Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Довольно значительны также половые различия в деятельности коры головного мозга. Так, мужчины лучше решают в уме пространственные задачи, легче выбирают маршруты пути. Женщины точнее выражают свои мысли словами, быстрее воспринимают изменения в окружающей обстановке.
Вся жизнедеятельность человека находится под контролем коры больших полушарий. Информация обо всем, что происходит в организме или вокруг него, в итоге обязательно попадает в кору. Таким образом, кора больших полушарий обеспечивает взаимодействие организма с окружающей средой и является материальной базой для психической деятельности человека.
Основной принцип деятельности ЦНС рефлекторный.
Рефлекс – это целостная стереотипная реакция организма на изменения внешней или внутренней среды, которая осуществляется при обязательном участии ЦНС. Путь, по которому осуществляется рефлекс, называется рефлекторной дугой. Она включает нейронную сеть, состоящую из афферентных, вставочных и эфферентных нейронов, соединённых посредством синапсов.При раздражении рецепторов рефлекторный акт возникает не мгновенно, а через некоторое время – латентный период рефлекса, который состоит из:
1) периода, необходимого для возбуждения рецептора и прохождения импульса по афферентной части рефлекторной дуги;
2) распространения возбуждения по нервному центру;
3) прохождения возбуждения по эфферентному нерву;
В основном время рефлекса определяется периодом распространения возбуждения через систему синапсов нервного центра, и чем сложнее рефлекторная дуга (рис.1, рис.2), тем больше время рефлекса. Время рефлекса также определяется особенностями нервных волокон (по которым осуществляется рефлекс), силой и временем прилагаемого раздражения, а также
площадью рецептивного поля, подвергаемого раздражению.
|

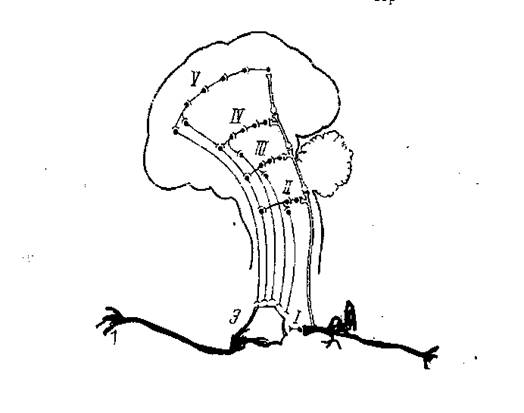
Рис.2. Схема многоуровневой (многоэтажной) рефлекторной дуги по А. Э. Асратяну.
А – афферентный сигнал; Э – эфферентный сигнал;
I – спинальный отдел; II – бульбарный отдел; III – мезенцефалический отдел; IV – диэнцефалический откел; V – корковый отдел.
КЛАССИФИКАЦИЯ РЕФЛЕКСОВ
I. ПО ПРОИСХОЖДЕНИЮ:
1. Безусловные, врожденные - видовые.
2. Условные - приобретенные в процессе индивидуальной жизни, выработанные при определенных условиях.
II. ПО БИОЛОГИЧЕСКОМУ ЗНАЧЕНИЮ:
1. Оборонительные, или защитные.
2. Пищевые – обеспечивающие добывание пищи, потребление и переваривание.
3. Половые – связанные с продолжением рода.
4. Ориентировочные – связанные с движением головы и глаз в сторону нового раздражителя.
5. Позно-тонические – рефлексы положения тела в пространстве.
6. Локомоторные рефлексы – перемещение тела в пространстве.
III. В ЗАВИСИМОСТИ ОТ РАСПОЛОЖЕНИЯ РЕЦЕПТОРОВ, В ОТВЕТ НА РАЗДРАЖЕНИЕ КОТОРЫХ ВОЗНИКАЕТ РЕФЛЕКС:
1. Экстероцептивные – рецепторы поверхности кожи.
2. Проприоцептивные – раздражение рецепторов мышц и сухожилий
3. Висцероцептивные – раздражение рецепторов внутренних органов.
IV. ПО ХАРАКТЕРУ ОТВЕТНЫХ РЕАКЦИЙ:
1. Секреторные – выражающиеся в выделении сока, выработанного человекам.
2. Трофические – связанные с изменением обмена веществ.
3. Двигательные – характеризующиеся сократительной деятельностью мышц (сгибательный, зрачковый, мигательный, сосудо-двигательный).
V. ПО УРОВНЮ ЗАМЫКАНИЯ - КАКИЕ ОТДЕЛЫ ЦНС УЧАСТВУЮТ В ИХ ОСУЩЕСТВЛЕНИИ:
1. Рефлексы спинного мозга – спинальные.
2. Рефлексы продолговатого мозга – бульбарные.
3. Рефлексы среднего мозга – мезенцефелические.
4. Рефлексы промежуточного мозга – диенцефалические.
5. Рефлексы коры больших полушарий – корковые.
НЕРВНЫЕ ЦЕНТРЫ.
В соответствии классической традицией представление о нервных центрах рефлексов составляет сердцевину всей рефлекторной теории. Под нервным центром понимают функциональное объединение нейронов, участвующих в осуществлении рефлекторного акта. Они возбуждаются притоком афферентной информации и адресуют свою активность эфферентным нейронам. Нервные центры функциональные, а не анатомические объединения нейронов, т. к. способны участвовать в замыкании нескольких рефлекторных дуг. Свойства нервных центров определяются свойствами центральных синапсов, поэтому в нервных центрах происходит односторонняя передача возбуждения, задержка проведения возбуждения и т. д.
Способность суммировать возбуждение присуща всем возбудимым тканям, однако наиболее ярко этот процесс выражен в нервных центрах.
Свойства нервных центров:
1. центральная задержка рефлекса. Объясняется явлением задержки проведения возбуждения в синапсах (синаптической задержкой), чем больше синапсов в нервном центре, тем дольше центральное время рефлекса;
2. пространственная и временная суммация возбуждения и торможения. Явление суммации возбуждения в ЦНС открыл И. М.Сеченов (1868). Временная суммация развивается при возбуждении одних и тех же рецепторов одного рецептивного поля с частотой импульсов меньшей, чем продолжительность ВПСП. ВПСП суммируются, и сила рефлекторной реакции возрастает. Пространственная суммация возникает при раздражении различных рецепторов рецептивного поля, при это суммация ВПСП происходит в разных синапсах нейрона и усиливает рефлекторный ответ. Благодаря суммации ВПСП возможно возникновение рефлекса на подпороговые раздражения.
3. спонтанная или фоновая активность нервных центров (тонус). Объясняется: а) спонтанной активностью нейронов; б) поступлением афферентных импульсов от других рефлексогенных зон; в) действие гуморальных факторов – изменением уровня гормонов, медиаторов, метаболитов; г) циркуляцией импульсов в ЦНС.
4. высокая чувствительность к недостатку О2, фармакологическим веществам и к изменению внутренней среды (уровня глюкозы, t°С и т. д.). Прекращение кровотока всего лишь на 10с приводит к потере сознания, а на 8-12мин приводит к гибели нейронов, прежде всего корковых.
5. утомляемость нервных центров. Была продемонстрирована Н. Е.Введенским. Она объясняется быстрым истощением запаса медиаторов, снижением чувствительности постсинаптической мембраны к медиатору вследствие накопления метаболитов.
6. пластичность нервных центров. Это способность менять свои функциональные свойства.
7. доминанта. Это стойкий господствующий очаг возбуждения в ЦНС, подчиняющий себе другие нервные центры. Явление доминанты открыл А. А.Ухтомский (1923). Доминантный очаг обладает следующими свойствами: а) повышенная возбудимость; б) стойкость возбуждения; в) суммация возбуждения; г) инерция – способность длительно удерживать возбуждение после прекращения действия раздражителя. Доминанта является важным механизмом координации рефлекторной деятельности, выработки условных рефлексов и формирования целенаправленной деятельности. Например, пищедобывательное поведение всегда определяется доминантой голода.
Интегративная и координационная деятельность ЦНС осуществляется при обязательном участии торможения.
Торможение в ЦНС – это активный процесс подавления или ослабления процесса возбуждения. Оно имеет мембранно-ионную теорию происхождения. Торможение в ЦНС открыл И. М.Сеченов (1863). Впоследствии были открыты специальные тормозные структуры в спинном мозгу (Реншоу) и в продолговатом мозгу (Х. Мэгун, 1944). В зависимости от способа тормозного процесса, различают следующие виды торможения:
I) первичное, связано с наличием первичных тормозных структур (вставочных тормозных клеток и тормозных медиаторов – ГАМК, глицин).
а. постсинаптическое. В основе постсинаптического торможения лежит стойкая гиперполяризация постсинаптической мембраны вследствие увеличения её проницаемости для ионов Cl - или K+. Гиперполяризация проявляется как ТПСП, а он уменьшает возбуждение. Бывает двух видов:
- эфферентное коллатеральное (возвратное). Это торможение Реншоу, при котором тормозные вставочные нейроны возбуждаются от коллатералей эфферентных нейронов и действуют на те же нервные клетки, которые их активируют (рис. 18). Возвратное торможение – пример отрицательной обратной связи.
- афферентное коллатеральное (реципрокное). Оно вызывает торможение центра мышцы-антагониста. Например, при раздражении кожных рецепторов возникает защитный сгибательный рефлекс: центр сгибания возбуждён, а центр разгибания заторможен. В этом случае возбуждающие импульсы поступают к центру мышцы-сгибателя, а через тормозную клетку к центру мышцы антагониста – разгибателю (рис.15).
б. пресинптическое. Это торможение пресинаптической мембраны возбуждающих синапсов, осуществляется в аксо-аксональных синапсах и проявляется в виде подавления процесса деполяризации пресинаптической мембраны и освобождения возбуждающих медиаторов в синаптическую щель. Стойкая деполяризация мембраны тормозит проведение импульса к эффекторному нейрону (рис.19).
II) вторичное (пессимальное).
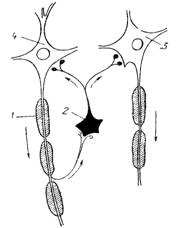
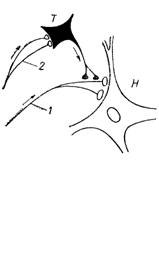
Рис.18. Схема связи между мотонейронами и клетками Реншоу: 1 – аксон с коллатералями, контактирующими с телом клетки Реншоу (2), по аксонам которой передаются тормозные влияния на мотонейроны (3,4). | Рис.19. Расположение тормозных синапсов на пресинаптических разветвлениях аксонов: 1 – волокно, проводящее афферентные импульсы, возбуждающие нейрон Н; 2 – афферентные волокна, возбуждающие тормозной нейрон Т. |
ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ.
Координация рефлекторной деятельности – это совокупность нервных процессов, обеспечивающих согласованную деятельность ЦНС и организма в целом. Взаимодействие возбуждения и торможения – основа координации рефлекторной деятельности. Основными принципами координации рефлекторной деятельности являются:

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
 Основные порталы (построено редакторами)
Основные порталы (построено редакторами)