это объясняется значительными колебаниями температуры в течение года и более коротким вегетационным периодом в озерах Южного Забайкалья, что не позволяет данным микробным сообществам развиваться и функционировать в постоянном режиме.
Сравнение данных по продукции цианобактериальных матов оз. Хилганта с литературными данными показало, что засушливые условия и низкая температура значительно снижают продуктивность сообщества (Намсараев, Намсараев, 2007). В летний период скорость общего фотосинтеза была максимальной при солености 32-41,5 г/л и составляла 3600-3860 мг С/м2∙сут (табл. 5). При повышении минерализации до 82-164 г/л она значительно снижалась (63,5-756 мг С/м2∙сут). Такая же тенденция наблюдалась и при понижении температуры.
Таблица 5
Продукция в цианобактериальных матах озера Хилганта (мг С/м2∙сут)
Дата | Т, оС | М, г/л | Общий фотосинтез | Темновая фиксация |
17.07.2003 | 30,0 | 164,0 | 63,5 | н. о. |
20.08.2001 | 26,0 | 82,0 | 756,0 | 41,40 |
21.07.1996" | 34,0 | 45,0 | 3840,0 | н. о. |
05.07.1995" | 28,0 | 41,5 | 3600,0 | н. о. |
08.08.1999" | 24,0 | 32,0 | 3860,0 | 26,27 |
07.04.2004 | 0,5 | 30,0 | 18,4 | 8,56 |
Здесь и далее: " - данные , (2007 г.).
Изотопный состав углерода цианобактериальных матов
Изотопный состав углерода d13С микробных матов и донных осадков был исследован в озерах Зун Торей, Белое, Соленое и Хилганта. Изотопный состав углерода ОВ в озерах варьировал от -14,80‰ до -30,63‰ (табл. 6). Значения d13С карбонатов микробных матов в исследованных озерах варьировали от -0,14 до -12,15‰.
В озере Хилганта изотопный состав углерода был исследован в разные годы. В водный период (1998 г.) в озере наблюдалось утяжеление изотопного состава углерода ОВ до -15,60‰. В засушливый период (2007-2008 гг.) отмечалось облегчение изотопного состава ОВ (-30,63‰). Соотношение 12С/13С углерода ОВ в нижних слоях цианобактериального мата в водный период было равно -14,8‰ (табл. 6). По-видимому, основная часть ОВ нижних слоев мата образуется в результате деструкции цианобактерий. Значения d13С карбонатов цианобактериальных матов в водный период был равен -0,14‰, в засушливый период – от 8,65‰ до 9,83‰.
Таблица 6
Изотопный состав углерода цианобактериальных матов озер Забайкалья (‰)
Озеро, толщина мата | Дата | d13СОВ | d13Скарб |
Хилганта", 0-2 см | 08.1998 | -15,60 | -0,14 |
2-4 см | « « | -14,8 | - |
Хилганта, 0-2 см | 08.2007 | -30,63 | -8,65 |
2-4 см | « « | -27,67 | -9,83 |
Хилганта | 08.2008 | -27,75 | -8,21 |
Зун Торей | 08.2008 | -27,90 | -9,20 |
Белое | 07.2008 | -19,48 | -12,05 |
Соленое | 08.2008 | -27,27 | -12,15 |
4.5. Экофизиология и систематическое положение цианобактерий
4.5.1. Филогенетическая характеристика цианобактерий
Из донных осадков и микробных матов исследованных озер было выделено шесть монокультур цианобактерий (табл. 7). Три культуры (Cya 1, 2, 10) отнесены к роду Phormidium, две культуры (Cya5, 6) – к роду Nodularia, одна культура (Cya 4) – к Pseudanabaena.
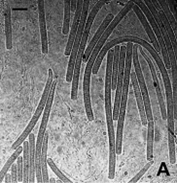
Культуры Cya 1 и Cya 2 были выделены из цианобактериальных матов озер Сульфатное и Ехэ Тором, но по морфологическим характеристикам, лишь с некоторыми вариациями в размерах, являлись идентичными Oscillatoria brevis (Еленкин, 1949) или Phormidium breve (Komárek & Anagnostidis, 2007) (рис. 3 А).
Таблица 7
Культуры цианобактерий, выделенные из исследованных озер
Культура | Озеро | Классификация по Еленкину (1949), Голлербах и др. (1953) | Классификация по Komárek&Anagnostidis (1999, 2007) | На основе сиквенса 16S РНК |
Cya 1 | Сульфатное | Oscillatoria brevis (Kűtz.) Gom. | Phormidium breve (Kűtz. et Gom.) | Phormidium sp. |
Cya 2 | Ехэ Тором | Oscillatoria brevis (Kűtz.) Gom. | Phormidium breve (Kűtz. et Gom.) | Phormidium sp. |
Cya 4 | Сульфатное | Phormidium frigidum Fritsch. | Pseudanabaena frigida (Fritsch) Anagnost. | - |
Cya 5 | Хилганта | Anabaena sibirica (Pop. et Degt.) Elenk. | Nodularia sp. | Nodularia sp. |
Cya 6 | Горбунка | Anabaena sibirica (Pop. et Degt.) Elenk. | Nodularia sp. | Nodularia sp. |
Cya 10 | Хилганта | Phormidium sp. | Phormidium sp. | Phormidium sp. |

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
