
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Курсовая работа на тему: Физиологические свойства сенсорных рецепторов
Введение …………………………………………………………………..……3-4
Основная часть
Глава 1. Понятие рецептора, классификация и морфология………..……..4-12
Глава 2. Физиологические свойства отдельных рецепторов…………..12-20
Глава 3. Значение темы для теоретика практических задач современной психологии……………………………………………………………..…………....20-22
Заключение……………………………………………………………….…...22-23
Список литературы……………………………………………………….......…24
Введение
Для развития современной теоретической психологи черезвычайно важен вопрос о том как формируются ощущения, то есть чувствование. В формировании того или иного ощущения принимает участие воспринятая тем или иным видом рецептора информация о состоянии внешней и внутренней среды. Информацию об изменяющихся условиях внешней и внутренней среды человек получает непрерывно. После чего он осуществляет переработку этой информации и составляет программы жизнедеятельности и жизнеобеспечения.
Способность приобретать сведения об изменениях окружающей среды, возможность оценивать свойства среды, ориентироваться в пространстве обеспечиваются специальными органами чувств, которые называются анализаторами. Это специальные анатомо-физиологические образования, в которых информация вводиться в мозг для переработки и анализа таковой.
Улавливателями специальных информационных сигналов в сенсорных системах служат специальные структурные образования - рецепторы. На основе их работы огромный поток информации различной модальности поступает в кору головного мозга, являющейся основой для формирования высших психических функций – мышления, сознания, памяти, эмоциональных проявлений. Поэтому изучение физиологии сенсорных рецепторов в частности физиологии сенсорных систем в целом рецепторов сенсорных, черезвычайно необходимо для более углублённого понимания психологом формирования тех или иных видов сенсорных ощущений.
Поэтому целью данной работы явилось выяснение физиологии рецепторного звена всех видов анализаторов.
В задачи работы входило
1. На основании литературных данных рассмотреть различные классификации рецепторов, особенности их структуры и морфологии.
2. На основании литературных данных рассмотреть различные физиологические свойства и особенности функционирования тех или иных рецепторов.
Глава 1. Понятие рецептора, классификация и морфология
Рецепторы выглядят как специализированные клетки, воспринимающие стимулирующее воздействие из среды организма внешней и внутренней и трансформируют его из физической или химической формы в энергию нервного импульса, так называемого потенциала действия. Другими словами можно сказать, что рецепторные клетки трансформируют энергию раздражителя действующего в паттерн нервных импульсов.
В зависимости от локализации все рецепторы делятся на экстерорецепторы ( внешние) и интерорецепторы (внутренние).
Разделяют рецепторы на определённые группы согласно природе раздражителя:
• Отделы соматической, скелетно-мышечной и вестибулярной систем, которые представляют собой периферические механорецепторы;
• рецепторы, которые воспринимают температуру внутренней среды организма, и окружающей внешней среды - терморецепторы ;
• рецепторы, которые реагируют на воздействие химических веществ - хеморецепторы;
• датчики сенсорные воспринимающие световые раздражители - фоторецепторы;
• рецепторы, возбуждающиеся механическими, химическими и температурными раздражителями, но формирующие болевые ощущения – болевые рецепторы(1).
По характеру ощущений в соответствии с психофизической классификацией рецепторов выделяют слуховые, , осязательные рецепторы зрительные, обонятельные; рецепторы боли; рецепторы положения тела в пространстве.
В зависимости от того как передаётся информация и согласно морфологии можно все рецепторы разделить на два типа : первично чувствующие и вторично чувствующие.
Рецепторы вторично чувствующие отличаются от первично чувствующих присутствием специализированных клеток - носителей первичных этапов трансформации энергии внешней в энергию нервного импульса
Из внутренних или внешних покровных тканей эти клетки происходят и являются специфическими трансмиттерами между окончанием нервным, с которым они связаны через синапсы, и внешними воздействиями по отношению к нервной системе
Свободные нервные окончания к первично чувствующим рецепторам относятся, воспринимающие смещения механические кожи, прикосновение, повреждение тканей (ноцицептивная рецепция), температурные изменения; в глубине тканей позвоночных механорецепторы. Это так называемые Тельца Пачини, Руффини, Мейсснера, диски Меркеля, колбы Краузе, генитальные тельца и другие, сюда же относятся окончания рецепторные системы сердечно-сосудистой и внутренних органов, аппарата опоры и движения.
Инициирующее звено трансформации заложено в самом нервном окончании в первично чувствующих рецепторах, в его специализированной части. Это происходит благодаря тому, что элементы-посредники почти все обладают хорошо
выраженными волосками, то есть ресничками различного размера, а у нервных окончаний зачастую такие волоски отсутствуют. Как волосково - реснитчатые так и безресничные рецепторы также могут быть. Механорецепторы таких высокоспециализированных органов чувств, как вестибулярный и слуховой аппараты относятся к вторично чувствующим, а также зрительный аппарат и ещё боковая линия рыб. из этой классификации несколько выпадают обонятельные, вкусовые и фоторецепторы.
Обонятельный аппарат восприятия и передачи информации от различных запахов представляет собой нейрон с несколько изменённой формой и его поэтому относят к образованиям первично чувствующим. Но он с другой стороны снабжён напоминающими волоски выростами, механорецепторов вторично чувствующих.
Адаптация рецептора происходит при длительном воздействии раздражителя и как следствие снижается его чувствительность. Не адаптируются рецепторы положения тела в пространстве (вестибуло рецепторы и проприоцепторы).
Человек обладает рядом периферических специализированных образований,– органов чувств. Орган чувств - это образование периферическое, перцептирующее и частично анализирующее факторы окружающей среды. Более ёмким понятием является анализатор. Анализатор – это объединение периферических и центральных образований, которые воспринимают и анализируют изменения внешней и внутренней сред организма. Не так давно возникло понятие сенсорная система. Оно стало заменять понятие анализатор дополняя его включением механизмов регуляции помощью прямых и обратных связей различных его отделов. Совокупность ощущений, обеспечиваемых одним видом анализатора или сенсорной системы обозначают термином модальность.
Органы, слуха, зрения, обоняния, вкуса, осязания относятся к органам чувств. Понятие “рецептор” и “орган чувств” не следует смешивать. Орган зрения - это глаз; а скопление фоторецепторов называется сетчатка, которая является воспринимающим световое раздражение из компонентом. Понятие “орган чувств” условно, т. к. он не может обеспечить ощущение сам по себе. Для того чтобы возникло субъективное ощущение необходимо, чтобы возбуждение, которое возникает в рецепторах, пришло в центральную нервную систему – специальные отделы коры больших полушарий, т. к. именно с функционированием отделов головного мозга связано формирование возникновения субъективных ощущений.
Рассмотрим различные органы чувств.
Орган зрения играет черезвычайно важную роль в жизни человека. Зрительный анализатор это совокупность элементов - глаза, зрительные нервы и зрительный центр, который расположен в затылочной области коры головного мозга. Около 80% информации об окружающем мире человек получает благодаря работе зрительного анализатора. Это львиная доля всей информации, которая поступает в человеческий мозг и благодаря ей человек может достаточно хорошо ориентироваться и адаптироваться в окружающем мире.
Орган слуха - характеризует возможность организма воспринимать и различать звуковые колебания. Слуховым анализатором таковая способность воплощается. Доступная область человеческому уху звуков, механических волновых колебаний составляет частоты 16-20000 Гц.
Обоняние – характеризует возможность запахи воспринимать, с помощью обонятельного анализатора. Оно осуществляется специальным рецептором, который является окончанием нервной клетки, которые располагаются в оболочке слизистой верхнего и, отчасти, среднего носовых ходов.
Вкус – специфическое ощущение, которое возникает при действии раздражителей на расположенные в различных участках языка. специфические рецепторы.
Сложное ощущение - осязание, которое появляется при рецепторном раздражении слизистых оболочек, кожи и мышечно-суставного аппарата. тактильная рецепция - прикосновение и давление принадлежит основная роль в ощущении.
Теперь рассмотрим морфологию отдельных видов рецепторов.
В первую очередь остановимся на рецепторном аппарате зрительного анализатора. Рецепторный аппарат зрительного анализатора называется сетчатка и состоит из палочек и колбочек.
Термин ко́лбочки происходит от английского cone что обозначает угол или конус представляют собой один из типов экстерорецепторов - фоторецепторы, являющимися отростками периферических светочувствительных нервных клеток сетчатки глаза. Эти клетки названы колбочками согласно форме, которая подобна лабораторной конической колбе.
Это группа рецепторов, включающая в себя различные типы нервных специализированных клеток, воспринимающих и преобразующих световые раздражения в нервное возбуждение в виде потенциалов действия биоэлектрических сигналов, которые идут в отделы зрительного анализатора головного мозга.
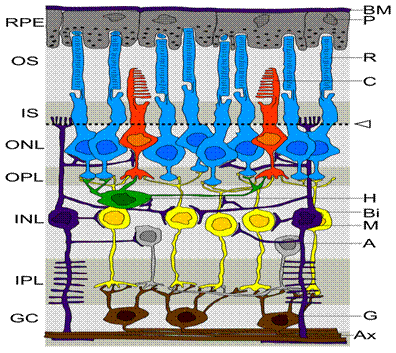
Рисунок 1. Сетчатка. Слои сетчатки RPE — пигментный эпителий сетчатки OS — наружный сегмент фоторецепторов IS — внутренний сегмент фоторецепторов ONL —внешний ядерный слой OPL — внешний сплетениевидный слой INL — внутренний ядерный слой IPL — внутренний сплетениевидный слой GC —ганглионарный слой BM — мембрана Бруха P — пигментные эпителиоциты R — палочки C — колбочки Стрелка и пунткирная линия — внешняя пограничная мембрана H — горизонтальные клетки B — биполярные клетки M — Клетки Мюллера A — амакриновые клетки G — ганглионарные клетки AX — аксоны
Колбочки воспринимают световое раздражение в широком диапазоне. Когда освещённость недостаточна для работы колбочек в сумерках, у человека работают другие рецепторные клетки-палочки. Ночью, когда люди становятся «цветнослепыми» — воспринимаемый ими мир становится монохромным(3).
Наличие в колбочках пигмента специфического — йодопсина определяет светочувствительность зрительных рецепторов; с переходом ретиналя в цис-транс форму и др. механизмами. Из нескольких зрительных пигментов состоит йодопсин. Хорошо известны и изучены на сегодняшний день такие пигменты как: хлоролаб, который чувствителен к жёлто-зелёной области спектра и эритролаб, который чувствителен к жёлто-красной части спектра.
У взрослого человека в сетчатке глаза насчитывается около 6 млн колбочек(2). Они имеют следующие размеры: около 50 мкм длина, от 1 до 4 мкм диаметром.
приблизительно в 100 раз менее чувствительны колбочки к свету, чем палочки, но они гораздо лучше воспринимают быстрые движения.
Сетчатка представляет собой слоистую структуру с четырьмя слоями нервных клеток, которые связаны синапсами. Ячейки фоторецепторов колбочек и палочек – это одиночные нейроны, которые являются непосредственно светочувствительными.
Палочки и колбочки имеют сходное строение и состоят из нескольких участков (рис2).
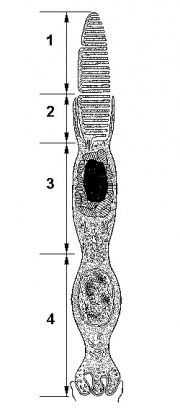
Рисунок 2. Строение рецепторной клетки сетчатки на примере колбочки
1.- Наружный сегмент, который содержит мембранные диски с йодопсином.
2.-Перетяжка - это следующий участок, так называемый связующий отдел
3.-Внутренний сегмент - это участок содержащий митохондрии.
4.- Место синаптического контакта последний участок.
Наружный сегмент колбочки заполняется мембранными полудисками, формирующимися из плазматической мембраны, которые отделяются от нее. Значит наружный сегмент это плазматической мембраны складки Мембранных полудисков меньше в колбочках, чем таковых в палочках, и их количество колеблется в районе нескольких сотен. Двумя соединёнными по краям мембранами толщиной порядка 50 - 75 Ангстрем диск каждыйобразован, которые разделены участками около 50 Ангстремт (3,4).
В районе перетяжки наружний сегмент отделен полностью от внутреннего впячиванием мембраны наружной. Цитоплазма и пара ресничек это связующее звено одного сегмента с другим с плавным переходом одного сегмента в другой. 9 пар двойных фибрилл содержат реснички, отходящих в соединительной ресничке от одной из двух центриолей, лежащих рядом перпендикулярно. Соединение ресничек, проходит от внутреннего сегмента до верхушки сегмента наружнего (5).
Есть скопление радиально ориентированных и плотно упакованных митохондрий во внутреннем сегменте. Митохондрии набухают при освещении колбочки и у них активность окислительных ферментов повышается. Здесь активный метаболизм. Митохондрии и полирибосомы, дают энергию для реализации световосприятия, при этом синтезируются белки, принимающие участие в образовании дисков мембранных и зрительного пигмента. Располагается здесь же ядро (6,7).
В области синапсов колбочек подходят и вдаются в них дендриды биполярных и горизонтальных нервных клеток сетчатки. Описаны контакты между колбочками и палочками сетчатки. В пресинаптических мембранах большое количество везикул с медиатором обнаружено. При изменении освещения количество и размер этих пузырьков меняются
Рецепторы вестибулярного анализатора состоят из двух различных типа клеток, между собой различающихся по физиологическим свойствам. Клетки двух типов имеют субмикроскопические реснички на свободной поверхности и следовательно называются волосковыми клетками. Электронной микроскопией можно увидеть стереоцилии (по 60 - 80 на каждой клетке) и киноцилии - по одной на каждой клетке. Вторичные сенсорные клетки - эторецепторы : они не имеют собственные отростки, но афферентными волокнами иннервируются нейронов вестибулярного ганглия, входящих в состав вестибулярного нерва(8,9)
Эфферентные нервные волокна оканчиваются также на рецепторных клетках. Информацию об уровне возбуждения периферического органа в центральную нервную систему передают афферентные волокна. Изменяют чувствительность рецепторов эфферентные волокна.
Различаются по строению и хеморецепторы. Первично чувствующие хеморецепторы это рецепторы элементами восприятия которых являются терминали свободные афферентных нервных волокон. Если же воспринимающей частью рецептора есть специализированная структура, не являющаяся нервным элементом, то такой рецептор называют вторично чувствующим сенсорным рецептором. Такими структурами являются сенсорные клетки у рецептора вкуса и сенсорные клетки у обонятельного рецептора. На этих эпителиальных клетках конвергируют терминали нервных волокон афферентных, транспортирующих информацию в центральные структуры сенсорных систем.
Следующим рецептором является проприоцептор, локализующийся в мышечной массе поперечнополосатой мускулатуры состоящей из экстрафузальных мышечных волокон и называется мышечное веретено. Мышечное веретено выглядит как капсула из соединительной ткани. В полости капсулы находится несколько мышечных волокон, способных к сокращению, но морфологически отличающихся от экстрафузальных волокон так называемые интрафузальные волокна. Интрафузальные волокна тоньше и короче экстрафузальных волокон.
Они имеют отличную структуру и функции от экстрафузальных мышечных волокон. Интрафузалъные мышечные волокна делятся на два типа.
Один тип- сумчато-ядерные интрафузальные мышечные волокна. средняя часть такого волокна выглядит как сумка с ядрами количество которых приближается к сотни(10,11). В другом типе волокон ядра располагаются в виде цепочки по оси интрафузального волокна. Поэтому эти волокна называют цепочно-ядерными интрафузальными волокнами. Эти волокна тоньше вдвое и почти короче вдвое, чем сумчато-ядерные волокна.
Человек имеет - около 10 млн обонятельных рецепторов. На поверхности обонятельной клетки есть так называемая обонятельная булава. По 6 - 12 тончайших волосков из неё выступает. В жидкую среду они погружены– это продукция боуменовых желёз. Присутствие волосков увеличивает в десятки раз контактную площадь рецептора с молекулами пахучих веществ. Увеличивает надежность захвата молекул пахучего вещества активная двигательная функция волосков. Булава это цитохимический центр обонятельной клетки; очевидно булава – центр генерации рецепторного потенциала.
Рецепторы вкуса — это вторичные рецепторы с микроворсинками. Вкусовые рецепторы являются частью вкусовых почек. Опорные и базальные клетки в их состав входят ещё. Отдельные рецепторы вкусовых почек являются полимодальными образованиями, так как могут воспринимать различные виды вкусовых раздражителей.
Глава 2 Физиологические свойства отдельных рецепторов.
Рецепторными клетками органа слуха являются волосковые клетки, которые состоят только из стереоцилий. На утолщении базилярной мембраны располагаются эти клетки, а их стереорцилии к текториальной мембране прикрепляются. Иннервируются чувствительными нейронами волосковые клетки (1) ганглия спирального (1). Волосковые клетки генерируют электрические реакции аналогичны таковым чувствительным нейронам в вестибулярной системе.
Механизм улавливания звука выглядит в целом следующим образом. Наружное ухо подобно акустической антенной служит направленной, улавливающей звуковые колебания системой, а проход слуховой реализует функцию волновода, который проводит их к перепонке барабанной, отделяющей ухо наружное от среднего.
Вибрации барабанной перепонки через систему слуховых косточек передаются перилимфе вестибулярной лестницы. При этом по двум механизмам усиление звукового сигнала происходит. Прежде всего, площадь перепонки барабанной значительно превышает площадь отверстия овального, закрытого стремечком. И следующее, усиливается сигнал благодаря неравенству плеч рычагов в системе слуховых косточек.
Колебания давления распространяются по вестибулярной перилимфе, а затем лестницы барабанной. Жидкость несжимаема во внутреннем ухе, поэтому окно округлое выполняет выравнивания давления функцию в улитке – круглого окна мембрана выгибается в направлении, противоположном движению стремечка. В свою очередь перилимфы колебания, порождают базилярной мембраны колебания. Благодаря этим колебаниям базилярная и текториальная мембраны смещаются друг относительно друга, что приводит к изгибанию стереоцилей волосковых клеток и изменению мембранного потенциала последних.
Палочки и колбочки имеют различную функцию и физиологические свойства. Двойственная природа зрения этим и обусловлена ночного и дневного. За ночное видение отвечают палочки, которые содержат родопсин (520 нм), функционирующие при слабом освещении. Колбочки отвечают за цветовое зрение Они содержат пигменты с максимумом чувствительности в синей, зеленой, красной частях спектра. Функционально активны при дневном свете, слабой чувствительностью к свету обладают, в темноте быстрой фазой адаптации обладают.
Распределены колбочки человека случайным образом и имеют примерно одинаковое число. Известно, что колбочки более чувствительные к длинноволновой и средневолновой частям спектра.
В фовеальной области распределение биполяров и ганглиозных клеток соответствует плотности колбочек. Соотношение фоторецепторов и ганглиозных нейроцитов на периферии сетчатки составляет 1000:1.
Образуют колбочки путь по принципу "один к одному", при котором колбочка контактирует с on - и off-биполяром и on - и off-ганглиозной клеткой, которые проецируются в клетках парвоцеллюлярного слоя наружного коленчатого тела(12,13,14).
Процесс фоторецепции инициируется поглощением кванта света светочувствительными молекулами родопсина. Он содержится в дистальной части мембраны фоторецепторов. Последующая цепь реакций молекулярных приводит к обесцвечиванию пигмента. Далее начинается каскад реакций, которые отвечают за процессы фототрансдукции, которая заканчивается гиперполяризацией мембраны клетки наружного сегмента фоторецептора.
Зрительный пигмент по своей структуре хромогликопротеид, молекула последнего хромофорную группу содержит одну, две цепочки олиго-сахаридные и водонерастворимый мембранный белок опсин.
В механизме зрения молекула зрительного пигмента выполняет две основные физиологические функции:
Во-первых, определяет диапазон фоторецепторной клетки при поглощение света в характерной области длин волн; во-вторых, процесс фоторецепторный запускает. Молекулы зрительного пигмента имеют спектр поглощения лежит в основе первой функции. Вторая функция основывается на способности молекулы пигмента зрительного при поглощении света менять свою конформацию :
В первую очередь группы хромофорной, а потом белка; молекула зрительного пигмента способность приобретает взаимодействовать с другими белками, на одной из стадий фотопревращения, которые участвуют в механизме фоторецепции. Протеиновая часть обесцвеченной молекулы зрительного пигмента активирует большое количество молекул трансдуцинового фермента, который молекулы циклического гуанозинмонофосфата инактивирует (цГМФ), который участвует в ионном обмене мембраны фоторецепторов, что к фотопревращению приводит зрительного пигмента (фотолиз) в свою очередь). на последних стадиях отщепляется ретиналь от опсина и в пигментный эпителий переносится. Хранится там он в виде эфиров кислот пальмитиновой или стеариновой, а потом в наружный сегмент фоторецепторов возвращается в виде 11-цис-ретиналя. При фотолизе происходит реакция фотохимическая — хромофорной группы изомеризация из 11-цис-формы ретиналя в транс-форму пикосекундная. Процессы последующие все в молекуле темновые и зависят от реакции температуры (15). Фотолиза процесс родопсина происходит следующим образом.
Палочковый путь включает в себя фоторецепторы палочковые, биполярные и ганглиозные клетки, а также несколько видов клеток амакриновых, которые являются интернейронами. передают зрительную информацию фоторецепторы к биполярным клеткам. Эти клетки нейронами второго порядка являются. Палочки контактируют при этом только с биполярными клетками, которые под действием света деполяризуются. Инвертирующий синапс имеется между палочками и деполяризующимися биполярами, осуществляющий передачу сигнала от гиперполяризующихся под действием света палочек с мембранным потенциалом негативным к палочковым деполяризующим с позитивным мембранным потенциалом биполярам. Эти биполяры деполяризующиеся назваются клетками с оn-центром. Такие пути палочковые называют on-путями. Система палочковая конвергентна и дивергентна, определяющая обработку и усиление сигнала для транспортировки его в мозг.
Путь колбочек отличается от палочкового тем, что уже в наружном слое имеют колбочки связи обширные и связывающие синапсы их с биполярами колбочковыми типов различных. Деполяризуются одни из них так же как палочковые деполяризующиеся биполяры и формируют on-путь колбочковый с синапсами инвертирующими, другие гиперполяризуются, формируя off-путь, в котором контактируют колбочки с биполярами посредством синапсов предохраняющих. В фовеальной области колбочки, каждая из которых связана с парой биполяров с on - и off-центром и ганглиозными клетками с on - и off-центром, формируют соответствующие on - и off-каналы.
Прямые синапсы имеют биполяры колбочек с дендритами клеток ганглиозных без амакриновых промежуточных клеток. Путь колбочковый состоит из двух каналов параллельных, которые идут от колбочек к нейроцитам биполярным и ганглиозным. Связан с сигналами один канал, который возникает на стимул, где он ярче фона (on-center channel), другой — на стимул, более темный, чем фон (off-center channel). Эти пути-каналы восприятие обеспечивают зрительных образов контраст.
Основные четыре типа ганглиозных клеток выделяют. В общеизвестных работах выделяют. Клетки 1-го типа имеющие круглые поля рецептивные, которые состоят из центральной и периферической зон с разной чувствительностью спектральной и связями тормозными в направлении от периферии к центру. Без двойной оппонентности эти поля называют концентрическими(16).
Второго типа клетки формируют круглые рецептивные поля, с отсутствующей периферической зоной. Они избирательно реагируют на излучение волн с различными длинами, и имеют противоположные связи внутри единственной своей зоны и отвечают реакциями с противоположным знаком при стимуляции различными цветами.
Встречаются среди нейронов к цвету не чувствительные, но свойствами пространственной организации полей обладающие, которые сходны со свойствами других нейронов. черно-белыми нейрональными клетками или on - и off-нейронами их называют.
Третьего типа клетки — это нервные клетки, которые не чувствительны к длине волны излучения, которые имеют организацию концентрическую рецептивных полей со связями тормозными от периферической к центральной зоне. Клетки четвёртого типа – это нейроны по типажу, по строению близкие к нейронам первого типа, но отличаются от них более обширной спектральной чувствительностью. Их поля рецептивные включают центральную и периферическую зоны, каждая из которых реагирует избирательно на цветовой стимул с различными длинами волн. Среди клеток 1-го типа выделяют два класса: и желто-синие (Y-B) и красно-зеленые (R-G). В 8 раз больше клеток красно-зеленых, чем сине-желтых. Таким образом, существуют в сетчатке три параллельных канала, которые связанны с цвето восприятием. Один из них передает различие сигнала между средне - и длинноволновыми колбочками, а второй — разницу сигнала коротковолновых колбочек из суммы сигналов средне - и длинноволновых колбочек, а третий — суммарный колбочковый ответ всех трех классов. В человеческой сетчатке переработка сигналов колбочек имеет оппонентный характер.
Появление нейронов, которые отвечают реакциями знаками противоположными, определяющим фактором является функционирования механизмов цветового зрения и пространственного. В сетчатке человека, таким образом, от рецепторного слоя к клеткам ганглиозного слоя тянется сеть связей нейронных, которые образуют множество путей, или каналов пострецепторных, которые предназначены для передачи и обработки информации сенсорной. Они выполняют разные функции и работают как параллельные системы.
РП(рецептивное поле) нейрона — это специфическая структура, которая состоит из нейрона выходного, клеток промежуточных и объединения рецепторов сетчатки, в пределах которой изменение светового потока в пространстве или времени вызывает изменение реакции выходного нейрона.
Имеют центральную и периферическую зоны рецептивные поля, которые при стимуляции формируют противоположные по знаку реакции, например оn-реакцию — в центре и off-реакцию — на периферии. Существуют взаимотормозные отношения между центром и периферией так, что одновременное освещение обеих зон к уменьшению приводит как центральной, так и периферической реакций. Периферия противоположная центру ганглиозной клетки по механизму обратной связи организована от горизонтальных клеток к фоторецепторам.
Физиологические свойства вестибулярных рецепторов. На нейрон афферентный возбуждение передается с помощью нейротрансмиттера ацетилхолина, который выделяется из клетки рецепторной. Причем в рецепторной клетки проходят и афферентные волокна, с помощью которых регулируется функциональное состояние клетки, а следовательно, ее чувствительность.
Сгруппированы рецепторные клетки одного типа вестибулярного органа в трех ампулах полукружных каналов и макуле мешочка. Клетка каждая имеет одну длинную волос - киноцилий и 50-60 коротких - стереоцилии. Стимулом адекватным рецепторной клетки является смещение пучка ресничек в сторону киноцилии волоса. В клетке повышается вследствие этого проницаемость мембраны для Na +, что приводит к возникновению рецепторного потенциала.
В ответ на его появление в синапсе, располагающийся между афферентным волокном и клеткой, медиатор выделяется, под действием которого в мембране постсинаптической, появляется потенциал генераторный. Постсинаптическая мембрана принадлежит афферентным волокнам. Суммирование генераторных потенциалов к возникновению потенциала действия (ПД) приводит. Следует понимать, что и в условиях естественных от рецепторных клеток во время наклона ресничек в сторону волоса длинного увеличивается частота потенциалов действия, а в противоположную сторону - уменьшается. Таким образом в улавливании адекватного раздражения участвуют волоски рецепторных клеток формируя при этом генераторный потенциал, который суммируясь даёт нервный импульс транспортирующийся в центральные отделы анализатора - специальные отделы неокортекса.
Остановимся на функциях мышечного веретена.
Мышечное веретено располагается внутри мышечной массы экстрафузальных волокон параллельно. При растяжении экстрафузальных мышечных волокон растягиваются и интрафузальные волокна мышечных веретён или при возбуждении гамма мотонейронов происходит сокращение интрафузальных мышечных волокон, что приводит к тому что центральная ядерная часть рецептора начинает колебаться и приводит в возбуждение нервные окончания, которые передают информацию через нейрон псевдо униполярный в задние рога спинного мозга и там переключаясь на интернейрон в конечном итоге заканчиваются на теле альфа мотонейрона, который иннервирует экстрафузальные волокна мышцы, где расположен данный рецептор. Это так называемая гамма петля. Главной функцией мышечного веретена является информировать мозг об изменении длины рабочих мышечных волокон и таким образом предохранение экстрафузальных волокон от перерастяжения и участвует в механизмах перераспределения поддержания мышечного тонуса.
Функции сухожильных рецепторов.
Сухожильные Гольджи рецепторы локализуются в месте перехода мышечных волокон в сухожилия. Их длина составляет 0,5-1,0 мм, а диаметр – 0,1-0,2 мм. Аксон отдельный нервный несет импульсы афферентные в спинной мозг и является аксоном Ib. Он в виде завиточков начинается, проходящих между волокнами коллагеновыми сухожилия. При сокращении мышечных волокон, волокна коллагеновые натягиваются и сжимают веточки нервные, которые начинают генерировать импульсы. Таким образом, в результате закрепления последовательного сухожильных органов к мышечным волокнам они возбуждаются при уменьшении длины возбужденной мышцы. Рецепторы сухожильные возбуждаются в 1,5 – 8 раз более эффективно при мышечном сокращении, нежели при пассивном растяжении.
Следующие рецепторы, которые необходимо рассмотреть это рецепторы вкусового анализатора.
Структурами, непосредственно воспринимающими вкусовой раздражитель являются микроворсинки вкусовых клеток. Потенциал мембранный вкусовых клеток колеблется от —30 до —50 мВ. Рецепторный потенциал величиной от 15 до 40 мВ возникает при действии вкусовых раздражителей. Он выглядит как деполяризация поверхности вкусовой клетки, которая служит причиной возникновения в волокнах барабанной струны и нерва языкоглоточного генераторного потенциала, переходящего по достижении критического уровня деполяризации в распространяющиеся потенциалы действия или нервные импульсы. Возбуждение с рецепторной клетки проходит через синапс на нервное волокно с помощью нейротрансмиттера ацетилхолина. Некоторые вещества, такие как например СаС12, хинин, соли металлов тяжелых, инициируют возникновение не первичной деполяризации, а первичную гиперпрляризацию. Ее возникновение связано с осуществлением отвергаемых отрицательных реакций [14]. При этом не возникает распространяющихся импульсов.
Физиологические свойства первично чувствующих рецепторов примерно одинаковы воспринимать адекватный раздражитель. Физиологический механизм передачи нервного импульса по сути един.
ГлаваЗ. Значение темы для теоретика практических задач современной психологии.
Человеку необходимо получать информацию о постоянно меняющихся условиях окружающей среды, для того что бы адаптироваться в окружающем мире. Для этого необходимо человеку нормальное функционирование так называемых сенсорных систем, которые передают информацию о физических проявлениях окружающего мира. Эта информация воспринимается человеком в виде ощущений. На основе формирующихся ощущений человек познаёт мир. Формируются знания и познания.
Проблема познания как отражения человеком объективной действительности является фундаментальной общей проблемой как писихологии так естествознания. В этой связи к психологии предъявляются требования глубокого исследования основных ступеней и форм познавательной деятельности человека, прежде всего — чувственного или образного, непосредственного отражения в мозгу человека предметов, явлений и процессов объективной действительности.
Именно поэтому на всех этапах развития психологической науки усилия многих ученых были сосредоточены на изучении ощущений, восприятий и представлений как образов внешнего мира. В этой связи важен вопрос чувственного познания, включающего основные анализаторные деятельности человеческого мозга в их взаимодействии. Это тем более необходимо, что наука открыла и открывает ранее неизвестные и скрытые возможности чувственного познания, которые все еще недостаточно используются в практике обучения, диагностики и лечения и т. д.
Становиться понятным значимость знаний физиологической работы наших сенсорных систем для более глубокого понимания теоретиком психологом формирования чувственных ощущений и на их основе чувственного познания окружающего мира. К этой проблеме интерес также связан с ленинской теорией отражения, которая открывает общую диалектическую закономерность перехода от ощущения к мысли. Систему определенную, подчиненную этой закономерности, составляют переходы и видоизменения чувственного познания, являющегося основой логического мышления, которое неразрывно связанно с языком и речью. На основе работы наших анализаторов поддерживается работа мозга, формируются не только ощущения, но и вся когнитивная сфера. Когнитивная сфера включает в себя
Такие категории как внимание, память, гносис, праксис, речь, мышление, сознание. И наконец на основе поступившей и переработанной нашим мозгом информации формируется целенаправленное поведение непременным обеспечением которого служат эмоции. Эмоции это тоже категория, которая изучается психологией, особенно их нарушение. Формирование наших эмоциональных проявлением так же связанно с работой сенсорных систем. Так например употребили мы вкусно приготовленную пищу, хорошо поработал вкусовой анализатор а человек в конечном итоге получил положительную эмоцию – радость, удовлетворение.
Приехали мы на природу активно заработали слуховой, зрительный, обонятельный и двигательный анализаторы. В конечном итоге масса положительных эмоций масса ощущений и множество знаний об окружающем природном ландшафте(15).
С другой стороны болезнь человек испытывает боль заработала болевая рецепция опять же знания о конкретной болезни, но и положительные эмоции тоже. Исходя из всего вышесказанного следует сделать заключение, что всю нашу психическую деятельность создаёт мозг, а работу мозга в свою очередь поддерживают нормально функционирующие сенсорные системы.
Особой структурой головного мозга, формирующей нашу психику является кора головного мозга. А в коре локализованы все конечные пути наших анализаторов. Нарушение работы хотя бы одного анализатора или его полное выпадение приводит к стойким нарушениям и высших психических функций. Но если не работает один орган чувств, то обостряется работа других органов чувств. Итак, на основе всего вышесказанного можно сделать вывод, что изучение и умножение знаний о фозиологии сенсорных рецепторов в частности и физиологии сенсорных систем в целом крайне необходимы для как для теоретической так и для практической психологии(15,16).
Заключение.
Резюмируя всё вышеизложенное, что в организме человека сосредоточенно множество различных видов рецепторов, каждый из которых направлен на улавливание воздействия определённого раздражителя со стороны внешней или внутренней среды, преобразования энергии раздражителя в энергию нервного импульса и дальнейшую передачу этого импульса в корковый отдел анализатора. На основании переработки и анализа пришедшей информации формируется программа соответствующего поведения и его реализация. На основе работы сенсорных рецепторов формируются те или иные ощущения Ощущения осуществляют непосредственную связь сознания человека с окружающим миром. Субъективное отражение в образах, как продукт рефлекторной деятельности мозга, есть лишь высшая форма отражения действительности. Простейшие формы которого свойственны любой движущейся материи, в том числе и неорганической. Проблема ощущений для психолога теоретика ставится как проблема первейшего условия познания. Отражение невозможно без постоянного и непрерывного превращения энергии, воздействующей на человека материи в факты сознания человека. Это превращение осуществляется бесконечной (для индивидуальной жизни) массой ощущений.... Все эти колоссальные массы ощущений ассоциируются по самым разным признакам, синтезируются в актах восприятия, закрепляются в виде следовых явлений, особенно в форме представлений, и входят обобщенными группами в состав мыслительных актов. Всё вышеизложенное свидетельствует о том, что знания физиологии рецепции черезвычайно важны для теоретической психологии.
Литература
1. Смирнов человека 2001 г. 605 с.
2. G. Osterberg (1935). "Topography of the layer of rods and cones in the human retina, " Acta Ophthalmol., Suppl. 13:6, pp. 1-102. Wolken J. J. 1966. Vision, Thomas C. C.
3. De Robertis E. I., A. Lasansky. 1961. In^ The strukture of the Eye. New York – London : 29
4. Wald G., P. K. Brown, J. R/ Gibbons. 1963. Jur. Opt. Soc. Amer., 53 : 20
5. 1961. Жур. Общей биол., 22 : 471
6. 1962. Доклад АН СССР, 145 : 669
7. 1961. Жур. Общей биол., 22 : 474
8. 1965. Жур. Высш. Нервн. Деят., 5 : 934
9. 1966. Жур. Высш. Нервн. Деят., 16 : 535
10. Н. Грин, У. Стаут, Д. Тейлор, Биология: в 3-х т, пер. с англ./ под. ред. Р. Сопера, Мир, т.2, 280—281
11. Zaidi FH, Hull JT, Peirson SN, Wulff K, Aeschbach D, Gooley JJ, Brainard GC, Gregory-Evans K, Rizzo JF 3rd, Czeisler CA, Foster RG, Moseley MJ, Lockley SW. Short-wavelength light sensitivity of circadian, pupillary, and visual awareness in humans lacking an outer retina. Curr Biol. 2007 Dec 18;17(24):2122-8 Abstract.
12. , , 1955. Тр. Гос. оптич. инст., 24 : 33.
13. Д. Хьюбел Глаз, мозг, зрение. — под ред. . — М.: Мир, 1990. — 172 с.
14. Маклаков психология. СПб: Питер, 2001
15. 1. B. Marks, W. U. Dobelle, E. F. Mac Nichol Visual Pigments of Single Primate Cones = "Science", v 143, 1964, p 1181. — Department of Biophysics, Johns Hopkins University, Baltimore, Maryland 21218: "Science", v 143, 1964, p 1181, 1964. — Т. v 143. — С. p 1181.
