


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
РЕНТГЕНОГРАФИЯ ЦЕЛЛЮЛОЗ
(Петрозаводский госуниверситет, Петрозаводск, aleshins@psu.karelia.ru)
Рентгенографические исследования целлюлоз, выполняемые в большеугловой области углов рассеяния 2q, позволяют определить следующие характеристики надмолекулярной структуры:
· размеры областей когерентного рассеяния (ОКР), чаще называемых блоками мозаики или кристаллитами, то есть областей регулярно расположенных молекулярных фрагментов;
· степень кристалличности, величина которой характеризует долю молекулярных фрагментов, имеющих регулярную упаковку, от полного числа регулярно и хаотически расположенных молекулярных фрагментов, участвующих в рассеянии.
Размеры кристаллитов определяются из ширины брэгговских отражений в направлении нормали к отражающим плоскостям. Наличие нескольких отражений а рентгенограмме позволяет определит размеры ОКР в соответствующих кристаллографических направлениях и оценить степень их изотропии.
Совокупность молекул, имеющих регулярную упаковку, обусловливает появление на дифракционной картине брэгговских отражений, а хаотически расположенные молекулярные фрагменты приводят к появлению диффузного рассеяния, имеющего максимум в области основных отражений на рентгенограммах целлюлоз.
Три основные фазы целлюлозы: триклинная Ia, моноклинные Iβ и целлюлоза II. В триклинной фазе Ia на элементарную ячейку приходится один целлобиозный остаток, так что в пространственной модели, получаемой путем трансляции элементарной ячейки, упаковка молекул параллельная. Для моноклинной ячейки целлюлозы Iβ предложено 3 модели: одна с антипараллельной и две с параллельной up и down ориентацией молекул [см. ссылки в 1]. В целлюлозе II упаковка молекул антипараллельная [2].
В таблице 1 приведены периоды и углы элементарной ячейки для триклинной и моноклинной моделей.
Таблица 1.
Кристаллографические характеристики фаз I, Iβ и целлюлозы II.
фаза | a, Å | b, Å | c,Å | a° | β° | γ° | V, Å3 | Источник |
Ia, триклинная | 6.74 | 5.93 | 10.36 | 117. | 113. | 81.3 | 339.44 | 1 |
Iβ, моноклинная | ||||||||
взаимное расположение молекул | a, Å | b, Å | c,Å | a° | β° | γ° | V, Å3 | Источник |
антипараллельное | 7.846 | 8.171 | 10.34 | 90 | 90 | 96.38 | 658.79 | 1 |
параллельное up | 8.18 | 7.84 | 10.38 | 90 | 90 | 97.04 | 660.66 | 1 |
параллельное down | 8.18 | 7.84 | 10.38 | 90 | 90 | 97.04 | 660.66 | 1 |
целлюлоза II | ||||||||
антипараллельное | 8.01 | 9.04 | 10.36 | 90 | 90 | 117.1 | 667.81 | 2 |
На рисунке 1 представлены соответствующие рентгенограммы, рассчитанные в программе Mercury при значении ширины отражений 0.1°.
На теоретических рентгенограммах целлюлозы I, соответствующих рентгенограммам хорошо закристаллизованных поликристаллов, наблюдается всего три интенсивных линии в области углов рассеяния на излучении FeKa от 17 до 30°. Друг от друга рентгенограммы отличаются наличием или отсутствием слабых отражений и перераспределением интенсивностей. Рентгенограмма целлюлозы II резко отличается от рентгенограмм целлюлозы 1. Тем не менее, рентгенограммы обеих фаз имеют вид, характерный для полимеров с цепочечной структурой.
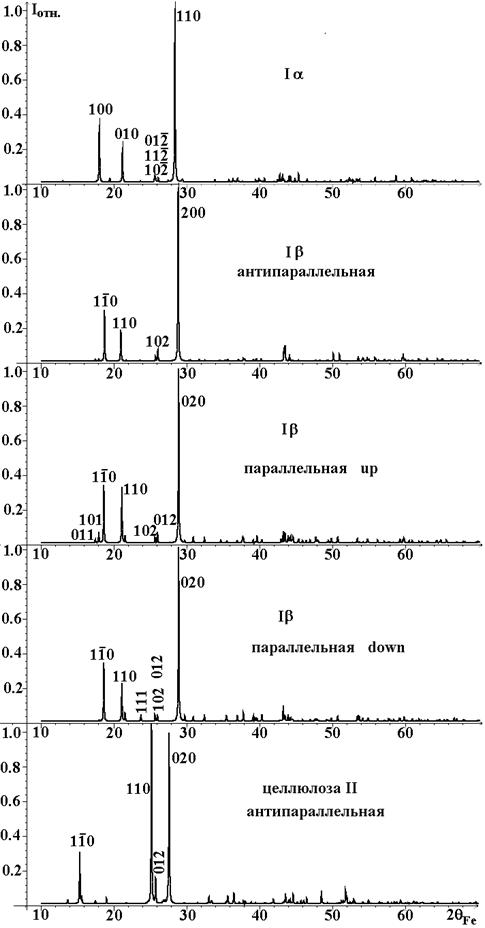
Рисунок 1. Рентгенограммы триклинной и моноклинных моделей строения целлюлозы.
Многочисленные исследования показали, что реальные объекты характеризуются аморфно-кристаллическим строением, при этом ширина отражений от кристаллической компоненты достигает нескольких градусов. В результате на рентгенограммах целлюлозы I имеет место перекрывание отражений даже в области интенсивных линий. Наличие ориентированных цепочек в кристаллической составляющей приводит к анизотропии строения и, как следствие, к анизотропии рассеяния. В результате рентгенограммы синтезированной из растительных объектов целлюлозы I и II, зарегистрированные в геометриях на отражение и прохождение, различны (рис. 2).
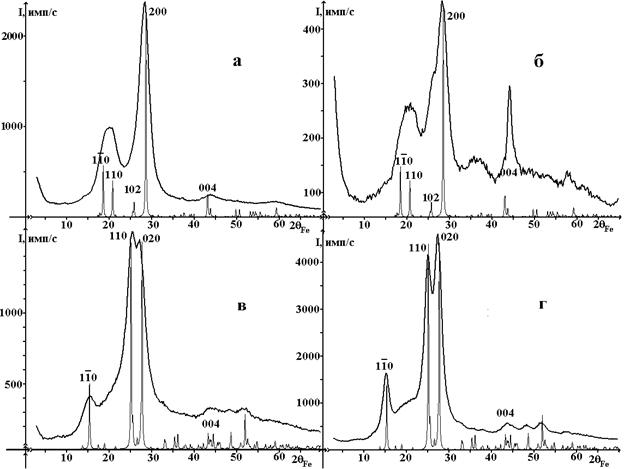
Рисунок 2. Характерные рентгенограммы исходной Ib (а, б) и мерсеризованной II(в, г) древесной целлюлозы: а, в - отражение; б, г) прохождение; в сравнении с теоретически рассчитанными рентгенограммами моноклинной фазы целлюлозы Iβ и II с антипараллельной ориентацией цепочек.
На рис. 3 указаны направления [hkl], вдоль которых из ширины соответствующих отражений, индексы которых приведены на рентгенограммах, рассчитывались размеры областей кристалличности Dhkl.
| Рис. 3. Ориентация осей элементарной ячейки целлюлозы 1β и некоторые кристаллографические направления. |
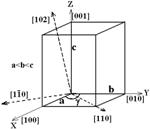
Из размеров кристаллитов в направлениях [![]() 10], [110] и [100] кристаллической решетки целлюлозы определяются линейные размеры и форма поперечного сечения элементарных фибрилл, длина области упорядочения вдоль оси элементарной фибриллы рассчитывается из размера Dhkl, определенного в направлении [001] из ширины отражения (004). Последнее появляется лишь на рентгенограммах, зарегистрированных на прохождение (рис. 1).
10], [110] и [100] кристаллической решетки целлюлозы определяются линейные размеры и форма поперечного сечения элементарных фибрилл, длина области упорядочения вдоль оси элементарной фибриллы рассчитывается из размера Dhkl, определенного в направлении [001] из ширины отражения (004). Последнее появляется лишь на рентгенограммах, зарегистрированных на прохождение (рис. 1).
Методики расчета степени кристалличности из дифракционных данных проанализированы в работе [3]. Показано, что наиболее точные результаты получены традиционными методами Сегала и Руланда, а проблемы последних связаны только со сложностью разделения отражений кристаллической компоненты. Использование программы обработки спектров позволяет модифицировать метод Руланда: разделить отражения от кристаллической фазы и корректно выделить рассеяние аморфной компонентой. Предварительно в экспериментальный спектр вводятся поправки на поглощение, поляризацию и учитывается угловая зависимость фактора интегральности [4].
Параллельно в процессе расчета определяются ширины отражений от кристаллической фазы [4], что дает возможность, используя формулу Шеррера, рассчитать размеры областей кристалличности в различных направлениях в решетке.
На рис. 4 приведена характерная диаграмма, построенная по значениям размеров ОКР, рассчитанных для различных направлений в решетке кристаллической фазы.
| Рис. 4. Форма поперечного сечения элементарных фибрилл для образца МКЦ. Цифры соответствуют длине ОКР в данном направлении, показанной от ее центра. + указана длина элементарных фибрилл, рассчитанная из рентгенограмм, отснятых в геометрии на прохождение. На направлениях, на которых отсутствует знак +, значения, рассчитанные из рентгенограмм, отснятых на просвет и отражение, совпадают и обозначены •. |
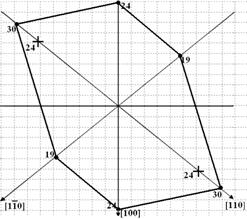
Следует отметить, что определенные из рентгенографических данных форма и размеры поперечного сечения элементарных фибрилл растительных целлюлоз согласуются с данными, полученными в [5] методом атомно силовой микроскопии.
Использование метода Ритвельда для анализа рентгенограмм целлюлоз позволяет определить фазу целлюлозы, периоды, углы и объем элементарной ячейки и проследить за их изменениями в процессе той или иной обработки, установить зависимость указанных характеристик от исходного сырья. Используя в качестве входных данных для целлюлозы I приведенные в литературе значения координат атомов можно установить соответствует ее структура фазе Ia или фазе Ib, а также характер взаимной ориентации молекул в фазе Ib.
В частности, установлено, что структура бактериальной целлюлозы соответствует триклинной модификации Ia [6]. Выполненные нами многочисленные исследования растительных целлюлоз [7] показали, что это, как правило, целлюлоза Ib различной степени совершенства и, чаще всего, с антипараллельной ориентацией молекул.
Регенерация целлюлозы в ДММА/LiCl [8], а также модификация мерсеризованной целлюлозы эпихлоргидрином в среде бензола и бромистым этилом в среде бензола приводит к образованию аморфных продуктов, о чем свидетельствует диффузная дифракционная картина (рис. 5).
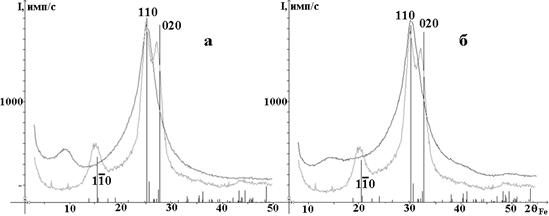
Рис. 5. Сравнение рентгенограмм мерсеризованной хвойной небелёной сульфатной целлюлозы: до модификации ▬ и в результате модификации ▬ : а) эпихлоргидрином в среде бензола,
б) эпихлоргидрином. Вертикальные линии - штрихдиаграмма целлюлозы II.
Для аморфных объектов из эксперимента рассчитываются кривые распределения парных функций, характеризующие распределение электронной плотности исследуемого материала Результаты расчета представлены в работах [8, 9].
Расчет координационных чисел и радиусов координационных сфер из кривых распределения парных функций дает только одномерную информацию о структуре, поэтому для получения атомных конфигураций в области ближнего упорядочения активно начинают привлекаться методы компьютерного моделирования [8, 9].
Размеры неоднородностей электронной плотности целлюлозных материалов и наличие или отсутствие их упорядоченного расположения исследуются методами малоугловой рентгенографии и нейтронографии [10]. Особенно активно данные исследования выполняются при изучении структуры лигнинов.
Литература
1. , , Силина представления о структуре целлюлоз //Химия растительного сырья. - 2001. - №1. - с. 5-36.
2. , Алешина метода Ритвельда к уточнению структуры порошковой мерсеризованной целлюлозы // Естественные и технические науки. - 2011. -№3. - с. 37-43.
3. Thygesen A., Oddershede J., Lilholt H, Thomsen A-B and Kenny S. On the determination of crystallinity and cellulose content in plant fibres // Cellulose - 2005.-12. - p.563–576.
4. , , Алешина выявления различий структурного состояния целлюлозы / патент RU 2570092, ФГБОУ ВО ПетрГу.
5. Baker A. A., Helbert W., Sugiyama J., Miles M. J., Wills H. H. New Insight into Cellulose Structure by Atomic Force Microscopy Showsthe I_ Crystal Phase at Near-Atomic Resolution // Biophysical Journal – 2000. - 79. - p. 1139 -1145
6. Гладышева рентгенографических исследований бактериальной целлюлозы // Фундаментальные исследования. -2015.- №7 (часть 2) – с. 240-244
7. Мелех и технические целлюлозы / Изд-во LAP Lambert Academic Publishing. -2014.- 104 с.
8. Прусский а. И., Алешина моделирование атомной структры регенерированной целлюлозы // ВМС А. – 2016.- 57.- с. 1-14.
9. Алешина л. а., Коновалова порядок в модифицированных эпихлоргидрином мерсеризованных целлюлозах / Структура и физико-химические свойства целлюлоз и нанокомпозитов на их основе. Глава5. -2014. - с.123-154.
10. Vainio U., Maximova N., Hortling B., Laine J, Stenius P., Simola L. K., Gravitis J. , Serimaa R. Morphology of Dry Lignins and Size and Shape of Dissolved Kraft Lignin Particles by X-ray Scattering // Langmuir. - 2004. - 20. - p. 9736-9744
