

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
УДК 547.466 + 547.269.81
ПРИРОДНЫЕ ОРГАНИЧЕСКИЕ СОЕДИНЕНИЯ СЕЛЕНА
Кафедра общей и органической химии КГМУ, Казань, 420012,
Россия. Е-mail: *****@***ru
Моему учителю, профессору Катаеву Евгению Геннадьевичу
посвящается, большую часть своей научной деятельности
селеноорганическим соединениям посвятившему.
Ключевые слова: селена соединения, аминокислоты, селенопротеины.
Резюме
Обзор посвящён актуальной в настоящее время селеновой проблеме, связанной с влиянием его природных органических соединений на здоровье человека. Приведены структуры всех основных биогенных соединений этого элемента, их биосинтез и метаболизм, содержание в природных источниках растительного и животного царств, медико-биологические свойства селеноцистеина и селенометионина, главных представителей этого класса.……
Вместо введения
Несколько десятилетий тому назад, все мы, студенты и аспиранты кафедры органической химии Казанского государственного университета, работавшие под руководством , в той или иной степени были втянуты им в исследования по химии органических соединений селена – работы нелёгкие и мало благодарные. И тогда, в ответ на вопрос о полезности этих веществ, я услышал ответ –“ пока, никакой “. Но вот, не прошло и четверти века, как зазвучал селеноцистеин, за ним – селенометионин ; и пошло поехало – образовалась биохимия селена и появились фармацевтические препараты селена. Возникла проблема селена. Вот я и вспомнил, в котрый раз, это “…пока…”.
Селеноцистеин и его окружение
Первые указания на селенопротеины последовавшие в 1973 году [1-4], естественным образом, инициировали поиски аминокислоты, носителя этого элемента, и к 1978 году завершились открытием селеноцистеина [5-7], который оказался 21-ой протеиногенной аминокислотой – он получил обозначение Sec и для него был найден генетический кодон в DNA в виде триплета TGA ( соответсвенно, UGA в tRNA ) ранее известный как стоп-кодон [8]. Последовавшие за этим работы по химии селеноцистеина (схема 1) были направлены на выяснение его роли в живых организмах, на определение схемы его биосинтеза, на выявление распределения его в природе и, наконец, на влияние его на здоровье человека. Естественно также, что возникли предположения о возможном включении селена и в другие органические природные соединения, не обнаруженные доселе по причине их очень низкой концентрации в природных источниках, что, кстати, следует сказать и о самом селеноцистеине.

Схема 1
Именно низкое содержание селеновых соединений во всех живых организмах и было причиной столь позднего их открытия, и только тогда, когда появились очень чувствительные методы анализа состава сложных смесей веществ с предельно малыми концентрациями компонент, такими как хромато-масс-спектрометрия, напрмер [9] , и был обнаружен ряд соединений этого семейства. Селеноорганические производные были найдены по ходу исследования биосинтеза и метаболизма селеноцистеина, этого ключевого соединения группы природных селен содержащих веществ, а также как аналоги известных серу содержащих природных соединений различных классов. Так были открыты: селенометионин (Se-met) - I, гомоселеноцистеин - II, Se-метилселеноцистеин - III, Se-аденозилселеногомоцистеин - IY, селеноцистатионин – Y [10-11], селенобиотин - YI[12], селеносахара (Se-метилселено-N-ацетил-галактозамин –YII и Se-метилселено-N-ацетил-глюкозамин – YIII ) [13], g-глутамил-Se-метилселеноцистеин – IX [14] , селеносинигрин – Х , которые и представлены на схеме 2.
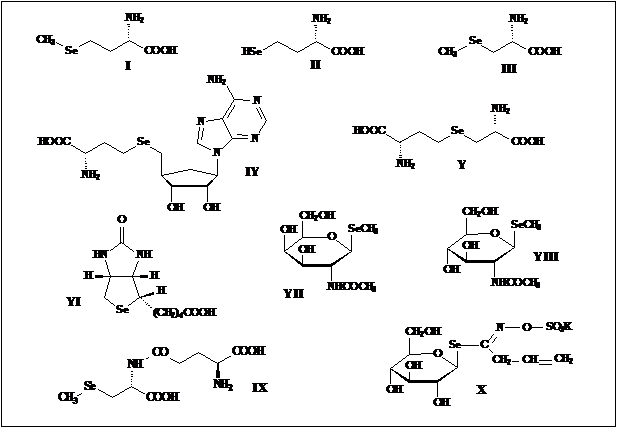
Схема 2
Распространение в природных источниках
Первые органические соединения селена в природных источниках были зафиксированы лишь в 1973 году, хотя указания биологического и медицинского характера на то появились значительно раньше [15-17], а этими соединениями оказались белковые вещества, селенопротеины, найденные во всех типах организмов – от простейших до млекопитающих [1-7, 11, 18] – в которых они выполняют функции энзимов, оксидоредуктаз, в основном. За исключением некоторых бактерий, селен в белковых молекулах находится в виде селеноцистеина, включенного в основную полипептидную цепь.
Ранее всего селенопротеины были обнаружены в низших eukarya подобных трематодам Schistosoma mansoni [19], в плодовых мушках Drosophila melanogaster [20] и Caenorhabditis elegans [21], предполагается достаточно широкое распространение их в низших животных [22],в archaea селенопротеины были обнаружены в родах Methanococcus и Methanopyrus [23].Зафиксирован селенопротеин, в виде глутатион пероксидазы, в зелёных водорослях Chlamydomonas rheinhardtii [24].
Класс селенопротеинов включает бактериальные формиат дегидрогеназы, глицин редуктазы, дейодиназы млекопитающих, тиоредоксин редуктазы, глутатион пероксидазы, включая фосфолипид гидропероксид глутатион пероксидазы. В организме человека они найдены в печени, в плазме крови, в мышцах сердца [25-27].
Низкомолекулярные соединения селена достаточно широко распространены в растениях, в некоторых из них накапливаются в значительных количествах, например, в чесноке и дрожжах – согласно данным метода HPLC-ICP-MS, они содержат селена 296 мг/кг и 1922 мг/кг, соответственно, при этом, в чесноке доминирует g-глутамил-Se-метилселеноцистеин (IX), а в дрожжах – селенометионин (I) [28]. Отмечена склонность к накоплению различных соединений селена у растений рода Astragalus и видов Brassica и Allium ( табл. 1 ).
Таблица 1. Низко молекулярные соединения селена в растениях.
Соединения | Источники | Содержание Se, мг/кг |
Селеноцистатионин (Y) | Astragalus praleongus | 517 [28] |
Se-Метилселеноцистеин (III) | Brassica oleracea botrytis Allium tricoccum Melilotus indica | 345 [29] 48 – 524 [28] 200 [30] |
g-Глутамил-Se-метилселеноцистеин (IX) | Allium sativum | 96 – 114 [28] |
Cеленометионин (I) | Brassica juncea | 138 [28] |
Селенобиотин (YI) | Phycomyces blakesleeanus | [31] |
g-Глутамилселенометионин (XI)
| Allium sativum | 68 – 1355 [28] |
Se-Аденозилселеногомоцистеин (IY) | Sacharomyces cerevisiae | 1200 – 2100 [28, 32-34] |
Селенолантионин (XII)
| Sacharomyces cerevisiae | 1922 [28] |
3-Бутенил изоселеноцианат (XIII)
| Stanleya pinnata | 100 [35] |
Селеносинигрины (X) | Stanleya pinnata | 100 [36] |
Селеносахара (YII-YIII) | Astragalus racemosus | [37] |
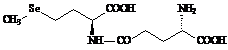
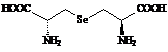
Биосинтез соединений селена.
Содержание селена в земной коре (6х10-5%), а следовательно и в почве, не велико и в чистом виде, или в виде чистых минералов, очень редок – обычно он сопутствует в следовых количествах природные сульфиды вместе с теллуром. Неорганические соединения селена могут усваиваться всеми видами организмов, хотя эффективность биоассимиляции соединений селена существенно зависит от их химической природы и от типа организма [38]. Главные источники селена в почве, селенаты, усваиваются высшими растениями тем же биохимическим путём что и сульфаты – это связано с близостью химических свойств соединений серы и селена, а также с совместным их присутствием в минеральном сырье.
Всю схему биоассимиляции селена можно разделить на три принципиальных этапа: восстановление окисленных неорганических почвенных соединений селена (селенатов и селенитов) до селенидов, транспорт двух-валентного селена в органические молекулы (селеноцистеин), последующие миграции селена в другие биоорганические соединения.
На первом этапе селенаты, биохимически доступные только для наземных растений и бактерий, восстанавливаются до селенитов, которые могут усваиваться всеми организмами. Восстановление селенатов до селенитов и восстановление последних до селенидов осуществляется под контролем как соответствующих сульфо-редуктаз, так и специфических селено-редуктаз, среди которых наиболее универсальными являются глутатион редуктаза и тиоредоксин редуктаза [39, 40]. Соединение двух-валентного селена, селеноглутатион-трисульфид, при гидрогенолизе выделяет селеноводород, скорее всего связанный, или его натриевые соли, которые при взаимодействии с АТР образуют селенофосфат – этот последний может быть назван “активным селенидом”, поскольку именно он участвует в реакции введения селена в молекулу серина, что и приводит к селеноцистеину (схема 3).
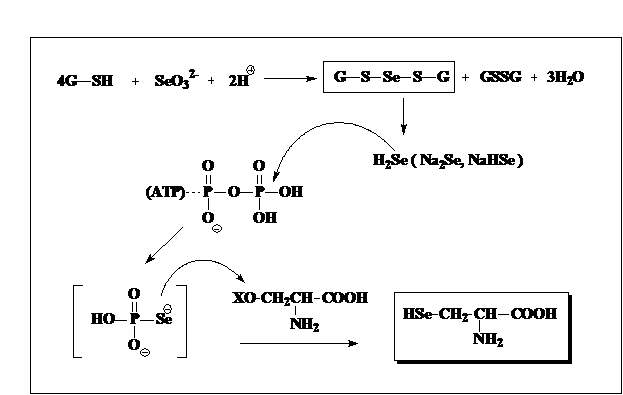
Cхема 3
Следует отметить высокий нуклеофильный характер двухвалентного селена во всех интермедиатах биосинтетического пути ведущего к селеноцистеину, но также надо отметить, что эта нуклеофильность сохраняется и в самом селеноцистеине, которая открывает путь к последующим реакциям нуклеофильного замещения с его участием в качестве нуклеофила и участием гомосерина как электрофила. Полученная таким образом молекула селеноцистатионина после восстановления образует селеногомоцистеин, а последний метелируется уже до селенометионина (схема 4). Эта биосинтетическая схема характерна для наземных растений, водорослей, пивных дрожжей и определяет высокую степень накопления селена у них в виде метионина.

Схема 4
Медико-биологические свойства природного органического селена.
Из выше рассмотренного материала вполне очевидно, что селеноцистеин является чуть ли не единственным значимым природным соединением селена, а все остальные найденные в природных источниках либо лежат на пути его биосинтеза (интермедиаты), либо являются его метаболитами. Сам же селеноцистеин, в какой то степени и селенометионин, включены в состав белковых молекул, обычно энзимов, и выполняют свои биологические функции в виде селенопротеинов.
В молекулах селенопептидов селенольная функция остаётся свободной и выполняет свою ключевую антиоксидантную функцию, по степени активности не имеющей себе равных среди других биоантиоксидантов. Для живых клеток наибольшую опасность представляют гидроксильные свободные радикалы, образующиеся при разложении органических гидроперекисей, ответственные за разрушение клеточных мембран, провоцирующие мутагенез и канцерогенез. Лучшими перехватчиками этих радикалов и являются селенопротеины. При этом селенопротеины не расходуются в редокс процессах, а регенерируются взаимодействием с цистеинпротеинами, такими как глутатион редуктаза и теродоксин редуктаза (схема 5) , поэтому любому организму, организму человека в том числе, необходимо очень малое количество органического селена ( Homo sapiens, в норме, при весе около 80 кг, содержит 0.004 мг этого элемента, а суточная норма 50-70 мкг его вполне достаточна).
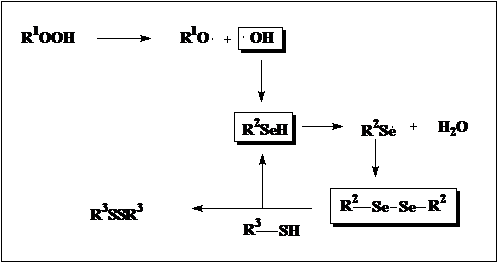
Схема 5
Каково же в принципе влияние селена на здоровье млекопитающих, человека в первую очередь, если он является обязательным микро элементом в нашем рационе? Другими словами, необходима информация о том, как сказывается его недостаток и избыток на состоянии нашего организма, какова токсикология различных соединений селена.
По токсикологии соединений селена были получены следующие данные (LD50, мг/кг, крысы): селенат натрия (1.6), селенит натрия (7.0), селендисульфид (138), селенсульфид (38) – это значит, что соединения двух-валентного селена значительно менее токсичны сравнительно с соединениями селдена(IY) и селена(YI)41. Кроме того установлена тенденция соединений селена к накоплению в организме – он медленно выводится с продуктами метаболизма – откуда следует, что злоупотреблять селено содержащими препаратами не следует ( симптомы отравления селеном – выпадание волос и ногтей, дерматит ). Селеновое отравление наблюдалось у крупного рогатого скота на пастбищах с высоким содержанием селена в почве, который эффективно накапливался астрагалами, там произрастающими.
С другой стороны, надёжно установлено, что хронический недостаток селена в первую очередь сказывается на сердечной деятельности – ведёт к кардиомиопатии (любые структурные и функциональные изменения миокарда сердечных желудочков). Высокая антиоксидантная активность селеноцистеина и селенопротеинов стимулировали широкий спектр медико-биологических исследований с привлечением их в качестве терапевтических веществ, в результате которых была показана определённая эффективность их в качестве разноплановых протекторов, что в общем то и естественно для соединений с такими биохимическими характеристиками (схема 6).

Схема 6
В настоящее время достаточно интенсивно ведутся работы по созданию полусинтетических и синтетиченских селен содержащих лекарственных субстанций с активностью базирующейся на тех свойствах селена, о которых говорилось выше. Но речь идёт о лекарствах, а не о пищевых добавках, поскольку применение соединений селена профилактически чревато непредсказуемостью эффекта. А так как селен в виде селеноцистеина и селенометионина содержится во многих продуктах, где меньше а где больше, то лучше придерживаться лозунга : “полноценное питание вместо лекарств”.
Литература
[1] D. C.Turner, T. C.Stadtman, Arch. Biochem. Biophys., 1973, 154, 366.
[2] J. R.Andreesen, L. G.Ljungdahl, J. Bacteriol., 1973, 116, 867.
[3] L. Flohe, W. A.Gunzler, H. H.Schock, FEBS Lett., 1973, 32, 132.
[4] J. T.Rotruck, A. L.Pope, H. T.Ganther, A. B.Swanson, D. G.Hafeman, W. G.Hoekstra, Science, 1973, 179, 588.
[5] J. E.Cone, R. M.Del Rio, J. N.Davis, T. C.Stadtman, Proc. Natl. Acad. Sci. U. S.A., 1976, 73, 2659.
[6] J. W.Forstrom, J. J.Hachowski, A. L.Tapel, Biochemistry, 1978, 17, 2639.
[7] A. Wendel, B. Kerner, K. Graupe, Hoppr-Seyler’s Z. Physiol. Chem., 1978, 359, 1035.
[8] F. Zinoni, A. Birkmann, T. C.Stadtman, A. Bock, Proc. Natl. Acad. Sci U. S.A., 1986, 83, 4650.
[9] Google: Selenium 7.Analytical Methods (www. atsdr. cdc. gov/toxprofiles/tp92-c7.pdf)
[10] G. N.Schrauzer, J. Nutr., 2000, 130, 1653.
[11] M. Birringer, S. Pilawa, L. Flohe, Nat. Prod. Rep.,2002, 19, 693.
[12] B. Tse Sum Bui, A. Mattioli, D. Florentin, G. Bolbach, A. Marquet, Biochemistry, 2006, 45, 3824.
[13] L. Bendahl, D. Gammelgaard, J. Anal. At. Spectrom.,2004, 19, 143.
[14] Y. Dong, D. Lisk, E. Block, C. Ip, Cancer Research, 2001, 61, 2923.
[15] J. G.Hutton, J. Am. Soc. Agron.,1931, 23, 1076.
[16] K. W.Franke, V. R.Potter, J. Nutr., 1935, 10, 213.
[17] A. A.Nelson, O. G.Fithugh, H. O.Calvery, Cancer Res.,1943, 3, 230.
[18] J. Kohrle, R. Brigelius-Flohe, A. Bock, R. Gartner, O. Meyer, L. Flohe, Biol. Chem.,2000, 381, 849.
[19] M. Maiorino, C. Roche, M. Kiess, K. Koenig, D. Gawlik, M. Matthes, E. Naldini, R. Pierce, L. Flohe, Eur. J.Biochem.,1996, 238, 838.
[20] F. Serras, M. Morey, B. Alsina, J. Baguna, M. Coromonas, Biofactors, 2001, 14, 143.
[21] C. Buettner, J. W.Harney, M. J.Berry, J. Biol. Chem., 1999, 274, 21598.
[22] V. N.Gladychev, in Selenium Its Molecular Biology and Role in Human Health, ed. D. L.Hatfield, Kluwer Academic Publisher, Boston, 2001, p.99.
[23], M. Rother, A. Resch, R. Wilting, A. Bock, Biofactors, 2001,14, 75.
[24] L. H.Fu, X. F.Wang, Y. Eyal, Y. M.Sce, L. J.Donald, K. G.Standing, G. Ben-Hayyim, J. Biol. Chem.,2002, 24, 24.
[25] P. R.Copelan, J. E.Fletcher, B. A.Carlson, D. L.Hatfield, D. M.Driscoll, EMBO Journal, 2000, 19, 306.
[26] A. Mehta, C. M.Rebsch, S. A.Kinzy, J. E.Fletcher, P. R.Copeland, J. Biol. Chem., 2004, 279, 37852.
[27] R. Guigo, V. N.Gladyshev, Science, 2003, 300, 1439.
[28] M. Kotrebai, M. Birringer, J. F.Tyson, E. Block, P. C.Uden, Analyst, 2000, 125, 71
[29] X. J.Cai, E. Block, P. C.Uden, X. Zhang, B. D.Quimby, J. J.Sullivan, J. Agric. Food Chem.,1995, 43, 1754.
[30] X. Guo, L. Wu, Ecotoxicol. Environ. Saf.,1998, 39, 207.
[31] J. M.Stewart, S. N.Nigam, W. B.McConnell, Can. J. Biochem., 1974, 52, 144.
[32] M. Korhola, A. Vainio, K. Edelmann, Ann. Clin. Res., 1986, 18, 65.
[33] C. Casiot, V. Vacchina, H. Chassaigne, J. Szpunar, M. Potin-Gautier, R. Lobinski, mun.,1999, 36, 77.
[34] S. McSheehy, F. Pannier, J. Szpunar, M. Potin-Gautier, R. Lobinski, Analyst., 2002, 127, 223.
[35] F. Bertelsen, G. Gissel-Nielsen, A. Kjaer, T. Skrydstrup, Phytochemistry, 1988, 27, 3743.
[36] T. K.Virupaksha, A. Shrift, Biochim. Biophys. Acta, 1963, 74, 791.
[37] R. A.Zingaro, J. Price, C. R.Benedict, J. Carbohydr. Nucleosides Nucleotides, 1977, 4, 271.
[38] A. Lauchli, Bot. Acta, 1993, 106, 455.
[39] M. Bjornstedt, S. Kumar, A. Holmgren, J. Biol. Chem., 1992, 267, 8030.
[40] I. Schroder, S. Rech, T. Krafft, J. M.Macy, J. Biol. Chem., 1997, 272, 23765.
[41] http://www. vetpharm. unizh. ch/clinitox/
2003
