

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
По аналогии с ГФК было доказано, что появление второго ИП на кинетических кривых восстановления антрахиноновых красителей также обусловлено накоплением в реакционной среде в аэробных условиях восстановленных форм кислорода. Вместе с тем и в данном случае, как и в случае с ГФК, для того, чтобы второй ИП проявлялся в виде плато, необходимо наличие в молекуле окислителя двух типов фрагментов, один из которых восстанавливается легче другого. Следовательно, восстанавливающиеся хинонные группы неэквивалентны, причиной чего, как свидетельствуют литературные данные, может быть вовлечение одной из них во внутримолекулярную водородную связь (ВВС) с водородом мостиковой аминогруппы. Проведенные квантово-химические расчеты также свидетельствуют о том, что в молекулах изученных красителей склонность одной из хинонных группировок к образованию ВВС в 1.38-4.68 раз выше, чем другой.
Для доказательства правомерности проведенных рассуждений в исследование было включено еще одно соединение (b-антрахинонмоносульфокислота), в котором из-за отсутствия заместителей в a-положении ВВС невозможна. Кинетическая кривая аэробного восстановления этого соединения абсолютно монотонна (второй индукционный период отсутствует).
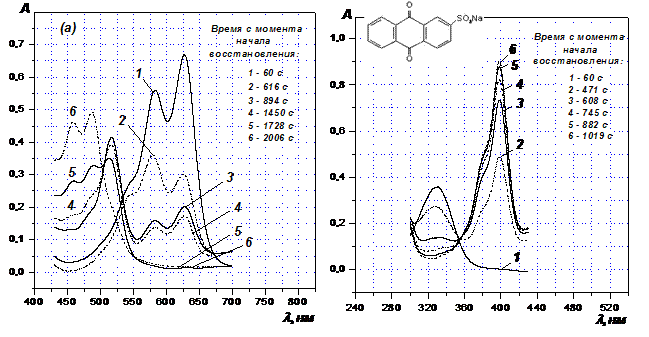
В спектрах красителей Acid Green 27и Acid Blue 80, регистрировавшихся по ходу их восстановления, зарегистрирован устойчивый промежуточный продукт (семихинон ион-радикал) (Рис. 5 (а)). В спектрах b-антрахинонмоносульфокислоты он отсутствует (Рис. 5 (б)). Следовательно, образование устойчивого семихинона как при аэробном, так и при анаэробном восстановлении 1,4-диаминопроизводных антрахинона обусловлено вовлечением одной из восстанавливающихся групп в ВВС.
|
 Проведенные исследования обобщены в виде схемы аэробного восстановления 1,4-диамино-производных антрахинона (Рис. 6). На схеме можно выделить параллельные реакции восстановления двух форм красителя, одна из которых имеет внутримолекулярную водородную связь (I), а вторая – нет (II). Первая образует устойчивый семихинон (III), вторая – неустойчивый (IV). Это и определяет кинетические особенности восстановления 1,4-ди-аминозамещенных антрахинона.
Проведенные исследования обобщены в виде схемы аэробного восстановления 1,4-диамино-производных антрахинона (Рис. 6). На схеме можно выделить параллельные реакции восстановления двух форм красителя, одна из которых имеет внутримолекулярную водородную связь (I), а вторая – нет (II). Первая образует устойчивый семихинон (III), вторая – неустойчивый (IV). Это и определяет кинетические особенности восстановления 1,4-ди-аминозамещенных антрахинона.
|
3.2.2 Гумусовые кислоты торфа как редокс катализаторы восстановления
хинонных группировок в молекулах красителей
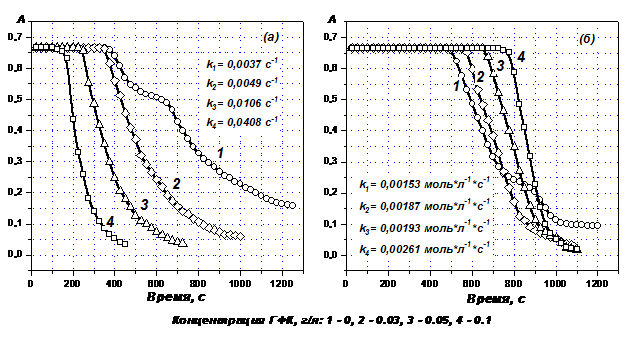 Изучено влияние ГФК торфа на кинетику восстановления красителей Acid Green 27 и Acid Blue 80 гидроксиметансульфинатом натрия и диоксидом тиомочевины в аэробных условиях в щелочной среде (Рис. 7).
Изучено влияние ГФК торфа на кинетику восстановления красителей Acid Green 27 и Acid Blue 80 гидроксиметансульфинатом натрия и диоксидом тиомочевины в аэробных условиях в щелочной среде (Рис. 7).
|
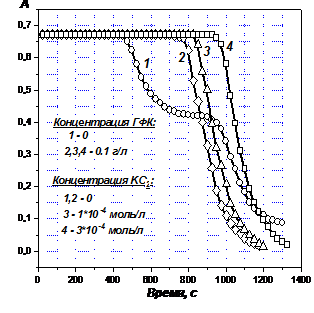
В этой связи оценено влияние ГФК на восстановленные формы кислорода. Как следует из зависимостей на Рис. 8, повышение концентрации О2- или HО2- при постоянной концентрации ГФК приводит к пропорциональному удлинению первого ИП, второй ИП не появляется. Следовательно, ГФК функционируют как эффективные катализаторы диспропорционирования О2- и HО2- в щелочных средах. Об этом
 свидетельствует и полное соответствие количества выделяющегося кислорода стехиометрическим схемам реакций диспропорционирования О2- и пероксида:
свидетельствует и полное соответствие количества выделяющегося кислорода стехиометрическим схемам реакций диспропорционирования О2- и пероксида:
2 HO2 → O2 + H2O2 (6)
2 H2O2 → O2 + 2 H2O (7)
|
Способность ГФК диспропорционировать активные формы кислорода очень значима для технологий колорирования тканей кубовыми красителями, поскольку, препятствуя накоплению активных форм кислорода, ГФК направляют восстановитель на взаимодействие с красителем. Итогом является снижение расхода восстановителя при одновременном увеличении степени фиксации красителя в волокне. Так, одна и та же степень фиксации красителя в волокне в случае печатания ткани традиционным составом достигается при содержании ГМС 100 г/кг, а составом, в который дополнительно введены ГФК (в количестве 0.5 г/кг) – 50 г/кг (Табл. 2).
Таблица 2
Влияние содержания ГФК в печатном составе
на степень фиксации в волокне красителя Кубового ярко-зеленого Ж
Содержание ГФК, г/кг | Степень фиксации красителя (%) при содержании ГМС в печатном составе | |
100 г/кг | 50 г/кг | |
– | 88.0 | 61.2 |
0.05 | 97.5 | 81.3 |
0.10 | 98.5 | 88.0 |
0.25 | 99.7 | 92.1 |
0.50 | 99.8 | 97.5 |

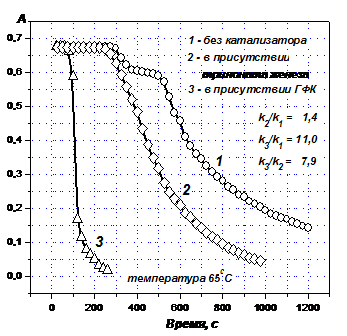 Для подтверждения участия в катализе реакции восстановления красителей обоих типов активных центров, те центры ГФК, которые представляют собой связанные в комплексы ионы переходных металлов (98 % из них составляет железо), моделировались с помощью трилоната железа, при этом концентрация ионов железа в трилонате равнялась их концентрации в ГФК. Как следует из данных на Рис. 9, введение в реакционную среду трилоната железа увеличивает константу скорости восстановления в 1.4 раза, тогда как введение ГФК – в 11 раз. Следовательно, за счет хинонных (карбонильных) групп указанный параметр увеличивается в 7.9 раза.
Для подтверждения участия в катализе реакции восстановления красителей обоих типов активных центров, те центры ГФК, которые представляют собой связанные в комплексы ионы переходных металлов (98 % из них составляет железо), моделировались с помощью трилоната железа, при этом концентрация ионов железа в трилонате равнялась их концентрации в ГФК. Как следует из данных на Рис. 9, введение в реакционную среду трилоната железа увеличивает константу скорости восстановления в 1.4 раза, тогда как введение ГФК – в 11 раз. Следовательно, за счет хинонных (карбонильных) групп указанный параметр увеличивается в 7.9 раза.
3.3 Сравнение эффективности каталитического действия ГФК торфа
и синтетических катализаторов, традиционно используемых в технологиях крашения текстильных материалов кубовыми красителями
Проведена сравнительная оценка эффективности каталитического действия гумусовых кислот и двух других традиционно используемых при колорировании тканей кубовыми красителями катализаторов, b–антрахинонмоносульфокислоты (катализатор 1) и 1,2-дигидроксиантрахинон-3-сульфокислоты (катализатор 2), на реакцию восстановления красителей. Сопоставление велось по времени половинного восстановления красителей (Табл. 3) и по изменению константы скорости реакции восстановления (Табл. 4).
Таблица 3
Влияние катализаторов на время половинного восстановления красителей
Acid Green 27 и Acid Blue 80 гидроксиметансульфинатом натрия при 650С
Катализатор | Скат-ра, 105, моль/л | t1/2, с | t1/2 некат/t1/2 кат | ||
Acid Green | Acid Blue | Acid Green | Acid Blue | ||
– | – | 630 | 750 | – | – |
1 | 0.5 | 350 | 500 | 1.8 | 1.5 |
2 | 0.5 | 200 | 300 | 3.15 | 2.5 |
ГФК | 0.5 | 100 | 200 | 6.3 | 3.75 |
Таблица 4
Влияние катализаторов на параметры кинетических кривых
восстановления красителей
гидроксиметансульфинатом натрия при 650С
Катализатор | Скат-ра, 106, моль/л | Константа скорости реакции (k), ·103, с-1 | kкат/kнекат | ||
Acid Green | Acid Blue | Acid Green | Acid Blue | ||
– | – | 3.7 | 3.6 | – | – |
1 | 0.42 | 4.6 | 4.3 | 1.25 | 1.19 |
3.10 | 11.0 | 6.8 | 2.98 | 1.89 | |
6.10 | 29.6 | 13.0 | 8.00 | 3.60 | |
10.0 | 21.8 | 9.8 | 5.89 | 2.50 | |
2 | 3.2 | 4.5 | 3.9 | 1.18 | 1.09 |
8.2 | 7.7 | 4.4 | 2.08 | 1.23 | |
16.7 | 25.0 | 12.2 | 6.70 | 3.40 | |
18.0 | 17.3 | 8.1 | 4.68 | 2.25 | |
ГФК | 0.4 | 4.9 | 4.2 | 1.30 | 1.17 |
2.0 | 10.6 | 6.4 | 2.86 | 1.78 | |
4.0 | 40.8 | 14.1 | 11.00 | 4.00 |
Данные по времени половинного восстановления красителей свидетельствуют о том, что при одной и той же концентрации гумусовые кислоты торфа эффективнее b–антрахинонмоносульфокислоты в 2-3.5 раза, а 1,2-дигидроксиантрахинон-3-сульфокислоты – в 1.5-2 раза.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
