Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Мышечные ткани
МЫШЕЧНЫЕ ТКАНИ
Мышечные ткани представляют собой тип тканей, объединенных общей специализацией – способностью отвечать на внешние и внутренние стимулы сокращением. Благодаря выраженной сократительной способности они осуществляют двигательные функции организма. Сократимость – это общее свойство клеток животных. Двигательные процессы основаны на взаимодействии сократимых белков цитоскелета – актина и миозина, но только в мышечных тканях на основе этих белков образуются специальные органеллы – миофиламенты и миофибриллы. Таким образом, для клеток мышечных тканей характерны:
· высокоупорядоченное строение сократительного аппарата (миофиламенты, миофибриллы, саркоплазматическая сеть, Т-трубочки);
· развитие специализированных цитоплазматических структур, обеспечивающих интенсивную энергетическую поддержку сократительного аппарата (многочисленные митохондрии, включения гликогена и липидов, наличие белка миоглобина, связывающего кислород)
Выделяют три основных типа мышечных тканей:
· скелетная мышечная ткань,
· сердечная мышечная ткань
· гладкая мышечная ткань.
В зависимости от организации миофибрилл - наличия или отсутствия их исчерченности, - мышечные ткани делятся на две группы:
· исчерченные (поперечнополосатые) и
· неисчерченные мышечные ткани.
Скелетная и сердечная мышечные ткани относятся к исчерченным мышечным тканям, гладкая, мионейральная и миоэпителиальные мышечные ткани являются неисчерченными.
Скелетная мышечная ткань и сердечная мышечная ткань образуют группу поперечно-полосатых мышечных тканей. В поперечнополосатых мышечных тканях миофиламенты упорядоченно организованны и образуют специальные органеллы миофибриллы. В саркоплазме клеток или мышечных волокон этой группы под микроскопом видна поперечная исчерченность.
Они имеют разное происхождение, различные тканевые элементы, способность к регенерации. Кроме того, выделяют мионейральную ткань, которая обеспечивает изменение размеров зрачка в радужной оболочке глаза (развивается из нейроэктодермы) и миоэпителиальную ткань, состоящую из миоэпителиальных (корзинчатых) клеток, которые способствуют выведению секрета из желез (слюнных, молочных) и развиваются из кожной эктодермы.
Источник развития скелетной мышечной ткани являются миотомы сомитов. Сердечная мышечная ткань имеет источником развития миоэпикардиальную пластинку, часть висцерального листка спланхнотома. Гладкая мышечная ткань развивается из мезенхимы, в основном спланхнотомной.
Тканевыми элементами скелетной мышечной ткани являются симпласты и клетки миосателлитоциты. Остальные мышечные ткани построены исключительно из клеток: в сердечной мышечной ткани они называются кардиомиоцитами, в гладкой – гладкими миоцитами.
Мышечные ткани выполняют свои функции при тесном взаимодействии с нервной тканью. При этом скелетная мышечная ткань получает соматическую двигательную иннервацию и физиологически является произвольной (её сокращение контролируется сознанием). Остальные виды мышечной ткани иннервируются вегетативной нервной системой и относятся к непроизвольным. У сердечной мышечной ткани сокращения автоматические.
Способность к регенерации на клеточном уровне у мышечных тканей различные. Скелетная мышечная ткань содержит камбиальные клетки – миосателлитоциты, и при необходимых условиях хорошо регенерирует. Гладкая мышечная ткань также способна к восстановлению, поскольку имеет стволовые камбиальные клетки. В сердечной мышечной ткани стволовые клетки отсутствуют, поэтому у взрослого человека подавляющая часть кардиомиоцитов не делится и замещается соединительной тканью. Все виды мышечной ткани способны к регенерации на внутриклеточном уровне – за счёт гипертрофии миофибрилл и других органелл.
Общая сравнительная характеристика основных видов мышечных тканей приведена в таблице 1.
Таблица 1
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА МЫШЕЧНЫХ ТКАНЕЙ
Тип мышечной ткани | |||
Поперечно-полосатая скелетная | Поперечно-полосатая сердечная | Гладкая | |
Источник развития | Миотомы мезодермы | Миоэпикардиальная пластинка (висцеральный листок спланхнотома) | Мезенхима |
Структурно-функциональная единица | Мышечное волокно - миосимпласт | Клетка - кардиомиоцит | Клетка - гладкий миоцит |
Расположение ядер | многочисленные, по периферии | В центре (одно или два) | В центре (одно) |
Исчерченность | Актин и миозин формируют характерные полоски | Актин и миозин формируют характерные полоски | Характерных полосок нет |
Сократительный аппарат | Миофибриллы | Миофибриллы | Миофиламенты |
Т-система | Т-трубочки на уровне A-I дисков; триады | Т-трубочки на уровне Z-линии, диады | Нет Т-трубочек, нет триад и диад, есть кавеолы |
Z-линия | есть | есть | Нет, имеются плотные тельца |
Кальций-связывающий белок | тропонин | тропонин | кальмодулин |
Межклеточные соединения | нет | Вставочные диски – комплекс межклеточных контактов | Щелевые контакты (нексусы) |
Источник регенерации | Деление миосателлитоцитов, гипертрофия | внутриклеточная гипертрофия | Деление миоцитов, гипертрофия |
Источник иннервации | Соматический отдел нервной системы | Вегетативный отдел нервной системы | Вегетативный отдел нервной системы |
Характер сокращения | Тетанический произвольный | Ритмический непроизвольный | Тонический непроизвольный |
Несократительные функции | Участие в терморегуляции и углеводном обмене | Синтез гормона – натрийуретического пептида (секреторные кардиомиоциты правого предсердия) | Синтез эластических волокон, коллагена III типа |
СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ
РАЗВИТИЕ
Источником развития скелетной мышечной ткани являются миотомы сомитов. Стадии развития:
- Миобластическая стадия. Клетки миотомов превращаются в миобласты и мигрируют к местам закладки мышц. Миобласты делятся митозом. Часть миобластов обособляется в виде миосателлитоцитов, которые сохраняют свойства малодифференцированных клеток до конца жизни. Миосимпластическая стадия. Миобласты располагаются в виде цепочек и сливаются друг с другом. Образуются миосимпласты. В их цитоплазме образуются миофибриллы из сократительных белков, которые начинают синтезироваться ещё в миобластах. Миофибриллы лежат на периферии миосимласта, ядра занимают центральное положение. Стадия миотубул. В симпластах увеличивается число миофибрилл. Длина их увеличивается. Стадия зрелого мышечного волокна. В эту стадию объём миофибрилл увеличивается до такой степени, что они занимают основную массу волокна, смещаясь в центр и сдвигая ядра на периферию.
СТРОЕНИЕ МЫШЕЧНОГО ВОЛОКНА.
Мышечные волокна являются структурно-функциональным элементом скелетной мышечной ткани. Они имеют длину до 20-30 см, толщину около 100 мкм. Состоят из двух частей:
· симпласта;
· миосателлитоцитов.
Мышечное волокно снаружи покрыто сарколеммой. Сарколемма состоит из толстой базальной мембраны и плазмолеммы мышечного волокна. Между базальной мембраной и плазмолеммой в отдельных участках имеются углубления, в которых расположены миосателлитоциты. Миосателлитоциты – камбиальные клетки скелетной мышечной ткани. Миосателлитоцит – типичная одноядерная клетка, окруженная своей плазмолеммой, имеет слабо развитые органеллы.
Миосимпласт содержит множество (до несколько тысяч) ядер, лежащих на периферии волокна, под сарколеммой.
Протоплазму волокна называют саркоплазмой. В ней находятся органеллы общего значения (за исключением центриолей), органеллы специального значения – миофибриллы, и включения.
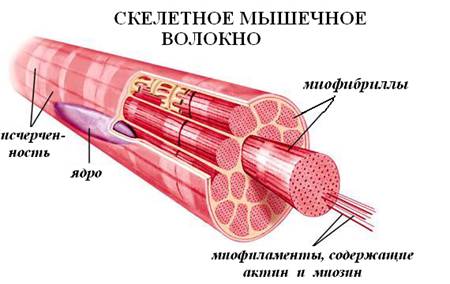

СТРОЕНИЕ МИОФИБРИЛЛ.
Миофибриллы, числом до двух тысяч в одном волокне, занимают основную часть волокна. Их длина равна длине волокна, диаметр до 2 мкм. В каждой миофибрилле при световой микроскопии обнаруживается исчерченность – чередование светлых и тёмных дисков. В поляризованном свете тёмные диски имеют двойное лучепреломление и поэтому тёмные диски называются анизотропными, или А-дисками. Светлые диски не имеют двойного лучепреломления и называются изотропными, или I-дисками.

Посередине I-диска проходит тёмная полоска, которая называется Z-линией, или телофрагмой. На поперечном разрезе телофрагма представляет собой решётку, в узлах которой закрепляются актиновые филаменты.
В центре А-диска находится более светлая полоска Н, а посередине её проходит тёмная линия М, или мезофрагма.
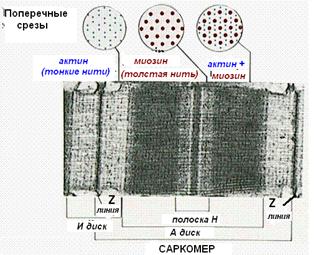
Участок миофибриллы, лежащий между соседними Z-линиями, называется саркомером. Саркомер – структурно-функциональная единица миофибриллы. В состав саркомера последовательно входят:
· Z-линия;
· ½ диска I;
· диск А;
· ½ диска I
· вторая Z-линия.
Каждый саркомер состоит из тонких актиновых и толстых миозиновых филаментов.
В составе тонких (диаметр 5 нм) актиновых филаментов входят белки:
· актин;
· тропонин;
· тропомиозин.
Молекулы актина имеют глобулярное строение – G-актин. Эти молекулы соединяются вместе в длинные цепочки – фибриллярный, F-актин. В актиновых филаментах две цепи F-актина образуют двойную спираль. В бороздках между цепями спирали лежат молекулы тропомиозина. К молекулам тропомиозина на равных расстояниях друг от друга прикрепляются молекулы тропонина. Молекула тропонина состоит из трёх субъединиц: TnT, TnI, TnC. TnT осуществляет прикрепление тропонина к тропомиозину. TnC отвечает за связывание с ионами кальция. TnI препятствует взаимодействию миозина с актином.


Толстые филаменты (диаметр 12 нм) содержат белок миозин. Каждая молекула миозина состоит из двух частей: головки и хвоста и может сгибаться в двух местах – шарнирных участках. Головка миозина имеют АТФ-азную активность и способна расщеплять АТФ с образованием энергии. Молекулы миозина соединяются в пучки и образуют толстые миозиновые филаменты. По периферии толстых филаментов находятся участки, содержащие головки миозина. Центральная часть не содержит головок.
В составе саркомера толстые филаменты лежат только в диске А. Тонкие филаменты расположены в диске I, но концами частично заходят в диск А между миозиновыми филаментами. Та часть диска А, которая содержит и актиновые и миозиновые филаменты, выглядит на срезах более тёмной, а та его часть, которая содержит только миозиновые филаменты, светлее. Эта часть диска А, которая содержит только миозиновые филаменты, и составляет полоску Н. Таким образом:
· диск I состоит из актиновых филаментов;
· полоска Н диска А состоит из миозиновых филаментов;
· на периферии диска А есть зона пересечения актиновых и миозиновых филаментов.
На поперечном срезе миофибриллы можно видеть, что в зоне пересечения вокруг одной толстой филаменты лежат шесть тонких филамент.
Тонкие филаменты неподвижно прикреплены к Z-линиям. В состав Z-линий входят белки α-актинин, десмин, виментин.
Линия М в центре Н-полоски – место соединения всех миозиновых филаментов друг с другом. В их скреплении участвуют белки миомезин и С-белок.
ДРУГИЕ ОРГАНЕЛЛЫ СКЕЛЕТНОГО МЫШЕЧНОГО ВОЛОКНА
Среди органелл общего значения в скелетном мышечном волокне хорошо развитая агранулярная (гладкая) эндоплазматическая сеть, которая называется саркоплазматическим ретикулумом (СПР). Саркоплазматический ретикулум при помощи ферментов накапливает ионы кальция. В некоторых участках плазмолемма мышечного волокна образует глубокие впячивания в виде трубочек, которые проходят перпендикулярно волокну через всю его толщину. Они называются Т-трубочками. Т-трубочки окружают каждую миофибриллу. К Т-трубочкам с обеих сторон подходят цистерны СПР – Т-цистерны. Вместе с Т-трубочками Т-цистерны образуют триады – особую мембранную систему. Триады играют важную роль в инициации мышечного сокращения.
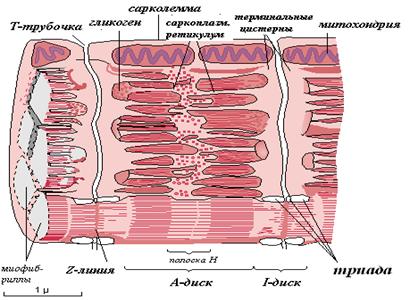

Мышечные волокна содержат также крупные митохондрии с многочисленными кристами.
Включения, характерные для скелетной мышечной ткани, - гликоген, липидные капли, которые используются для получения энергии, а также пигмент миоглобин. Миоглобин является железосодержащим пигментом, аналогичным гемоглобину, и способен связывать кислород.
МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ
Общепринятой теорией является модель мышечного сокращения, предложенная Х. Хаксли (в 1954 г.). Это теория скольжения нитей. Суть этой теории в следующем:
· нервный импульс проходит по нервному волокну и передаётся на плазмолемму мышечного волокна;
· электрический импульс идёт по Т-трубочке вглубь мышечного волокна и передаётся на лежащие рядом Т-цистерны СПР;
· мембраны СПР после деполяризации становятся проницаемыми для ионов;
· ионы кальция выходят из Т-цистерны, связываются с молекулами ТnC;
· при связывании ионов кальция изменяется конфигурация тропонина и на актиновых филаментах открываются активные центры для связывания головок миозина (которые в состоянии расслабления были закрыты TnI);
· головки миозина начинают взаимодействуют с молекулами актина: они изгибаются в шарнирных областях и создают тянущиеся усилия;
· это вызывает скольжение
Для возвращения головки миозина в исходное положение необходима энергия АТФ, которая распадается благодаря АТФ-азной активности миозина.
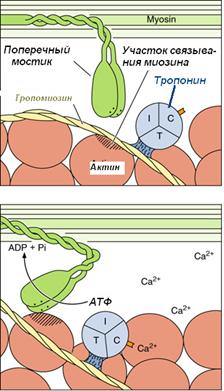 Отдельное мышечное сокращение является результатом сотен циклов образования и разъединения мостиков. Активное сокращение, которое приводит к полному взаимному перекрыванию между тонкими и толстыми филаментами, продолжается до тех пор, пока не будут удалены ионы Са.
Отдельное мышечное сокращение является результатом сотен циклов образования и разъединения мостиков. Активное сокращение, которое приводит к полному взаимному перекрыванию между тонкими и толстыми филаментами, продолжается до тех пор, пока не будут удалены ионы Са.
При отсутствии нервных импульсов ионы кальция вновь откачиваются в СПР, и тропонин-тропомиозиновый комплекс вновь закрывает участки связывания миозина на актине.
Длина филаментов при сокращении не изменяется, но поскольку актиновые филаменты заходят глубже между миозиновыми филаментами:
· длина I-дисков уменьшается;
· длина Н-полоски уменьшается, хотя длина А-диска остается неизменной;
· Z-линии сближаются и уменьшается протяженность саркомера в целом.
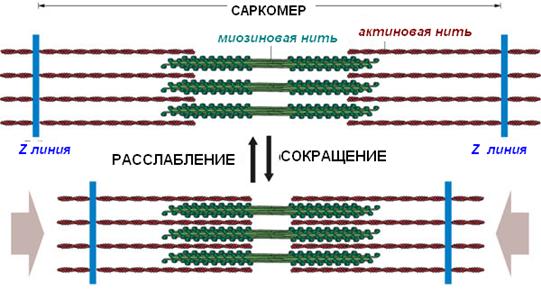

Конформационные изменения в сокращенной скелетной мышце
в сравнении с покоящейся мышцей
Сокращенная мышца | |
Диск А | Без изменений |
Диск I | Укорачивается |
Полоска Н | Укорачивается |
Z-линия | Приближаются друг к другу |
ВОЗМОЖНОСТИ РЕГЕНЕРАЦИИ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
Скелетная мышечная ткани способна регенерировать как на внутриклеточном уровне (образование в миофибриллах новых саркомеров, новых органелл), так и на клеточном уровнях. Клеточная регенерация происходит благодаря активации миосателлитоцитов вблизи зоны травмы мышечных волокон. Миосателлитоциты образуют миобласты, которые быстро делятся, сливаются друг с другом и формируют мышечные трубочки. Мышечные трубочки постепенно превращаются в полноценные мышечные волокна.
СТРОЕНИЕ СКЕЛЕТНОЙ МЫШЦЫ КАК ОРГАНА
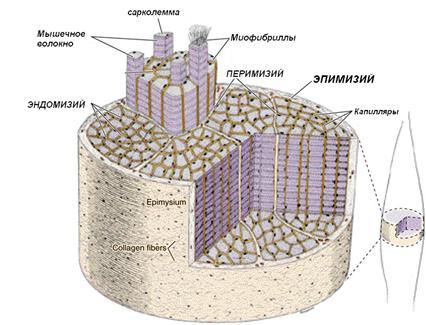

Мышца состоит из множества мышечных волокон, связанных в единое целое соединительной тканью. Между мышечными волокнами лежат тонкие прослойки рыхлой волокнистой соединительной ткани, которая называется эндомизием. Несколько мышечных волокон (от 10 до 100) окружены более толстыми прослойками рыхлой волокнистой соединительной ткани – перимизием. В эндомизии и перимизии проходят кровеносные сосуды, питающие мышцу и нервные волокна. Снаружи мышца покрыта оболочкой из плотной волокнистой соединительной ткани - эпимизием.
ТИПЫ МЫШЕЧНЫХ ВОЛОКОН
Выделяют три основных типа мышечных волокон:
- I тип – красные мышечные волокна. Это медленные тонические мышечные волокна: они могут сокращаться в течение длительного времени, но медленно. Красные мышечные волокна имеют небольшой диаметр, содержат относительно тонкие миофибриллы, много митохондрий, имеют высокую активность окислительно-восстановительных ферментов и много липидных включений. Красный цвет волокна связан с тем, что в саркоплазме этих волокон очень много белка миоглобина. Содержат много миосателлитоцитов и кровеносных сосудов. II В тип – белые мышечные волокна. Это быстрые тетанические мышечные волокна: они способны вызывать сокращения большой силы, но быстро утомляются. Характеризуются большим диаметром, сильным развитием миофибрилл, меньшим количеством митохондрий и запасы питательных веществ в виде гликогена. Миоглобина в саркоплазме мало. В волокнах низкая активность окислительных ферментов и, напротив, высокая активность гликолитических ферментов. Кровоснабжение относительно слабое. II А тип – промежуточный тип мышечных волокон. Занимают и в структурном, и в функциональном отношении среднее положение между красными и белыми мышечными волокнами.
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ
РАЗВИТИЕ
Источником развития сердечной мышечной ткани является миоэпикардиальная пластинка – часть висцерального листка спланхнотома в шейном отделе зародыша. Её клетки превращаются в миобласты и дифференцируются в кардиомиоциты. В процессе дифференцировки миофиламенты приобретают упорядоченное строение и поперечную исчерченность. Таким образом, сердечная мышечная ткань так же как и скелетная мышечная ткань относится к поперечнополосатым мышечным тканям, но в отличие от скелетной имеет клеточное строение: сердечное волокно состоит из клеток кардиомиоцитов. Развивающиеся кардиомиоциты вытраиваются в виде цепочек, соединяются друг с другом при помощи вставочных дисков и образуют функциональные мышечные волокна. В результате терминальной дифференцировки кардиомиоциты к моменту рождения или в первые месяцы постнатального развития теряют способность к делению. В зрелой сердечной мышечной ткани камбиальные клетки отсутствуют.
ТИПЫ КАРДИОМИОЦИТОВ
В результате дифференцировки кардиомиоциты превращаются в клетки трёх типов:
- рабочие, или типичные, сократительные; проводящие; секреторные (эндокринные).
СТРОЕНИЕ
Сердечная мышечная ткань образована клетками кардиомиоцитами. Сердечное мышечное волокно состоит из цепочки кардиомиоцитов, соединенных конец в конец вставочными дисками. Кардиомиоциты одного волокна имеют ответвления и их боковые поверхности образуют анастомозы с соседними волокнами. В результате межклеточных контактов миокард представляет собой сложную трёхмерную сеть, которая ведёт себя как функциональный синцитий.
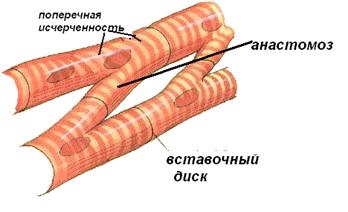
Сократительный кардиомиоцит имеет вытянутую цилиндрическую слабоотростчатую форму. Крупное светлое ядро кардиомиоцита находится в центре клетки. Многие клетки имеют два ядра и являются полиплоидными.
Цитоплазма (саркоплазма) кардиомиоцита окрашивается оксифильно. Периферическую часть саркоплазмы занимают расположенные продольно исчерченные миофибриллы, построенные так же, как и в скелетной мышечной ткани. Саркоплазматическая сеть (СПР) и Т-трубочки развиты слабее, чем в скелетной мышечной ткани, что связано с автоматией сердечной мышцы и меньшим влиянием нервной системы. В отличие от скелетной мышечной ткани:
- Т-трубочки расположены на уровне линии Z, (а не на границе А и I дисков); Т-трубочки более глубокие, покрыты базальной мембраной; Т-трубочки образуют не триады, а диады (к Т-трубочке прилежит одна латеральная цистерна СПР); типичные терминальные цистерны отсутствуют.
Между миофибриллами цепочками лежат очень крупные митохондрии с высокой плотностью крист. Митохондрии занимают до 40% объёма цитоплазмы, что связано с огромной энергетической нагрузкой. В цитоплазме кардиомиоцитов содержится большое количество включений, особенно липидных капель.
Вставочный диск – это место контактов двух кардиомиоцитов. Вставочный диск представляет собой комплекс межклеточный контактов, которые обеспечивают как механическую, так и химическую, функциональную коммуникацию кардиомиоцитов. В световом микроскопе вставочные диски имеют вид тёмных поперечных полосок, более толстых и интенсивно окрашенных в сравнении с дисками А миофибрилл. В электронном микроскопе вставочные диски имею вид зубчатой линии или ступенек. В этой зубчатой линии можно выделить горизонтальные и вертикальные участки и три зоны:
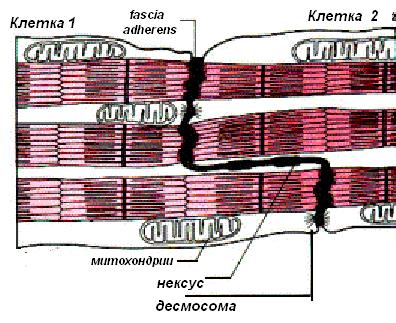

- Зоны десмосом и полосок слипания (fascia adherens). Находятся на вертикальных (поперечных) участках диска. Обеспечивают механическое соединение кардиомиоцитов, препятствуют их отделению во время циклов сокращения. Зоны прикрепления миофибрилл. Находятся на вертикальных (поперечных) участках диска. Служат местами прикрепления актиновых филаментов конечных саркомеров к плазмолемме кардиомиоцитов и аналогичны Z-линиям. Зоны нексусов (щелевых контактов) – места передачи возбуждения с одной клетки на другую, обеспечивают распространение деполяризации, вызывающей сокращение. Находятся на горизонтальных (продольных) участках диска.
Снаружи кардиомиоциты покрыты базальной мембраной, тесно связанной с межклеточным веществом, в неё вплетаются ретикулярные и эластические волокна.
Особенности строения проводящих (атипичных) кардиомиоцитов. Атипичные кардиомиоциты образуют систему генерирующие электрические импульсы (пейсмекерные кардиомиоциты синусно-предсердного узла) и проводящие эти импульсы к сократительным кардиомиоцитам миокарда (кардиомиоциты пучка Гиса, его ножек, волокон Пуркинье). Для атипичных кардиомиоцитах характерны:
- слабое развитие миофибриллярного аппарата и эндоплазматической сети,; отсутствие Т-трубочек; отсутствие типичных вставочных дисков; включения гликогена.
Несмотря на автоматизм, работа сердечной мышечной ткани регулируется вегетативной нервной системой.
Особенности строения секреторных кардиомиоцитов. Секреторные кардиомиоциты локализуются в основном в правом предсердии. В отличии от сократительных кардиомиоцитов в цитоплазме этих клеток хорошо развит секреторный аппарат: гранулярная эндоплазматическая сеть и комплекс Гольджи, и обнаруживаются многочисленные электронно-плотные секреторные гранулы. Эти гранулы содержат пептидный гормон – натрийуретический фактор (кардиодилатин). Этот гормон оказывает различные эффекты: усиливает секрецию натрия почками, расслабляет гладкие миоциты стенки артерий, подавляет секрецию гормонов, вызывающих гипертензию (альдостерона и вазопрессина). Всё это ведёт к увеличению диуреза и просвета артерий, снижению объёма циркулирующей жидкости и в результате – к снижению артериального давления.
РЕГЕНЕРАЦИЯ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ
Поскольку во взрослом организме в сердечной мышечной ткани нет камбиальных клеток, регенерация протекает на внутриклеточном уровне. При повышенной нагрузке на сердце происходит гипертрофия (увеличение размеров) и гиперплазия (увеличение количества) органелл, в том числе миофибрилл. При ранениях сердечной мышцы, инфарктах миокардах на месте погибших клеток образуется рубец из соединительной ткани.
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ
РАЗВИТИЕ
Источником развития гладкой мышечной ткани является в основном спланхнотомная мезенхима. Её клетки мигрируют и окружают эпителиальные зачатки тех органов, в состав которых входит гладкая мышечная ткань. Во взрослом организме гладкая мышечная ткань входит в состав стенки органов пищеварительного тракта, бронхиального дерева, яйцеводов, матки, мочевыделительного тракта, стенки кровеносных и лимфатических сосудов, капсулы и трабекул селезенки, стромы предстательной железы и др.
СТРОЕНИЕ
Структурно-функциональным тканевым элементом является гладкий миоцит (ГМК).
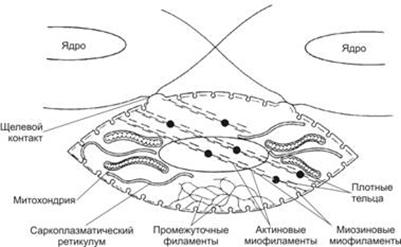
Гладкий миоцит – клетка веретеновидной формы, то есть их толщина максимальна в среднем участке, а к концам они конически сужаются. Длина гладкого миоцита может колебаться от 20 мкм (в мелких кровеносных сосудов) до 500 мкм (в матке при беременности). В каждой клетке имеется одно ядро палочковидную или эллипсоидную форму, с плотным хроматином, расположенное в центре наиболее широкой части клетки. Плазмолемма клеток покрыта тонкой базальной мембраной. В цитоплазме у полюсов ядра концентрируются элементы секреторного аппарата: эндоплазматическая сеть – место синтеза белков межклеточного вещества, комплекс Гольджи, а также мелкие митохондрии.
Для гладких миоцитов характерны многочисленные пузырьки, лежащие под цитолеммой – кавеолы. Кавеолы и пузырьки редуцированного саркоплазматического ретикулума (СПР) депонируют ионы Са, необходимые для сокращения, и являются аналогом одновременно и саркоплазматического ретикулума и Т-трубочек в исчерченной мышечной ткани. Кавеолы и СПР содержат белки транспорта кальция.
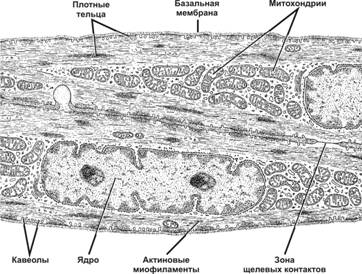

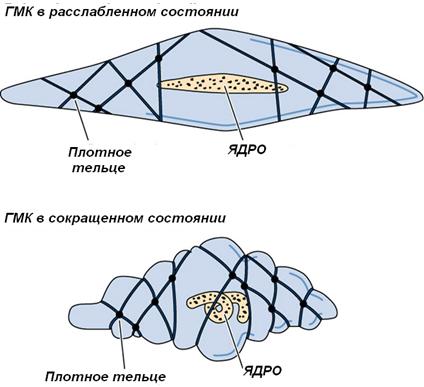
Другие структуры, характерные только для гладких миоцитов, - плотные тельца. Плотные тельца состоят из белка α-актинина и являются аналогами Z-линий саркомеров исчереченной мышечной ткани. Различают две разновидности плотных телец:
• связанные с внутренней поверхностью сарколеммы;
• свободно лежащие в цитоплазме в виде правильной цепочки.
К плотным тельцам прикрепляются актиновые и промежуточные десминовые филаменты.
В гладких миоцитах нет исчерченных миофибрилл, сократительный аппарат представлен тонкими актиновыми филаментами, которые располагаются под углом крест-накрест, образуя сеть. Актиновые филаменты закреплены в плотных тельцах.
Миозиновые нити в гладкомышечных клетках лабильны: происходит их постоянная сборка и разборка при сокращении и расслаблении соответственно.
Механизм сокращения гладких миоцитов принципиально сходен с сокращением исчерченных мышечных тканей: происходит взаимодействие актиновых и миозиновых филаментов.

• Под действием нервного импульса из пиноцитозных пузырьков высвобождается кальций, который образует комплекс с белком кальмодулином, связывающим Са;
• комплекс «Са-кальмодулин» активирует фермент киназу лёгких цепей миозина;
• фосфорилирование миозина придаёт ему способность взаимодействовать с актиновыми филаментами;
• в результате движений миозиновых филамент вдоль актиновых плотные тельца сближаются, и гладкий миоцит сокращается.
При сокращении гладкой мышечной ткани границы клеток становятся фестончатыми, а ядро – складчатым или штопорообразным. Плотные тельца передают сократительное усилие на соседние ГМК и окружающую их сеть ретикулярных волокон.

Гладкие миоциты функционируют не изолированно, а образуют миоцитарные комплексы из 10-12 ГМК. Нервные окончания подходят не ко всем миоцитам, а только к одному из комплекса. Миоциты тесно взаимодействуют друг с другом при помощи нексусов (щелевых контактов). В области нексусов базальные мембраны прерываются. Через нексусы происходит передача возбуждения, и сокращение охватывает весь комплекс.
Генетически гладкие миоциты близки к фибробластам (у них один источник развития в эмбриогенезе – мезенхима), и так же, как и фибробласты способны к синтезу межклеточного вещества. ГМК секретируют коллаген, эластин, протеогликаны.
Гладкая мышечная ткань способна к активной регенерации:
• клеточная регенерация - за счёт делений малодифференцированных клеток;
• внутриклеточная регенерация – гипертрофия, восстановление органелл.
По строению и функциям к гладкой мышечной ткани висцеральных клеток близки:
• мионейральная ткань, которая обеспечивает изменение размеров зрачка в радужной оболочке глаза (развивается из нейроэктодермы);
• миоэпителиальная ткань, состоящая из миоэпителиальных (корзинчатых) клеток, которые способствуют выведению секрета из желез (слюнных, молочных) и развиваются из кожной эктодермы.
 Основные порталы (построено редакторами)
Основные порталы (построено редакторами)