Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Свободнорадикальное окисление и его роль в системе «почва-растение»
Автор: Василевская Яна Юрьевна,11 класс, самарский медико-технический лицей, г. Самара, e-mail: *****@***ru
Научный руководитель: , доцент кафедры медицинских диагностических систем Самарского Государственного Аэрокосмического Университета (СГАУ), кандидат медицинских наук.
Содержание Стр.
Введение…………………………………………………………………………………………. 2 Глава I. Свободнорадикальные реакции и методы их изучения………………………….2
Первые шаги в изучении свободных радикалов……………………………………………2
Реакции с участием активных форм кислорода……………………………………………3
1.3 Свечение при реакциях цепного окисления липидов…………………………………..4
1.4 Хемилюминесценция в реакциях с участием окиси азота…………………………….6
1.5 Биохимические методы изучения реакций с участием
свободных радикалов……………………………………………………………………………8
Глава II. Растительные пигменты…………………………………………………………….9
Глава III. Биохимические процессы поглощения растениями азота…………………..11
Цель исследования……………………………………………………………………………..12
Задачи……………………………………………………………………………………………..12
Глава IV. Практическая часть…………………………………………………………………13
4.1 Методика и техника исследования………………………………………………………13
4.2 Объем исследования………………………………………………….…………………..13
4.3 Полученные результаты и их обсуждение…………………………………………….13
Выводы……………………………………………………………………... ……………………17
Заключение………………………………………………………………………………………18
Библиографический список………………………………………………. …………………18
Введение.
Реакции растений на химические стрессы, вызванные избытком химических элементов в окружающей среде, нельзя рассматривать как определенные раз и навсегда. Следует учитывать, что у растений в ходе процесса эволюции и в течение жизни индивида (онтогенез и филогенез) вырабатываются механизмы, приводящие к адаптации и нечувствительности к изменению и нарушению химического баланса в окружающей среде. Способность перерабатывать химические элементы у растений весьма изменчива. Поэтому реакции растений на изменение состава почвы должны всегда исследоваться для конкретной системы «почва-растение». [6, 7, 11, 13]
Гипотеза: В связи с накоплением химических веществ органического и неорганического характера в почве усиливается свободнорадикальное окисление (СРО), небезразличное для биологических структур, в частности, для мембран корневой системы. Меняющееся взаимоотношение в системе «почва-растение» сказывается на свойствах растений. Так, ионы металлов в условиях СРО изменяют свои токсические свойства. Но если у мембран корневой системы растений в основе гелевые структуры, которые могут изменять скорость химических реакций, то у почвы такие свойства выражены слабо. Усиление гелевых структур почвы представляет научный интерес как для выявления защитных механизмов от токсичных продуктов СРО, так и для изменения качества растений, в частности, избирательного накопления в них определенных органических структур.
Глава I. Свободнорадикальные реакции и методы их изучения.
1.1. Первые шаги в изучении свободных радикалов.
Известно, что в органических молекулах, включая те, из которых состоит организм человека, электроны на внешней электронной оболочке располагаются парами – одна пара на каждой орбитали. Свободные радикалы отличаются от обычных молекул тем, что у них на внешней электронной оболочке имеется неспаренный (одиночный) электрон. Можно дать следующее определение:
Свободные радикалы – это молекулярные частицы, имеющие неспаренный электрон на внешней электронной оболочке. Их изучение ведется методами ЭПР, хемилюминесценции и с помощью ингибиторов.
Первая экспериментальная работа, подтверждающая образование семихинонов - промежуточных продуктов одноэлектронных стадий, имеющих свободнорадикальный характер, принадлежит Хаасу, который обнаружил появление промежуточного окрашенного соединения фермент-субстратного комплекса свободно-радикального типа. Затем Теорелл, Чанс и ряд других авторов провели целый ряд важных работ по изучению быстрых одноэлектронных стадий в окислительно-восстановительных ферментативных процессах. Однако наиболее существенный шаг вперед был сделан в работе Коммонера, Таундсена и Пейка, в которой впервые зарегистрированы свободно-радикальные состояния в метаболизирующих тканях и клетках животного и растительного происхождения методом электронного парамагнитного резонанса (ЭПР). По амплитуде и форме сигналов (спектров) ЭПР можно судить о существовании непарных электронов в образце, определять их концентрацию, а иногда и выяснить, какова химическая структура радикалов, которые эти непарные электроны содержат. Не менее эффективным методом изучения реакций с участием радикалов оказался метод хемилюминесценции. Хемилюминесценцией называется свечение, сопровождающее химические реакции. Она наблюдается в том случае, если в реакции происходит выделение большого количества энергии, направленной в реакции взаимодействия двух радикалов или в реакциях с участием перекисей. Помимо этого слабым свечением сопровождается образование свободных радикалов при действии некоторых физических факторов на объект: при ионизирующей радиации наблюдается радиохемилюминесценция, после облучения ультрафиолетовым (УФ) или видимым светом – фотохемилюминесценция, при пропускании электрического тока – электролюминесценця, при действии ультразвука – сонохемилюминесценция, при воздействии сил трения – триболюминесценция. [3]
1.2 Реакции с участием активных форм кислорода.
Активными формами кислорода (АФК) обычно называют перекись водорода (Н2О2), гипохлорит (ClO-) и кислородные радикалы: супероксид и радикал гидроксил (НО.). Главным источником АФК в организме человека и животных служат клетки-фагоциты: гранулоциты и моноциты крови и тканевые макрофаги. Мембраны фагоцитов содержат ферментативный комплекс НАДФ–оксидазу, который окисляет НАДФ. Н2 до НАДФ+ за счет восстановления двух молекул кислорода до супероксидного радикала:
![]() НАДФ. Н2 + 2О2 НАДФ+ + 2О2
НАДФ. Н2 + 2О2 НАДФ+ + 2О2
При взаимодействии (диспропорционировании) двух супероксидных радикалов образуется перекись водорода и кислород:
 О2 + О2 + 2Н+ Н2О2 + О2
О2 + О2 + 2Н+ Н2О2 + О2
Фагоцит выделяет в окружающую среду не только АФК, но и несколько ферментов, среди которых важнейший – миелопероксидаза, катализирующая реакцию образования гипохлорита из аниона хлора и перекиси водорода:
 Н2О2 + Cl- Н2О + ClO-
Н2О2 + Cl- Н2О + ClO-
Кроме того, в присутствие ионов металлов переменной валентности, например железа, происходит образование радикалов гидроксила из перекиси водорода (реакция Фентона) и гипохлорита (реакция Осипова):
 Н2О2 + Fe2+ Fe3+ + НО - + НО.
Н2О2 + Fe2+ Fe3+ + НО - + НО.
 ClO - + Fe2+ Fe3+ + Cl - + HO.
ClO - + Fe2+ Fe3+ + Cl - + HO.
Эти реакции приводят к печальным последствиям для окружающих клеток. Радикал гидроксила чрезвычайно активен химически и разрушает почти любую встретившуюся ему молекулу. Действуя на SH-группы, гистидиновые и другие аминокислотные остатки белков, НО. вызывает денатурацию последних и инактивирует ферменты. В нуклеиновых кислотах НО. разрушает углеводные мостики между нуклеотидами и, таким образом, разрывает цепи ДНК и РНК, в результате чего происходят мутации и гибель клеток. Внедряясь в липидный слой клеточных мембран, радикал гидроксила запускает реакции цепного окисления липидов, что приводит к повреждению мембран, нарушению их функций и гибели клеток. Таким образом, радикал НО. - это радикал-разрушитель.
Таким образом, активированные фагоциты для борьбы с чужеродными клетками образуют большое количество АФК, которые могут при взаимодействии друг с другом и с другими молекулами испускать кванты света в виде хемилюминесценции.
1.3 Свечение при реакциях цепного окисления липидов.
Одна из главных составляющих собственной (неактивированной) хемилюминесценции животных клеток и тканей – свечение, сопровождающее цепное окисление липидов в мембранных структурах клеток и липопротеинах крови. Эта реакция идет с участием свободных радикалов липидов (L.) и липопероксидов (LOO.), которые образуют цепи окисления.
Реакция цепного окисления липидов.
 НО. LH L. LOO. L.
НО. LH L. LOO. L.
HOH LOOH
Время от времени радикалы, ведущие цепь окисления, взаимодействуют друг с другом. В реакции взаимодействия двух радикалов липопероксида (LOO.) образуются молекулы кетона и кислорода в электронно-возбужденном состоянии, которые затем переходят в основное состояние, испуская квант света (фотон).
Чем больше радикалов LOO. в системе, т. е., чем энергичнее идут цепные реакции окисления липидов, тем выше интенсивность хемилюминесценции, сопровождающей реакцию радикалов. Вещества, реагирующие со свободными радикалами и тем самым тормозящие цепное окисление липидов (антиоксиданты), одновременно подавляют хемилюминесценцию. Именно подавление собственной хемилюминесценции тканей и клеток такими антиоксидантами, как, например, токоферол (витамин Е), указывает на то, что это свечение обусловлено реакциями цепного окисления липидов. С другой стороны, изучая влияние различных природных и синтетических соединений на кинетику ХЛ, можно судить о способности этих веществ защищать организм от вредного действия свободных радикалов и тем самым отбирать кандидатов на определенные лекарства.
Увеличенное образование свободных радикалов в организме, которое иногда называют оксидативным стрессом, и связанное с этим усиление процессов пероксидации липидов сопровождаются нарушениями в свойствах биологических мембран и функционировании клеток. Наиболее изучены три основных следствия перекисного окисления липидов.
Первое: перекисное окисление липидов сопровождается окислением тиоловых (сульфгидрильных) групп мембранных белков (Pr). Это может приводить к неферментативной реакции SH-групп со свободными радикалами липидов. При этом образуются сульфгидрильные радикалы, которые затем взаимодействуют с образованием дисульфидов или окисляются кислородом с образованием производных сульфоновой кислоты:
![]() Pr-SH + L. LH + Pr-S.,
Pr-SH + L. LH + Pr-S.,
![]() Pr1-S. + Pr2-S. Pr1-SS-Pr2,
Pr1-S. + Pr2-S. Pr1-SS-Pr2,

![]() Pr1-S. + O2 Pr-SO.2 молекулярные производные.
Pr1-S. + O2 Pr-SO.2 молекулярные производные.
Большую роль в патологии клетки играет также инактивация ион-транспортных ферментов, в активный центр которых входят тиоловые группы, в первую очередь Са-АТФазы[4]. В числе причин поражения Са-АТФазы у больных с гипертонической болезнью называют активацию процессов с участием свободных радикалов. Так в модельных опытах с изолированными везикулами саркоплазматического ретикулума было показано, что Са-АТФаза очень чувствительна к перекисному окислению липидов, при котором происходит окисление SH-групп, входящих в активный центр фермента. Са-АТФаза, перестав качать ионы кальция; превращается из насоса в канал для кальция, через который эти ионы начинают переноситься не из цитозоля в ретикулум, как им полагается, а, наоборот, из ретикулума, где их концентрация выше, в цитоплазму, где их концентрация ниже. Подобного типа повреждение кальциевых насосов происходит нередко. Хорошо известна роль свободных радикалов в развитии дегенеративных болезней, включая рак, многие интоксикации, болезни, связанные с атеросклерозом и иммунными нарушениями. Во многих случаях повреждение Са-АТФаз свободными радикалами может играть не последнюю роль в зарождении и развитии заболевания. Наконец, окисление тиоловых групп мембранных белков приводит к появлению дефектов в липидном слое мембран клеток и митохондрий. Под действием разности электрических потенциалов на мембранах через такие поры в клетки входят ионы натрия, а в митохондрии – ионы калия. В результате происходят увеличение осмотического давления внутри клеток и митохондрий и их набухание. Это приводит к еще большему повреждению мембран.
Второй результат перекисного окисления липидов связан с тем, что продукты пероксидации обладают способностью непосредственно увеличивать ионную проницаемость липидного бислоя. Так, показано, что продукты перекисного окисления липидов делают липидную фазу мембран проницаемой для ионов водорода и кальция. Это приводит к потере митохондриями способности осуществлять синтез АТФ, и клетка оказывается в условиях энергетического голода. Одновременно в цитоплазму выходят ионы кальция, которые повреждают клеточные структуры.
Третий (возможно, самый важный) результат пероксидации – это уменьшение стабильности липидного слоя, что может привести к электрическому пробою мембраны собственным мембранным потенциалом, то есть под действием разности электрических потенциалов, существующей на мембранах живой клетки. Электрический пробой приводит к полной потере мембраной её барьерных функций.
В нормальных условиях процесс перекисного окисления липидов находится под строгим контролем ферментативных и неферментативных систем клетки, отчего скорость его невелика. Принято делить химические соединения и физические воздействия, влияющие на скорость перекисного окисления липидов, на прооксиданты, усиливающие процессы пероксидации, и антиоксиданты, тормозящие процессы перекисного окисления липидов.
1.4 Хемилюминесценция в реакциях с участием окиси азота.
Окись азота (NO) – это газ, хорошо растворимый в воде и обладающий высокой реакционной способностью. Последнее связано с тем, что NO – свободный радикал (NO.). Любопытно, что само открытие выделения окиси азота клетками человека и животных было сделано в 1984 году методом хемилюминесценции, правда, не в растворе, а газовой фазе. При смешивании окиси азота и озона наблюдается яркая хемилюминесценция, и в 80-е годы газовые хемилюминометры для определения окиси азота выпускались в промышленном масштабе. К концу 80-х годов биологическая роль окиси азота уже ни для кого не была секретом. Его первая расшифрованная функция – расслабление стенок кровеносных сосудов (вазодилатация), которое, в свою очередь, приводит к улучшению кровоснабжения органов и снижению общего кровяного давления. Вскоре оказалось, что NO выделяется клетками-фагоцитами и участвует в борьбе организма-хозяина с микроорганизмами. Сейчас хорошо известно, что окись азота выделяется очень многими типами клеток и является одним из основных регуляторов внутриклеточных процессов.
Участие нитроксида в собственной хемилюминесценции тканей животных было показано в опытах Джулио Терренса и его сотрудников, которые изучали свечение перфузируемого легкого. Оказалось, что свечение снижается на 85% при введении в перфузат нитро-L-аргинина, ингибитора NO-синтетазы - фермента, катализирующего образование окиси азота в живых клетках. Свечение удавалось наблюдать также в растворах, содержащих окись азота, супероксидные радикалы и какой-нибудь белок. Известно, что при реакции окиси азота и супероксида образуется пероксинитрит - токсичное соединение. По-видимому, именно реакция пероксинитрита с белком приводит к свечению, которое вносит существенный вклад в хемилюминисценцию всего органа.
Надо заметить, что природа процессов, определяющих собственное свечение тканей, может меняться при изменении состояния этой ткани. В опытах того же автора было показано, что у животных с воспалением легких ингибитор NO-синтетазы слабо влиял на свечение органа, зато свечение снижалось антиоксидантами – супероксиддисмутазой и ловушками липидных радикалов. Логичен сделанный вывод, что при воспалении на первый план выходят реакции, связанные с активацией клеток-фагоцитов и образованием активных форм кислорода, а затем липидных перекисей, тогда как в норме за свечение ответственны реакции окиси азота.
Хемилюминесценция, сопровождающая реакции свободных радикалов характеризуется низкой интенсивностью свечения. Существует целых три причины, объясняющие это. Во-первых, сама концентрация радикалов в биологических системах очень мала из-за высокой химической активности, поэтому малы и скорости реакций, сопровождающихся свечением. Во-вторых, не всякое химическое взаимодействие радикалов непременно приводит к образованию электронно-возбужденных молекул продуктов реакции. Напротив, в подавляющем большинстве окислительно-восстановительных взаимодействий между молекулами или радикалами электрон переносится не на уровень возбужденного состояния, а на самый нижний свободный уровень и последующего высвечивания кванта не происходит. В-третьих, даже если и образовалась возбужденная молекула продукта, вероятность того, что высветится квант, а не произойдет растрата энергии в тепло, тоже обычно очень мала. Две последние причины приводят к тому, что квантовый выход хемилюминесценции в случае реакции двух перекисных радикалов всего 10-8 – 10-10. Это происходит потому, что квантовый выход образования возбужденных молекул продукта равен всего 10-4 -10-5, а квантовый выход люминесценции продукта составляет для кетонов, образующихся при взаимодействии перекисных радикалов, в свою очередь около10-4 -10-5 . Выходит, что общий квантовый выход хемилюминесценции составляет всего лишь Qхл. = Qвозб. ´ Qлюм. = 10-8 -10-10. Отсюда понятна стратегия для использования хемилюминесценции как метода обнаружения радикалов.
1.5 Биохимические методы изучения реакций с участием свободных радикалов.
Такие методы изучения свободнорадикальных реакций, как метод хемилюминесценции и метод электронного парамагнитного резонанса относятся к методам биофизики. Но существуют также биохимические методы изучения реакций, в которых участвуют свободные радикалы. Радикалы обладают высокой реакционной способностью, и изучать их обычными химическими методами невозможно. Важную роль при решении таких вопросов играет так называемый ингибиторный анализ. Классическим примером может служить применение фермента супероксиддисмутазы (СОД). Этот фермент, открытый Дж. Мак-Кордом и И. Фридовичем около тридцати лет назад, катализирует реакцию взаимодействия (дисмутации) двух супероксидных радикалов с образованием перекиси водорода и молекулярного кислорода. Открытие СОД совершило революцию в умах биохимиков: если есть фермент, удаляющий кислородные радикалы и специально вырабатываемый живыми клетками (и, как выяснилось, широко распространенный в живой природе), то ясно, что и сами радикалы существуют в природе и почему-то их надо обязательно удалять. Постепенно СОД стали широко использовать во всех исследованиях, где изучают роль супероксида в том или ином процессе, будь то индивидуальная биохимическая реакция или развитие болезни у лабораторных животных или человека. Если добавление СОД тормозит изучаемый процесс, значит, для его протекания необходим супероксид-радикал. Остается выяснить, в какой именно реакции этот радикал участвует. Ингибиторный анализ используют и для изучения реакций с участием других радикалов. Так, для выяснения участия в каком-нибудь процессе реакций цепного окисления липидов используют жирорастворимые ловушки липидных радикалов, которые «ведут» цепи окисления. К числу таких ловушек относятся токоферол (витамин Е) и некоторые синтетические соединения, например трет-бутилгидрокситолуол (ионол). Водорастворимые радикалы эффективно перехватываются аскорбиновой или мочевой кислотой. Для улавливания радикалов гидроксила (НО.) используют маннитол или бензойную кислоту, а иногда этанол. Надо, однако, сказать, что далеко не всегда ловушки специфичны: многие из них реагируют не только с радикалами, но и с достаточно активными молекулами.
Глава II. Растительные пигменты.
Изучение оптических свойств листьев и поглощения ими света имеет большое значение для понимания общих принципов усвоения солнечной энергии, механизмов фотосинтеза и адаптационных процессов в растениях. Необходимо также отметить, что изучение этих механизмов очень важно при интерпретации данных, полученных с помощью дистанционного оптического зондирования.
Поглощение квантов света в видимой части спектра (400-700 нм), которые имеют достаточную энергию для осуществления фотосинтеза, обладают органические соединения (пигменты), содержащие системы сопряженных связей. В клетках эволюционно более древних фотосинтезирующих организмов (бактерии, водоросли) присутствуют разнообразные соединения (хлорофиллы, бактериофиллы, бактериородопсин, каротиноиды, фикобилины), способные при определенном их сочетении обеспечить акцептирование солнечного излучения во всей видимой (и даже в ближней инфракрасной) части спектра. Их набор и соотношение специфичны для различных групп и во многом зависят от условий обитания организмов. Пигменты фотосинтеза у высших растений значительно менее разнообразны.
В листьях (хлоропластах) высших растений присутствуют два хлорофилла (a и b), представляющие собой Mg-содержащие порфирины. Основная их часть включена в состав светособирающих комплексов, обеспечивает поглощение и передачу световой энергии на так называемые реакционные центры. В этих центрах, которые содержат лишь небольшую часть общего хлорофилла а, и осуществляются первичные фотосинтетические реакции. Кроме хлорофиллов в фотосинтетических мембранах всегда присутствуют желтые пигменты - каротиноиды (как правило, пять типов). Каротиноиды выступают также в роли светосборщиков и наряду с этим играют важную роль в защите фотосинтетического аппарата от фотоповреждений. В растворах как хлорофиллы, так и каротиноиды обладают интенсивными, узкими и достаточно далеко отстоящими друг от друга полосами поглощения света. Хлорофиллы эффективно поглощают синий, красный и очень слабо - зеленый свет. Спектры пигментов существенно уширяются и в значительно большей степени перекрываются в хлоропластах, которые можно легко выделить из ткани листа. Это связано с тем, что в отличие от растворов в хлоропластных мембранах пигменты ассоциированы с белками, липидами и взаимодействуют друг с другом. В результате этого достигается передача (миграция) энергии от светосборщиков на реакционные центры [10]. Еще более сглажены и сложны спектры поглощения листьев.
Часть света, падающего на лист, отражается от него. Различают зеркальное (на гладких, блестящих поверхностях) и диффузное отражения. Отражение листьев определяется главным образом диффузным отражением. Свет, проникая в толщу листа, сильно рассеивается и многократно изменяет свое направление. Это связано с тем, что как на поверхности, так и в толще листа существует множество поверхностей раздела, обладающих различными коэффициентами преломления (кутикула, эпидермис, клетки паренхимы, заполненные воздухом межклетники). Сходные эффекты происходят при прохождении света через клетки (различия в коэффициентах преломления между воздухом, жидкой фазой, клеточными стенками, рассеивание на субклеточных структурах). В результате при отсутствии сильного поглощения оптические пути света значительно увеличены по сравнению с геометрической толщиной листа. Благодаря этому листья поглощают значительно больше света, чем экстрагированные из них пигменты или изолированные хлоропласты [10].
Лучи разного цвета (длины волны) поглощаются неодинаково. Зеленые листья обладают очень высоким поглощением (низким отражением и пропусканием) в фиолетовой, синей, голубой и красной частях спектра. В этих областях поглощение мало зависит от концентрации хлорофиллов, что объяснятся тем, что в сильно рассеивающей среде и при высокой концентрации пигментов свет практически полностью поглощается уже у поверхности листа. Значительно большая часть излучения отражается и проходит через лист в области 520-580 нм, именно поэтому лист выглядит зеленым. Таким образом, зеленый свет, несмотря на низкие коэффициенты удельного поглощения хлорофилла в этой спектральной области, эффективно поглощается и достаточно равномерно заполняет ткань листа. Это имеет большое значение для фотосинтеза теневых, расположенных внутри кроны листьев, поскольку свет, отраженный или прошедший через расположенные выше листья, обогащен зелеными лучами [7, 10]. Интересно, что максимум спектральной чувствительности зрения у человека и многих наземных позвоночных расположен именно в зеленой области. Это, вероятно, является следствием того, что эволюция животных протекала в окружении растений, и можно понять, почему животные в своей жизнедеятельности используют эту часть солнечного излучения.
Глава III. Биохимические процессы поглощения растениями азота.
Среди органогенных элементов азот играет одну из важных ролей в жизни растений, несмотря на то, что в сухой массе растительных тканей его содержится всего 1–3%. Невысокая урожайность многих сельскохозяйственных культур чаще всего определяется недостатком именно азота. Для формирования урожая зерновых культур 2–3 т с одного гектара необходимо 150–200 кг азота в доступной для растений минеральной форме при общем содержании его в почве от 5 до 15 т на 1 га. При недостатке азота в среде обитания тормозится рост растений, ослабляется образование боковых побегов и кущение у злаков, наблюдается мелколистность.
Одно из ранних проявлений азотного дефицита – бледно-зеленая окраска листьев, вызванная ослаблением синтеза хлорофилла. Длительное азотное голодание ведет к гидролизу белков и разрушению хлорофилла прежде всего в нижних, более старых листьях и оттоку растворимых соединений азота к более молодым листьям. Вследствие разрушения хлорофилла окраска нижних листьев в зависимости от вида растения приобретает желтые, оранжевые или красные тона, а при сильно выраженном азотном дефиците возможно появление некрозов (омертвление). Азотное голодание приводит к сокращению периода вегетативного роста и более раннему созреванию семян. Вот почему вопрос об источниках азота для растений представляет большой интерес.
Растения потребляют в основном почвенный азот, причем даже в том случае, когда в почву вносятся высокие дозы минерального азота, т. к. это способствует минерализации органического вещества почвы.
Процесс превращения органического азота почвы в NН4+ носит название аммонификации. Он осуществляется гетеротрофными микроорганизмами (питаются готовыми органическими веществами) и схематически может быть представлен следующим образом:
органический азот почв RNН2 + СО2 + побочные продукты,
RNН2 + Н2О = NH3 + RОН,
NН3 + Н2О = NH4+ + ОH–.
Биологическое окисление NН3 или NH4+ до NO3– называется нитрификацией.
Еще в 1870 г. Шлезинг и Мюнц доказали опытным путем биологическую природу нитрификации. Для этого они добавляли к сточным водам хлороформ, в результате чего прекращалось окисление аммиака. Основная заслуга в раскрытии механизма нитрификации и выделении осуществляющих этот процесс микроорганизмов принадлежит выдающемуся российскому микробиологу С. Н.Виноградскому. В 1889 г. он открыл бактерии нитрификации и определил две фазы этого процесса.
Бактерии первой фазы окисляют аммиак до азотистой кислоты:
2NH3 + 3O2 = 2HNO2 + 2H2O.
В ходе второй фазы образовавшаяся азотистая кислота окисляется до азотной:
2HNO2 + О2 = 2НNO3
Превращение молекулы азота в аммиачную форму происходит следующим образом. Молекула азота двухатомна, связь между атомами тройная, и поэтому энергия термической диссоциации молекулы N2 велика (941,64 кДж/моль). Для разрыва этих связей, например при получении аммиака, требуется давление около 30 МПа, температура приблизительно 500 °С и катализатор, состоящий из Fe, активированного К2О, Аl2O3, СаО и др.
Биологическая фиксация N2 происходит при нормальных температуре и давлении благодаря высокой эффективности ферментативной системы данных микроорганизмов – нитрогеназе. Последняя состоит из Мо-Fe-белка (собственно нитрогеназа) и Fе-белка (редуктаза нитрогеназы). Активностью обладает лишь комплекс обоих компонентов.
Молекулярный азот связывается и восстанавливается на Мо-Fe-белке, а Fe-белок служит источником электронов для этого процесса. Процессу биологической азотфиксации также необходим постоянный приток энергии в форме АТФ (аденозинтрифосфорная кислота). Это обеспечивается благодаря дыханию и брожению, происходящим в микроорганизмах или в результате фотосинтеза. [1].
В связи с изложенным были определены цели и поставлены задачи исследования:
Целью исследования было изучение влияния СРО в системе «почва-растение» на биологические показатели растений, в частности, антиоксидантную защиту, и степень накопления в них органических веществ.
Задачи:
1. Выявить роль СРО в системе «почва-растение» в характере развития растений и накопления в них органических веществ. Исследовать антиоксидантную характеристику растений в этих условиях.
2. Определить характер изменений в растениях при смещении в системе «почва-растение» скорости химических реакций за счет увеличения в почве гелевых структур.
3. Показать оптимальные условия для усвоения азота растениями при повышении СРО в почве и изменении звена «почва-растение» за счет гелевых структур.
Глава IV. Практическая часть.
4.1 Методика и техника исследования.
Использовались зерна овса, которые высаживались
*в обычную почву и поливались водой (1 серия),
*в обычную почву и поливались 1% раствором Н2О2 (2 серия),
*в почву «гель+азот» и поливались водой (3 серия),
*в почву «гель+азот» и поливались 1% раствором Н2О2 (3 серия).
На десятый день после посадки зерен у них определялись и регистрировались:
v всхожесть (подсчет количества зерен, давших всходы из 100 посаженных; в %.),
v интенсивность роста (измерение длины ростков; в см),
v вес зеленой массы (в г.)
v оптическая плотность водных экстрактов в зависимости от длины волны
v антиоксидантная способность водных экстрактов (в мл 0,1% КмnO4 на 100 мл водной вытяжки из листьев)
4.2 Объем исследования.
Были проделаны 4 серии опытов, в каждом из которых проводились 5 повторов всех исследований.
4.3 Полученные результаты и их обсуждение.
Через 10 дней после посадки растения дали показатели, отраженные в табл. №1 на диаграммах 1a,1b,1c.
Таблица1
Н2О | Н2О2 | Н2О+азот | Н2О2+азот | |
длина (см) | 12 | 14 | 13 | 13,5 |
всхожесть (%) | 60 | 80 | 70 | 85 |
масса (г на 100 рост.) | 10,0 | 13,0 | 12,0 | 17,0 |
Диаграмма 1a
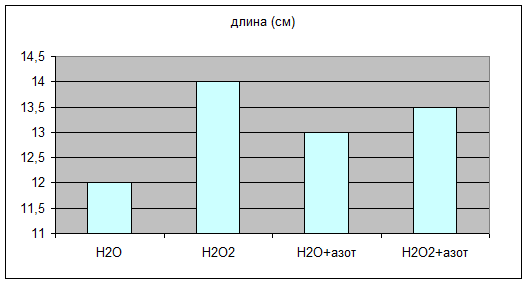 |
Диаграмма 1 b
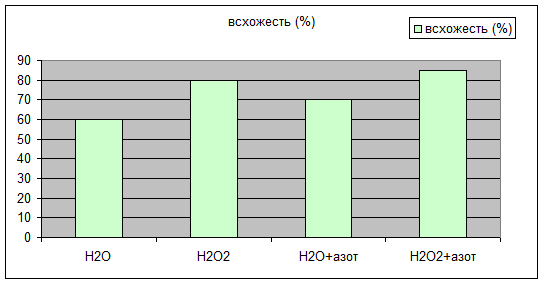 |
Диаграмма 1 c
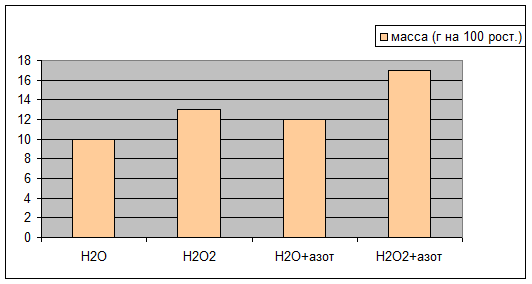 |
Просматривается четкая зависимость увеличения всхожести и зеленой массы овса в присутствии Н2О2 , а гелевые структуры усиливают этот эффект. Сканированием светопоглощения водной вытяжки из зеленой массы на КФК-3 были получены данные табл.№2 Графически это отражено на диаграмме 2a.
Таблица 2
длина волны (в нм) | оптическая плотность (D) | |||
Н2О | Н2О2 | Н2О+азот | Н2О2+азот | |
650 | 0,380 | 0,510 | 0,440 | 0,444 |
550 | 0,326 | 0,489 | 0,390 | 0,420 |
450 | 0,721 | 0,859 | 0,799 | 0,732 |
Диаграмма 2a
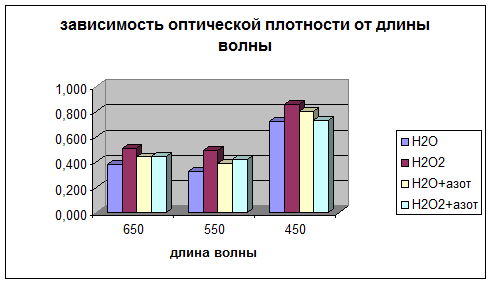
На основе полученных результатов была выявлена зона поглощения для хлорофилла (650 и 550 нм) и каротиноидов (450 нм). При этом масса хлорофилла увеличилась при усилении СРО и гелевых образований, а каротиноидов только за счет СРО.
Результаты исследования антиоксидантной активности вытяжки из зеленой части растений представлены в табл. №3 и на диаграмме 3a.
Таблица 3
Н2О | Н2О2 | Н2О+азот | Н2О2+азот | |
антиоксидант. спос-ть | 12 | 10 | 14 | 16 |
(в мл 0,1% KmnO4 на 100 мл) |
Диаграмма 3a
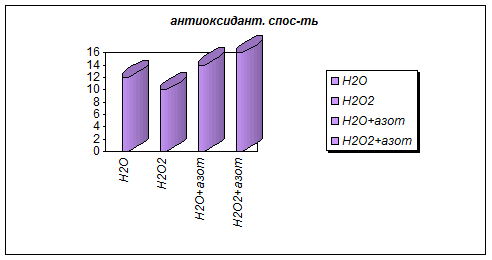
Они показывают, что обычные условия при увеличении азота не снимают стресса растений при СРО. Н2О2 снижает этот эффект, а гелевые структуры почвы усиливают его.
На основе всех полученных результатов можно сделать следующие выводы:
1. СРО в системе «почва-растение» стимулирует многие процессы в растениях: увеличивают массу, накапливают биологически активные пигменты.
2. Изначально формируемые в условиях повышенного СРО растения имеют меньший запас антиоксидантов, но азот способствует повышению антиоксидантных свойств растений.
3. Гелевые формы фиксации азота снижают эффект СРО, но усиливают процессы по сравнению с обычными структурами.
4. Регулируя СРО, а также скорость химических реакций за счет гелевых структур можно целенаправленно менять качество растений.
Заключение.
Практическое применение полученных данных.
Полученные результаты свидетельствуют о том, что регулируя процессы свободнорадикального окисления и скорость химических реакций можно менять качество растений, в частности, повысить их антиоксидантные свойства. Эти данные могут найти практическое применение в медицине (например, при изготовлении препаратов-антиоксидантов, полученных из растительных экстрактов). При этом использование особой гелевой формы азота в качестве почвенной добавки вполне оправдано, т. к. если у мембран корневой системы растений в основе гелевые структуры, которые могут изменять скорость химических реакций, то у почвы такие свойства выражены слабо. Поэтому, усиление таких гелевых структур способствует выявлению защитных механизмов растений от токсичных продуктов свободнорадикального окисления. Это очень актуально в связи с химическим загрязнением почвы.
Библиографический список.
1. А. А.Анисимова. «Основы биохимии» М.: Высшая школа, 1986.
2. Т. Роль неорганических ион-радикалов в органических и неорганических реакциях
// Соросовский Образовательный Журнал. – 1999. - №1. – с. 28-34.
3. А. Свечение, сопровождающее биохимические реакции // Соросовский Образовательный Журнал.1999 №6. с.25-32
4. А. Кальциевые насосы живой клетки // Соросовский Образовательный Журнал. 1998. №3. С. 20-27
5. А., а., С. и др. Общая химия. Биофизическая химия. Химия биогенных элементов // Учебное пособие для студентов медицинских спец. Вузов (под редакцией Ю. А. Ершова) – М.: Высшая школа. – 1993. – с.204-205.
6. А. Физиология растений (факультативный курс). – М.: Просвещение. – 1988.
7. И.Актичные формы кислорода и оксидативная модификация макромолекул: польза, вред и защита // Соросовский Образовательный Журнал. – 1999. - №1. – с. 2-7, 11-16.
8. Экология растений. М.: Мир, 1978. 186 с.
9. Н. Экология растений с основами биогеоценологии // Пособие для учителя. – М.: Просвещение. – 1978. – 270с
10. В. Ион-радикалы в органическом синтезе М.: Химия, 19861.
11. Фотосинтез: (Физиол. аспекты). М.: Мир, 1972. 315 c.
 Основные порталы (построено редакторами)
Основные порталы (построено редакторами)