

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Моделирование морфогенеза Arabidopsis thaliana в терминах клеточного автомата.
1*, 2, 1, 2, 1, 1
1-Институт Цитологии и Генетики СО РАН, Новосибирск, Россия
2-Новосибирский Государственный Университет, Новосибирск, Россия
e-mail: *****@***nsc. ru, *Автор для связи
Ключевые слова: клеточный автомат, математическая модель, развитие меристемы побега, параметры модели, период деления, база данных.
Краткое изложение
Развитие организма является чрезвычайно сложным процессом, в котором происходит интеграция большого количества генных сетей разных типов клеток. Разработка математической модели, дает возможность проведения численного анализа регуляторных механизмов, управляющих морфодинамикой клеточных ансамблей. В работе представлена модель клеточного автомата, моделирующего эмбриональное развитие меристемы побега Arabidopsis thaliana. Представлены результаты анализа влияния мутаций, приводящие к тому, что а) клетки меристема дифференцируется в срощенные зародышевые листки и растение дальше не развивается; б) вместе с дифференцианцией клеток меристемы уменьшается зона меристемы.
Введение
Постэмбриональное развитие наземной части высших растений определяется активностью апикальной меристемы побега, динамической структуры, образующей листья, цветы и стебель. Формирование апикальной меристемы побега происходит на самых ранних стадиях эмбриогенеза. Далее, функционирование промеристемы определяет развитие зародышевых листочков, паралельно с развитием которых происходит также и формирование сложной структуры меристемы (рис. 1) [2]. В 16-клеточном зародыше, клетки апикальной и базальной части уже различаются экспрессией различных генов. Без меристемы побега, дальнейшее развитие зародыша, по крайней мере, апикальной его части, невозможно [3].
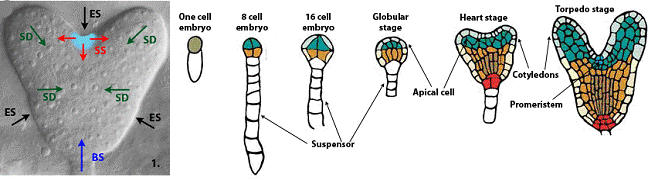 |
Рис 1. (1) Направления распределения гипотетических сигналов модели в ткани heart-stage embryo: ES - External Signal, SS - Stem Signal, SD - Signal of Differentiation and BS - Basal Signal. (2) Стадии развития зародыша растения с указанием значимых для модели органов и тканей.
На основе данных, представленных в базе AGNS разработана математическая модель развития меристемы побега Arabidopsis thaliana в эмбриогенезе. В модели описаны процессы закладки меристемы, формирования ее сложного строения и функционирования. Зародыш представляется в рамках данного подхода как двумерный массив клеток, темпы делений которого, зависят от клеточного окружения. Модель построена в терминах клеточного автомата. Клетки автомата соответствуют отдельным клеткам моделируемой области растения. Каждая клетка способна воспринимать сигналы от других клеток и, в зависимости от типа клетки, продуцировать сигналы, которые могут воспринимать другие клетки модели. Биологическим смыслом сигналов являются концентрации определенных диффундирующих веществ, морфогенов, имеющих специфическое влияние на клетку.
Методы и алгоритмы
В модели предполагается, что состояние и темпы деления отдельной клетки зависят от влияния на нее сигналов, распространяющихся от других клеток зародыша. В рамках модели было выбрано 4 биологически-обоснованных сигнала, которые однозначно моделируют морфодинамику ткани растения на принятом уровне абстракции (рис. 1).
Все клетки в модели зародыша растения делятся на восемь типов:
1. Null - тип клеток, не продуцирующих никакого сигнала и неспособных к делению.
2. NullEx - тип клеток эпидермального слоя. NullEx продуцируют ES и располагаются по всему периметру зародыша. Клетки этого типа также не способны к делению, но мы искусственно окружаем модель зародыша клетками данного типа.
3. NullSus - клетки суспензора. Эти клетки продуцируют BS и располагаются в нижней части зародыша. В модели их всегда две.
4. Lateral - клетки этого типа имитируют «дифференцированные» клетки, которые продуцируют SD.
5. Promeristem - клетки меристемы зародыша. Эти клетки продуцируют SS и находятся в верхней части зародыша. В процессе развития эти клетки переходят в клетки типа L2meristem и L3meristem.
6. Transit - клетки, находящиеся вблизи меристемы. Они так же продуцируют низкий SD, но имеют самый высокий темп деления
7. L2meristem - клетки меристемы, находящиеся во втором слое (считая от эпидермального слоя) верхней части зародыша. Эти клетки продуцируют SS.
8. L3meristem - клетки, находящиеся на слой ниже клеток типа L2meristem. Так же продуцируют SS.
Каждая клетка имеет ряд внутренних параметров, которые характеризуют ее состояние:
1. Type – тип клетки.
2. ExternSignal0, BasalSignal0, StemSignal0, DifferentSignal0 – значения сигналов продуцируемых данной клеткой.
3. ExternSignal, BasalSignal, StemSignal, DifferentSignal - значения сигналов, которые данная клетка воспринимает.
4. Kij – характеристика состояния клетки с координатами (i, j) , вычисляемая как отношение SS к SD.
5. Tij– период деления клетки, который зависит от текущего значения характеристики Kij.
6. Tpij – количество итераций прошедших после последнего деления (i, j)-ой клетки.
Совокупное влияние всех клеток на клетку с координатами (i, j) мы рассчитываем по формулам
![]() ,
,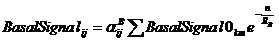 ,
,
![]() ,
,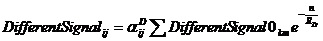 .
.
Суммирование ведется по всем клеткам ткани, включая клетки эпидермального слоя и клетки суспензора; (k, m) – координаты клетки, влияние которой мы учитываем, n=max(abs(k-i),abs(m-j)) – относительный номер слоя (т. е. рассматриваемая клетка является нулевым слоем, прилежащие клетки являются первым слоем, прилежащие клетки к первому слою являются вторым слоем и т. д.); ![]() - константы, характеризующие проницаемость (коэффициент диффузии) для ES, BS, SS, и SD соответственно; константы
- константы, характеризующие проницаемость (коэффициент диффузии) для ES, BS, SS, и SD соответственно; константы ![]() - характеризуют восприимчивость клетки к определенному типу сигнала, эта константа зависит только от типа клетки. В текущий момент времени рассматриваемая клетка находится в определенном состоянии, которое характеризуется параметром
- характеризуют восприимчивость клетки к определенному типу сигнала, эта константа зависит только от типа клетки. В текущий момент времени рассматриваемая клетка находится в определенном состоянии, которое характеризуется параметром ![]() .
.
Качественное поведение функции Tij в соответствии с биологической постановкой, отражено на (рис. 2):
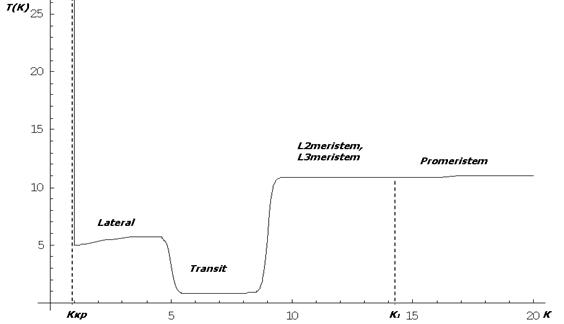 |
Рис. 2. Зависимость периода деления от параметра К для различных типов клеток
Из рис.2 видно, что клетки меристемы зародыша (Promeristem, L2meristem, L3meristem) в ткани делятся медленнее всех, клетки типа Transit делятся очень быстро, темпы делений клеток типа Lateral на среднем уровне. При достижении параметром ![]() значения
значения ![]() период деления уходит на бесконечность и клетка впредь больше не делится. При достижении в клетке типа Promeristem параметра K порогового значения
период деления уходит на бесконечность и клетка впредь больше не делится. При достижении в клетке типа Promeristem параметра K порогового значения ![]() клетка делится на L2meristem и L3meristem. Такую зависимость мы установили с помощью функции вида
клетка делится на L2meristem и L3meristem. Такую зависимость мы установили с помощью функции вида ![]() . Если Tij – не целое число, то функция Tij округляется согласно стандартным правилам округления. Решение о делении клетки принимается на основании сравнения значений Тpij и Tij. Для каждой не делящейся клетки последующая итерация будет увеличивать значение Tpij на 1. Если Тpij>Tij, то принимается решение о делении клетки на две дочерние. Для каждой дочерней клетки Tpij полагается равным 0. Отметим следующее очевидное свойство описанной процедуры: Если Тij растёт быстрее, чем Tpij, то клетка не будет делиться. Направление деления зависит от типа клетки и ее ближайших соседей.
. Если Tij – не целое число, то функция Tij округляется согласно стандартным правилам округления. Решение о делении клетки принимается на основании сравнения значений Тpij и Tij. Для каждой не делящейся клетки последующая итерация будет увеличивать значение Tpij на 1. Если Тpij>Tij, то принимается решение о делении клетки на две дочерние. Для каждой дочерней клетки Tpij полагается равным 0. Отметим следующее очевидное свойство описанной процедуры: Если Тij растёт быстрее, чем Tpij, то клетка не будет делиться. Направление деления зависит от типа клетки и ее ближайших соседей.
Результаты
 Разработана математическая модель меристемы побега Arabidopsis thaliana в эмбриогенезе (рис. 3).
Разработана математическая модель меристемы побега Arabidopsis thaliana в эмбриогенезе (рис. 3).
Рис. 3. Стадии развития меристемы побега, моделируемые клеточным автоматом
![]()
Подобраны параметры математической модели для адекватного описания функционирования системы. С помощью клеточного автомата исследованы следующие мутации, описанные в литературе и влияющие на развитие меристемы побега:
а) меристема образуется, но так как не может справиться с потоком ауксина из зародышевых листков, то дифференцируется, что приводит к формированию срощенных зародышевых листков и растение дальше не развивается [4];
б) меристема образуется, зародышевые листки формируются, но при этом, не осуществляется регуляция размеров пула клеток меристемы. Это приводит к тому, что клетки меристемы дифференцируются и зона меристемы уменьшается [5; 2].
На основе разработанного клеточного автомата был создан визуализатор модели, с помощью которого можно задавать любое начальное состояние ткани и значения параметров модели, и уже на основе их, наблюдать визуализированное развитие меристемы побега (рис. 4).
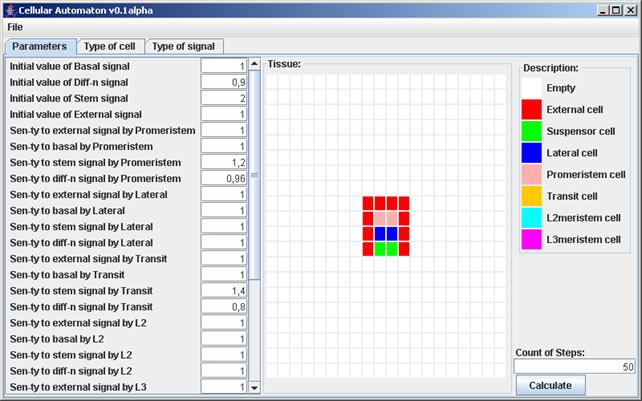
Рис. 4. Визуализация разработанного клеточного автомата
При этом можно наблюдать не только развитие ткани, но и распределение биологических сигналов по ткани, что является важной частью при анализе расчёта модели экспертом (рис. 5).
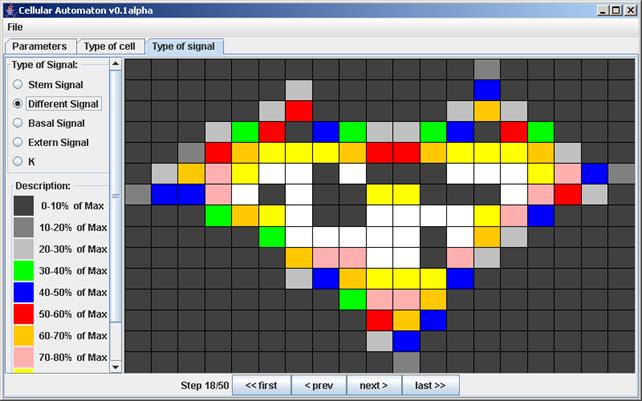
Рис. 5. Распределение сигнала дифференцировки
Обсуждение
Для исследования механизмов развития тканей растений необходимо создание специализированных баз данных, содержащих генетическую информацию об исследуемой системе. На основе экспериментальных данных, содержащихся в базе AGNS, создана математическая модель в терминах клеточного автомата, имитирующего морфодинамику развития зародыша под действием сигналов, продуцируемых отдельными клетками зародыша, что является первым этапом в моделировании процессов развития, в общем, и генных сетей морфогенеза, в частности. Развитие меристемы растения в эмбриогенезе, характеризуется сочетанием процессов лавинообразного развития дифференцирующейся ткани, и стабильного развития стволовых клеток ткани. Оба эти процесса были промоделированы в созданном клеточном автомате. Такой автомат позволит не только предсказывать динамику процессов деления и дифференцировки клеток, протекающих в рассматриваемой системе, но также анализировать влияние реальных мутаций на ее функционирование. Хотя описание функционирования такой ткани, как меристема побега, с помощью клеточного автомата является грубым, всё-таки его создание - необходимый и важный шаг на пути к созданию математической модели, описывающей развитие меристемы побега, с учётом генетической регуляции. В дальнейшем планируется развитие клеточного автомата, включающее моделирование различных, экспериментально полученных мутаций и усложнение определения характеристик клетки для более адекватного описания процесса развития тканей растений.
[1] Omelyanchuk N. et al. (2005) AGNS-A DATABASE ON EXPRESSION OF ARABIDOPSIS GENES. Bioinformatics of Genome Regulation and Structure II. Springer Science+Business Media, Inc. 433-442.
[2] Sharma V. K. and Fletcher J. C. (2003). Maintenance of Shoot and Floral Meristem Cell Proliferation and Fate. PNAS. 100. 11823- 11829.
[3] Kaplan D. R. and Cooke T. J. (1997). Fundamental concepts in the embryogenesis of Dicotyledons: a morphological interpretation of embryo mutants. The Plant Cell. 9. 1903- 1919.
[4] Aida M., Ishida T., Tasaka M. (1999). Shoot apical meristem and cotyledon formation during Arabidopsis embryogenesis: Interaction among the CUP-SHAPED COTYLEDON and SHOOT MERISTEMLESS genes. Development. 119. 823–831
[5]Clark S. E., Running M. P., Meyerowitz E. M. (1995) CLAVATA3 is a specific regulator of shoot and floral meristem development affecting the same processes as CLAVATA1. Development. 121. 2057- 2067.
[6] Jaiswal P. et al. (2005) Plant Ontology (PO): a controlled vocabulary of plant structures and growth parative and Functional Genomics. 6. 388- 397.
