

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
УДК: 612.821
Методика регистрации сети НР с помощью фМРТ
, ,
В работе представлена адаптированная к фМРТ методика регистрации активности сети негативности рассогласования (НР) при пассивном восприятии звуков речи в режиме odd-ball. Показано, что в условиях разработанной парадигмы активируются области, соответствующие представлениям об организации сети НР – в правой и левой верхневисочной извилинах билатерально, а также в лобной и теменной долях только справа. Обосновывается применимость данной методики как для здоровых испытуемых, так и для клинических исследований.
Ключевые слова: фМРТ; негативность рассогласования (НР); odd-ball–парадигма
In this paper we present the methodology of mismatch negativity (MMN) network registration for passive perception of speech sounds in the odd-ball mode adapted to the fMRI. It was shown that under the conditions of the developed paradigm activated regions corresponded to the knowledge about MMN networking and located in the right and left supratemporal gyri and in the right frontal and parietal lobes. Thus this methodology can be useful for both healthy subjects and in clinical studies.
Key word: fMRI; mismatch negativity (MMN); odd-ball–paradigm
Введение
Негативность рассогласования (НР) – компонент вызванных потенциалов (ВП), возникающий в ответ на редкие девиантные стимулы, предъявляемые в ряду частых «стандартных» (odd-ball–парадигма), с пиковой латентностью в 150-250 мс. Впервые этот компонент был описан психофизиологом Ристо Наатаненом [1, 2]. НР считается отражением автоматического процесса констатации изменений, происходящих в акустической среде [3]. НР может быть получена в ответ на любые различимые изменения слуховых раздражителей, таких, например, как частота или длительность сигнала [4], сложные музыкальные звуки [5] и различные речевые стимулы [6, 7].
НР является одним из эффективных электрофизиологических индексов распознавания звуков, в том числе, речевых. Данный индекс отражает процесс нейронного рассогласования между девиантной слуховой импульсацией, или сенсорным «входом» и «следом» сенсорной памяти о стандартных стимулах (рис. 1).
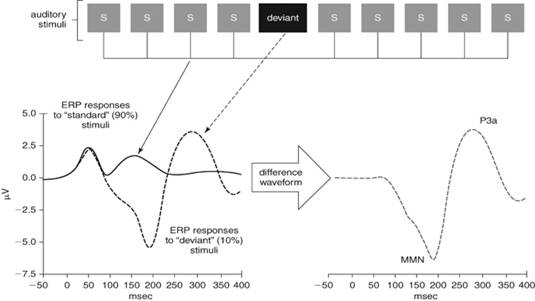
Рис. 1. Схема получения негативности рассогласования как компонента ВП (из Light, G. A. et al., 2010) [8]
Этот автоматический процесс предположительно играет центральную роль в констатации изменений, происходящих в акустической среде [3). НР может быть инициирована у человека даже при отсутствии внимания к предъявляемым слуховым стимулам [2].
Независимость от задачи делает НР неоценимым электрофизиологическим показателем восприятия слуховой информации не только в норме, но и в клинике. НР является полезным инструментом при исследовании пациентов с нарушениями речи (оценка состояния различительной способности слухоречевой системы), внимания или сознания, с трудностями в понимании или выполнении инструкций (когнитивные нарушения, кома, синдром запертого человека и т. д.) [2].
Однако использование ВП для диагностики в клинике ограничивается их неопределенной локализацией, возникающей из-за распространения электрического сигнала при записи ЭЭГ от скальпа.
В последние десятилетия в нейронауках активно применяется метод функциональной магнитно-резонансной томографии (фМРТ), позволяющий локализовать с миллиметровой точностью структуры мозга, активируемые при протекании тех или иных психических процессов. Благодаря данной методике удалось значительно расширить представления о топографии большинства когнитивных процессов, а также выявить их особенности у различных групп испытуемых.
Наиболее изучаемым при фМРТ-сканировании является сигнал, зависящий от уровня оксигенации крови - BOLD (blood oxygenation level-dependent contrast). BOLD-сигнал обратно пропорционален локальному содержанию дезоксигемоглобина в крови, который является парамагнетиком, в отличие от диамагнетиков – оксигемоглобина и ткани мозга [9]. Вслед за возрастанием нейронной активности и, соответственно, увеличением потребления кислорода (кратковременно возрастает локальная концентрация дезоксигемоглобина, BOLD-сигнал кратковременно падает), наблюдается локальное увеличение артериального мозгового кровотока (при этом содержание оксигемоглобина превышает потребность в нем), что сопровождается снижением относительной концентрации дезоксигемоглобина в капиллярах, венулах и дренирующих венах и, соответственно, увеличением BOLD-сигнала [10]. В среднем в норме пик амплитуды BOLD-сигнала приходится на 5 секунд от начала стимула, а длительность всего комплекса составляет 16 секунд (рис. 2).
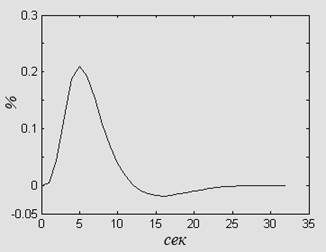
Рис. 2. Схематическое изображение гемодинамического BOLD-ответа на единичное событие малой длительности.
Следует отметить, что при высоком пространственном разрешении, среднее временное разрешение при фМРТ сканировании на томографе мощностью 1,5 Тл – 2 секунды, поэтому особое внимание должно уделяться разработке стимуляционной парадигмы, учитывающей, в том числе, временные характеристики BOLD-ответа (рис. 2).
Целью настоящего исследования было разработать методику регистрации негативности рассогласования на речевые стимулы, адаптированную к функциональной МРТ и пригодную для клинических исследований.
Материалы и методы
Работа проводилась на базе Центра патологии речи и нейрореабилитации (ГБУЗ ЦПРиН ДЗМ). Выборка содержала 25 человек, (12 мужчин, 13 женщин), правши, в возрасте 63.5 [42.5; 73] лет, без признаков заболеваний центральной или периферической нервной системы (согласно клиническим и инструментальным данным).
Учитывая перспективы применения методики в клинике, предполагалось пассивное выделение ответа на стимулы. Испытуемым была дана инструкция лежать расслабленно с закрытыми глазами, не считать и не обращать внимания на звуки. Стимуляция производилась бинаурально через наушники, поставляемые в комплектации с презентационным оборудованием (“Invivo”,США).
Все участники исследования подписали информированное согласие, одобренное этическим комитетом ИВНД и НФ РАН и ЦПРиН.
Тестируемая парадигма
В качестве «стандартных» и «девиантных» стимулов использовались слоги русского языка с оппозиционной фонемой /ба/ и /па/ (произнесенные женским голосом, выравненные по основной форманте, одинаковой длительности (340 мс) и громкости (85 дБ)) (рис. 3).
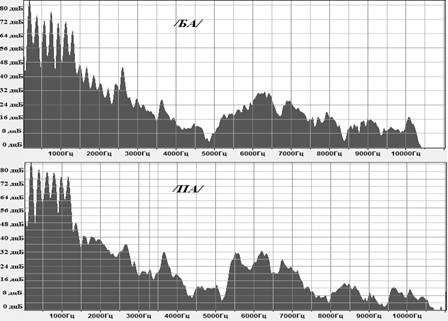
Рис. 3. Графики спектров для стимулов /ба/ и /па/
Для того чтобы добиться точности в локализации НР, парадигма была адаптирована к процедуре фМРТ-исследования и далее будет использоваться термин «негативность рассогласования», что подразумевает его фМРТ-эквивалент.
Стандартная последовательность предъявления стимулов в режиме oddball, применяемая в электрофизиологических исследованиях, была разбита на блоки. Каждый блок парадигмы содержал 10 слуховых стимулов. Слоги /ба/ использовались в качестве стандартных, /па/ - в качестве девиантных (предъявляемых с частотой 20% в рамках одного блока). В 1/3 последовательностей 2 из 10 стимулов являлись девиантными (на рисунке в рамке), в другой 1/3 блоков все 10 стимулов были только стандартными, последняя треть блоков предназначалась для измерения базового уровня активации без предъявления каких-либо звуков (рис. 4). Блоки предъявлялись в случайной последовательности.
Для обеспечения тишины во время предъявления акустических стимулов использовались удлиненные промежутки (задержка сканирования – 9500 мс, см. ниже) между сканированиями, так как шум сканера может искажать данные активности слуховой коры [11]. При этом, согласно классическим представлениям о временных характеристиках BOLD-сигнала, его регистрация возможна как от стандартных, так и от девиантных стимулов блока (как первых, так и последних в последовательности).

Рис. 4. Схема предъявления стимулов в эксперименте 2, серым обозначены периоды работы сканера (сбора функциональных данных, одного вольюма), в цвете – предъявляемые стимулы (слоги /ба/ и /па/), в рамке – основное задание, без рамки – контрольное
Получение структурных и функциональных изображений
МРТ проводилась на томографе MAGNETOMAVANTO 1.5 Т (“Siemens”, Германия). Для получения анатомического изображения в сагиттальной плоскости использовалась последовательность Т1MPRAGE (TR –1900 мс, TE– 3.4 мс, 174 среза, толщина среза – 1 мм, FoV– 250 мм, матрица реконструкции – 256×256 , размер вокселя – 1×1×1 мм).
Процедура сбора функциональных данных была адаптирована к специфике парадигмы. Использовалась последовательность со следующими характеристиками: TR – 12500 мс, задержка – 9500 мс, TE – 50 мс, 32 среза, толщина среза – 3 мм, FoV – 192 мм, матрица – 64×64, размер вокселя – 3×3×3 мм. Исследование включало сбор 90 измерений (объемов) для каждого участника.
Статистические методы анализа данных
Анализ фМРТ-данных проводили с помощью пакета статистической обработки SPM8 [12] на платформе MATLAB 7.0.4 (MathWorks, Natick, MA, USA) на индивидуальном и групповом уровнях. На индивидуальном уровне два первых функциональных объема исключались из анализа, оставшиеся изображения выравнивались относительно первого объема для коррекции движений. Далее проводили корегистрацию среднего функционального изображения со структурным. Процедуру пространственной нормализации в обеих группах проводили с помощью инструмента DARTEL (DiffeomorphicAnatomicalRegistrationThroughExponentiatedLieAlgebra) и NewSegment. Последним этапом предобработки данных являлось сглаживание изображения с помощью фильтра Гаусса с ядром фильтра размером 6×6×6 мм.
Статистические параметрические карты строились с помощью общей линейной модели (GLM) [13]. На групповом уровне результаты t-статистики (SPM{t}) были трансформированы в нормальное стандартное распределение (SPM{z}). Воксельный порог активации соответствовал pFWEcorr<0,05 с поправкой на множественные сравнения, z<3.09. Кластерный порог активации определялся при некорректированном для множественных сравнений уровне puncorr<0.05, минимальный кластерный порог составлял 200 вокселей.
Для построения индивидуальных карт активации использовали контраст, направленный на выявление компонента НР, т. е. производили вычитание активации от стандартных последовательностей из активации от девиантных (D-S). Кластерная поправка вносилась только при построении групповых карт активации, на индивидуальном уровне данная корректировка не производилась. При определении локализации областей активации по координатам использовали приложение Talairach Deamon [14].
Результаты исследования
Групповые карты активации
При построении контраста НР, т. е. при вычитании функциональной активации на стандартные предъявления из активации на девиантные предъявления (D-S) и, таким образом, вычисления сетей, ответственных за феномен НР, были выявлены гемодинамически активные области в правой и левой верхневисочной извилинах (ВВИ), в области правой нижней лобной и правой теменной извилин (рис. 5).
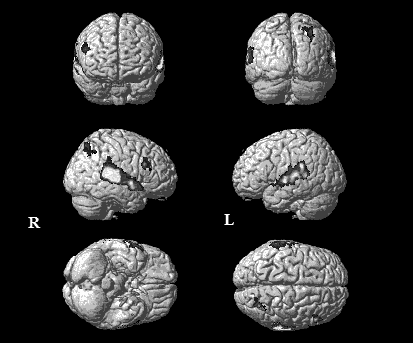
Рис. 5. Групповые карты активации у испытуемых при вычислении контраста НР, Т> 3.14 (рuncorr<0.001), FDRc=200
Объем активации в верхней височной извилине составил (поля 21, 40, 42 по Бродману) слева и справа 4828 и 6993 вокселей соответственно. Соотношение лево - и правосторонней активации составило 1:1,44.
В правой лобной доле активировались поле 9 в нижней и поле 46 в средней лобной извилинах. Также имела место активация в правой теменной доле (табл. 1).
Таблица 1
Области активации при построении группового контраста у здоровых испытуемых
Область коры | Координаты кластера x,y,z | Объем, воксели | Поле по Бродману |
Правая височная доля | 66, -19, 7 | 6993 | 21, 42 |
Левая височная доля | -62, -10, -2 | 4828 | 21, 40, 42 |
Правая лобная доля | 53, 21, 25 | 409 | 9, 46 |
Правая теменная доля | 32, -64, 45 | 452 | 7 |
Обсуждение результатов
Полученная в настоящем исследовании локализация активации соответствует представлениям об организации сети НР. Известно, что к ней относятся области первичной и вторичной слуховой коры, а также нижней и средней лобной извилины.
При построении контраста негативности рассогласования в группе здоровых добровольцев были выявлены наиболее крупные гемодинамически активные области в правой и левой ВВИ (рис. 5). Эти данные соответствуют исследованиям локализации генераторов НР, которые показали, что основной источник НР находится в верхневисочной слуховой коре [15, 16], хотя локализация генератора НР в слуховой коре и зависит от частотных и временных характеристик звука [15, 16]. Интракраниальные записи на животных также подтвердили значительный вклад слуховой коры в генерацию НР у макак [17].
Также в настоящем исследовании была получена активация в правой лобной доле. Некоторые авторы связывают переднелобную активность в рамках механизма НР с непроизвольным переключением внимания на изменение стимула, происходящее вне фокуса внимания. Rinne с соавт. (2000) показал, что фронтальные генераторы НР активируются позже, чем генератор в слуховой коре [18].
Следует отметить, что в литературе, среди прочих данных, было показано, что передняя нижняя лобная кора задействована в принятии решений, связанных с анализом фонологической, семантической и синтаксической речевой информации [19].
Однако, в фМРТ-исследовании C. Doeller с соавт. (2003) с применением неречевых стимулов также была выявлена активация правой нижней лобной извилины (НЛИ), которая в данном случае связывалась авторами с выполнением функции усилителя контраста стандарт – девиант, благодаря наличию обратной зависимости силы этой активации от степени отличия частотных характеристик девианта [20].
Зависимость активации области НЛИ от степени различия между стандартным и девиантным неречевым стимулом также была установлена при последовательной регистрации ЭЭГ и фМРТ [21] и в других исследованиях [22], при этом наблюдалась обратная зависимость объема активации от степени отличия девианта.
Следует отметить, что включение НЛИ, выявляется не всегда. Более того, по данным некоторых авторов, такое включение не является необходимым, а доминирующая роль в распознавании отличий принадлежит правой ВВИ [23]. Такая точка зрения также согласуется с нашими данными, где активация в правой ВВИ почти в полтора раза превосходит активацию в левой ВВИ по объему (табл. 1).
Кроме того, активация самой ВВИ слева и справа по данным M. Sabri с соавт. (2004), выявляется только при достаточно коротком интерстимульном интервале [24].
Другие авторы связывают характеристики (а именно, объем и латерализацию) активации НР с особенностями отличия предъявляемого девианта от стандарта [25].
В настоящем исследовании была получена область активации в правой теменной доле. Включенность этой области в нейрональную сеть НР до настоящего времени дискутируется. Вовлеченность правой теменной доли в генерацию НР подтверждается, в основном, электрофизиологическими исследованиями [26, 27) и только одним фМРТ-исследованием [28].
Активность теменной области также является причиной правостороннего преобладания источников НР [29, 30, 31].
Есть работа, связывающая активность теменной области с фонологическим анализом [32], однако, в рамках исследования НР, такая интерпретация представляется недостаточно правомерной, поскольку в указанных выше исследованиях активация в этой области была получена на тоновые стимулы. Авторы склоняются скорее к роли данной области в механизме НР как субстрата функции мультимодального подпорогового внимания.
Таким образом, полученный в настоящей работе активационный паттерн является характерным для сети НР согласно накопленным данным.
В пользу состоятельности вышеописанной методики свидетельствуют также результаты нашей предыдущей работы с предъявлением тех же стимулов, но в другой последовательности (один раз в 30 секунд в псевдорандомном порядке), где не было выявлено паттерна активации, характерного для сети НР. В этом исследовании в ответ на предъявление тех же речевых стимулов в аналогичной группе здоровых добровольцев выявлялась значительно меньшая по объему (599 и 386 активированных вокселей слева и справа соответственно) корковая активация только в ВВИ с локализацией лишь в полях 21 и 22 по Бродману [33].
Заключение
Разработанная odd-ball–парадигма, адаптированная к ФМРТ последовательности, позволяет выявить области активации, сопоставимые с имеющимися представлениями о генераторах НР, и может, таким образом, быть использована как в экспериментальных, так и в клинических целях.
Работа выполнена при поддержке Российского гуманитарного научного фонда (грант №12-06-00711а).
Литература
1. Näätänen R. The mismatch negativity: a powerful tool for cognitive neuroscience// Ear Hear. 1995. V.16. N1. P. 6-18.
2. Näätänen R., Escera C. Mismatch negativity (MMN): clinical and other applications// Audiol. Neuro–Otol. 2000. N.5. P. 105–110.
3. Näätänen R. The perception of speech sounds by the human brain as reflected by the mismatch negativity (MMN) and its magnetic equivalent (MMNm)// Psychophysiology. 2001. N.38. P. 1– 21.
4. Wunderlich J. L., Cone–Wesson B. K. Effects of stimulus frequency and complexity on the mismatch negativity and other components of the cortical auditory–evoked potential// J. Acoust. Soc. Am. 2001. V.109. P. 1526–1537.
5. Tervaniemi M., Ilvonen T., Sinkkonen J., Kujala A., Alho K., Huotilainen M. Näätänen R. Harmonic partials facilitate pitch discrimination in humans: electrophysiological and behavioral evidence// Neurosci. Lett. 2000. V.279. P. 29–32.
6. Kraus N., McGee T., Sharma A., Carrell T., Nicol T. Mismatch negativity event–related potential elicited by speech stimuli// Ear Hear. 1992. V.13. P. 158–164.
7. Näätänen R., Lehtokoski A., Lennes M., Cheour M., Huotilainen M., Iivonen A., Vainio M., Alku P., Ilmoniemi R. J., Luuk A., Allik J., Sinkkonen J., Alho K. Language–specific phoneme representations revealed by electric and magnetic brain responses// Nature. 1997. V.385. P. 432–434.
8. Light G. A., Williams L. E., Minow F., Sprock J., Rissling A., Sharp R., Swerdlow N. R., Braff D. L. Electroencephalography (EEG) and event-related potentials (ERPs) with human participants.//Curr Protoc Neurosci. 2010. V.6. N 6 P. 25.1-24.
9. Ogawa S., Lee T. M., Kay A. R., Tank D. W. Brain magnetic resonance imaging with contrast dependent on blood oxygenation. // Proc. Natl. Acad. Sci. U S A. 1990. V. 87. N 24. P. 9868-9872.
10. Buxton R. B., Uludağ K., Dubowitz D. J., Liu T. T. Modeling the hemodynamic response to brain activation //Neuroimage. 2004. V. 23. N. 1. P. 220-233.
11. Novitski N., Alho K., Korzyukov O., Carlson S., Martinkauppi S., Escera C., Rinne T., Aronen H. J., Naatanen R. Effects of acoustic gradient noise from functional magnetic resonance imagingon auditory processingas reflected by event-related brain potentials. Neuroimage. 2001. N. 14. P. 244 –251.
12. Welcome. Trust. Centre. forNeuroimaging: http://www. fil. ion. ucl. ac. uk/spm.
13. Friston K. J., Holmes A. P., Worsley K. J., et al. Statistical parametric maps in functional imaging: A general linear approach. //Hum. Brain Mapp. 1994. N 2. P.189–210.
14. Lancaster J. L., Woldorff M. G., Parsons L. M., et al. Automated Talairach Atlas labels for functional brain mapping. //Hum. Brain Mapp. 2000. N 10. P.120-131.
15. Alho K. Cerebral generators of mismatch negativity (MMN) and its magnetic counterpart (MMNm) elicited by sound changes // Ear Hear. 1995. V.16. P. 38-51.
16. Alho K., Tervaniemi M., Huotilainen M., Lavikainen J., Tiitinen H., Ilmoniemi R. J., Knuutila J., Näätänen R. Processing of complex sounds in the human auditory cortex as revealed by magnetic brain responses // Psychophysiology. 1996. V.33. P. 369–375.
17. Javitt D. C., Steinschneider M., Schroeder C. E., Vaughan H. G. Jr., Arezzo J. C. Detection of stimulus deviance within primate primary auditory cortex: intracortical mechanisms of mismatch negativity (MMN) generation// Brain Res. 1994. V.667. P. 192–200.
18. Rinne T., Alho K., Ilmoniemi R. J., Virtanen J., Näätänen R. Separate time behaviors of the temporal and frontal mismatch negativity sources// Neuroimage. 2000. V.12. P. 14–19.
19. Hirshorn E. A., Thompson–Schill S. L. Role of the left inferior frontal gyrus in covert word retrieval: neural correlates of switching during verbal fluency// Neuropsychologia. 2006. V.44. N12. P. 2547– 2557.
20. Doeller C. F., Opitz B., Mecklinger A., Krick C., Reith W., Schröger E. Prefrontal cortex involvement in preattentive auditory deviance detection: neuroimaging and electrophysiological evidence// NeuroImage. 2003. V.20, N2. P. 1270–1282.
21. Opitz B., Rinne T., Mecklinger A., von Cramon D. Y., Schröger E. Differential contribution of frontal and temporal cortices to auditory change detection: fMRI and ERP results// NeuroImage. 2002. V.15, N1. P. 167–174.
22. Rinne T., Degerman A., Alho perior temporal and inferior frontal cortices are activated by infrequent sound duration decrements: an fMRI study// NeuroImage. –2005. – V.26, N1. – P. 66–72.
23. Liebenthal E., Ellingson M. L., Spanaki M. V., Prieto T. E., Ropella K. M., Binder J. R. Simultaneous ERP and fMRI of the auditory cortex in a passive oddball paradigm// NeuroImage. 2003. V.19. N4. P. 1395–1404.
24. Sabri M., Kareken D. A., Dzemidzic M., Lowe M. J., Melara R. D. Neural correlates of auditory sensory memory and automatic change detection// NeuroImage. 2004. V.21, N1. P. 69–74.
25. Molholm S., Martinez A., Ritter W., Javitt D. C., Foxe J. J. The Neural Circuitry of Pre–attentive Auditory Change–detection : An fMRI Study of Pitch and Duration Mismatch Negativity generators// Cerebral Cortex. 2005. V.15, N5. P. 545–551.
26. Kasai K, Nakagome K, Itoh K, Koshida I, Hata A, Iwanami A, et al. Multiple generators in the auditory automatic discrimination process in humans //Neuroreport. 1999. V.10. P. 2267–71.
27. Levänen S, Ahonen A, Hari R, McEvoy L, Sams M. Deviant auditory stimuli activate human left and right auditory cortex differently //Cereb Cortex. 1996. N 6. P.288–96.
28. Schall U., Johnston P., Todd J., Ward P. B., Michie P. T. Functional neuroanatomy of auditory mismatch processing: an event-related fMRI study of duration-deviant oddballs. Neuroimage 2003. V. 20. P. 729–36.
29. Giard MH, Perrin F, Pernier J, Bouchet P. Brain generators implicated in the processing of auditory stimulus deviance: a topographic event-related potential study.//Psychophysiology. 1990. V. 27. N. 6. P. 627-40.
30. Giard MH, Perrin F, Pernier J. Scalp topographies dissociate attentional ERP components during auditory information processing.//Acta Otolaryngol Suppl. 1991. V. 491. P. 168-74.
31. Giard MH, Lavikahen J, Reinikainen K, Perrin F, Bertrand O, Pernier J, Näätänen R. Separate representation of stimulus frequency, intensity, and duration in auditory sensory memory: an event-related potential and dipole-model analysis.//J Cogn Neurosci. 1995. V. 7. N. 2. P. 133-43.
32. McDermott KB, Petersen SE, Watson JM, Ojemann JG. A procedure for identifying regions preferentially activated by attention to semantic and phonological relations using functional magnetic resonance imaging.// Neuropsychologia. 2003. V.41. N 3 P. 293-303.
33. , , Федина , влияющие на характеристики BOLD-ответа у пациентов после инсульта //Вестник РНЦРР. 2013. Т. 4. № 13. с. 18.
The method of MMN network detecting using fMRI
Mayorova L. A. –
Institute of higher nervous activity and neurophysiology of RAS
Centre of speech pathology and neurorehabilitation
Petrushevsky A. G. –
Centre of speech pathology and neurorehabilitation
Fedina O. N. –
Centre of speech pathology and neurorehabilitation
The mismatch negativity (MMN) – a component of evoked response potentials (ERP) with a peak latency of 150-250 ms – occurs in response to rare “deviant” stimuli presented in the sequence of the “standard” (odd-ball-paradigm). The task – independency makes MMN as invaluable indicator of auditory perception in normal subjects and in number of neurological and psychiatry diseases. However, use of the ERP for the diagnosis in the clinic is limited to their uncertain localization, arising from the spread of the electrical signal when recording EEG from the scalp. This problem can be solved by using functional magnetic resonance imaging (fMRI). The aim of this study was to develop a technique for recording mismatch negativity (MMN) adapted to functional MRI and suitable for clinical studies. We took the standard sequence of odd-ball stimuli presentation mode used in the electrophysiological studies and divided it into blocks. Each block of paradigm contained 10 auditory stimuli. Syllables /ba / was used as a “standard” and /pa/ - as “deviant” (with a frequency of 20% in a single block). In 1/3 of sequences 2of 10 stimuli were “deviant”, in another 1/3 blocks all 10 stimuli were only “standard”, the last third of the blocks used to measure baseline activation without presenting any sounds. As a "standard" and "deviant" stimuli the syllables of Russian opposition phonemes /ba/ and /pa/ (pronounced by female voice, aligned along the main formants of the same duration (340 ms) and volume (85 dB) were used. The blocks were presented in random order. The developed paradigm approved on 25 right-handed healthy subjects (12 men, 13 women) in fMRI conditions. The fMRI equivalent of mismatch negativity in passive response to speech phonemes – the syllables "ba" and "pa" – in the odd-ball paradigm was registered. The active areas in the right and left supratemporal gyri (STG), as well as in the right frontal and parietal lobes corresponded to MMN contrast. The STG’s activation was right lateralized as well as in the whole MMN activation pattern. This obtained in the present study pattern of activation consistent with the concept of MMN networking. The developed passive odd-ball–paradigm adapted to fMRI reveals activation regions with available representations of generators MMN and can thus be used in experimental and clinical purposes.
–
к. м.н., научный сотрудник, лаборатория прикладной физиологии высшей нервной деятельности человека, Институт высшей нервной деятельности и нейрофизиологии РАН, 117485, Москва, Бутлерова, 5а
ГБУЗ Центр патологии речи и нейрореабилитации ДЗМ, 109240, Москва, Николоямская, 20
+79057878496
*****@***ru
Mayorova Larisa Alexeevna –
Ph. D., research fellow, Laboratory of applied physiology of human higher nervous activity, Institute of higher nervous activity and neurophysiology of RAS,117485, Moscow, Butlerova, 5a
Centre of speech pathology and neurorehabilitation, 109240, Moscow, Nikoloyamskaya, 20
+79057878496
*****@***ru
–
врач-рентгенолог, ГБУЗ Центр патологии речи и нейрореабилитации ДЗМ,
109240,Москва, Николоямская, 20
+79031194081
*****@***ru
Petrushevsky Alexey Garrievich –
radiologist, Centre of speech pathology and neurorehabilitation, 109240, Moscow, Nikoloyamskaya, 20
+79031194081
*****@***ru
–
врач-рентгенолог, ГБУЗ Центр патологии речи и нейрореабилитации ДЗМ,
109240, Москва, Николоямская, 20
+79197709467
*****@***ru
Fedina Oksana Nikolaevna –
radiologist, Centre of speech pathology and neurorehabilitation, 109240, Moscow, Nikoloyamskaya, 20
+79197709467
*****@***ru
