

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
УДК 543.25:543.8
Тематическое направление: Химия биологически активных соединений.
Реакции папаина с электрогенерированными галогенами и их аналитическое применение
ã +, , *
Кафедра аналитической химии Химического института им. , Казанский государственный университет
, г. Казань, 420008, Республика Татарстан, Россия. Тел.: (843)231-54-91. E-mail: ziyatdinovag@mail.ru
_______________________________________________
*Ведущий направление; +Поддерживающий переписку
Ключевые слова: гальваностатическая кулонометрия, электрогенерированные галогены, папаин, цистеиновые протеиназы.
Резюме
Найдены стехиометрические коэффициенты реакций папаина с электрогенерированными галогенами, которые составили 1:5 и 1:3 для хлора и брома, соответственно. Разработан способ определения папаина методом кулонометрического титрования электрогенерированными хлором и бромом. Обсуждено возможное участие серосодержащих центров молекулы фермента в реакциях с галогенами. Показана возможность определения микрограммовых количеств папина в модельных растворах с величиной относительного стандартного отклонения 0.03-0.08.
Введение
Папаин является одним из протеолитических ферментов – цистеиновых протеиназ, присутствующих в латексе дынного дерева (Carica papaya L.). Название "папаин" было впервые предложено в конце XIX века А. Вюрцем и Е. Бошутом и используется до настоящего времени для обозначения препаратов латекса.
Этот фермент содержится во всех частях растения за исключением корней. Неочищенный или частично очищенный папаин в прошлом был объектом многочисленных исследований. Однако, позднее было установлено, что сухой латекс дынного дерева, кроме папаина, содержит целый набор цистеиновых протеиназ, которые разделяются на три главные группы, собственно папаин, химопапаины А и В и пептидазы А и В. В последние годы папаин и химопапаины были обнаружены также в латексе из вегетативных органов (листьев и черешков) дынного дерева. В 1937 г. Болз и сотр. впервые получили кристаллический папаин, а в 1941 г. Янсен и Болз - кристаллический химопапаин [1].
Три главные группы ферментов латекса папайи различаются по физико-химическим свойствам. Так, они характеризуются разными значениями изоэлектрических точек, которые находятся в зоне рН 9,5, 10,1-10,6 и 11,1 для папаина, химопапаинов и пептидаз, соответственно. Различаются эти группы ферментов и по содержанию титруемых свободных SH-групп. Ферменты первой и третьей групп содержат 1 SH-группу на молекулу белка, ферменты второй группы – 2 [2].
Папаин является наиболее изученным ферментом, хотя на его долю приходится только около 5 % общего содержания растворимого белка латекса дынного дерева. Молекулярная масса фермента, определенная Э. Смитом и др. на основании скорости седиментации и диффузии, равна 20,7 кДа. Молекула папаина состоит из одной полипептидной цепи и содержит 212 аминокислотных остатков. Характерной особенностью аминокислотного состава фермента является отсутствие остатков метионина [3].
Папаин относительно устойчив в нейтральной и слабощелочной среде, но быстро и необратимо теряет активность в области кислых значений рН. По сравнению с другими протеиназами растительного происхождения (например бромелаином и фицином) папаин проявляет значительную устойчивость к действию повышенных температур. Фермент стабилен в лиофилизованном неактивированном состоянии. Потеря активности в водных растворах происходит медленно, но зависит от температуры хранения. При инкубации в присутствии активаторов папаин теряет активность в результате автолиза. Впервые процесс автолиза папаина наблюдали при ионнообменной хроматографии. Медленное протекание процесса автолиза является причиной того, что даже наиболее активные препараты фермента оказываются загрязненными низкомолекулярными продуктами [4].
Молекула папаина обладает высокой устойчивостью к денатурирующим воздействиям. Так, оптическое вращение молекулы папаина существенно не меняется в 6-8 М растворе мочевины или в 70 % метиловом спирте.
Кристаллический папаин обладает, как правило, низкой активностью в отсутствие активаторов. В роли активаторов могут выступать различные низкомолекулярные тиолы, цианиды и ряд других веществ, обладающих свойствами восстановителей, например цистеин, глутатион, тиогликолевая кислота, тиосульфат и боргидрид натрия.
Для получения максимальной активности папаина необходимо одновременное присутствие восстановителя и веществ, способных связывать ионы металлов. Например, используют смесь цистеина (0,005 М) с этилентетрауксусной кислотой (0,002 М) или с версеном (0,001 М). Эта смесь может быть успешно заменена 2,3-меркаптопропанолом, соединением, которое способно одновременно связывать металлы и действовать как восстановитель. Максимальная активность папаина в присутствии активаторов достигается через 5-10 мин [5].
Природа активаторов, а также отношение папаина к окислительным агентам и веществам, специфически блокирующим сульфгидрильные группы белков, позволили давно сделать вывод, что папаин относится к классу цистеиновых протеиназ. Содержание тиоловых групп у неактивного папаина менее 0,5 моля на моль белка, и это же количество определяется при различных способах денатурации фермента. В то же время в активированном папаине присутствует до 1 моля SH-групп на моль белка. Более того, существует корреляция между содержанием тиоловых групп в молекуле папаина и его удельной активностью [6].
Исследования механизма процесса активации и природы неактивной формы папаина позволили И. Клейну и Дж. Киршу получить убедительные доказательства того, что основной компонент неактивного папаина представляет собой смешанный дисульфид белок – цистеин [7].
Папаин используется в пищевой и легкой промышленности для обработки кож, умягчения мяса и осветления напитков [8]. В последние десятилетия папаин нашел широкое применение в составе лекарственных и косметических средств как основной компонент, влияющий на активность ферментов кожи, регулируя ряд биохимических процессов в ней. Косметический эффект папаина заключается в том, что он удаляет отмершие клетки, выравнивает рельеф кожи и стимулирует обмен клеток эпидермиса. Поэтому исследование папаина и разработка способов его определения представляет актуальную задачу.
Цель работы – оценить возможности гальваностатической кулонометрии с электрогенерированными галогенами для количественного определения папаина.
Результаты и дискуссия
Кулонометрическое титрование стандартных растворов электрогенерированными хлором и бромом папаина показало, что реакции этих титрантов протекает быстро и количественно. Результаты титрования позволили установить стехиометрические коэффициенты реакций, которые составили 1:5, 1:3 для хлора и брома, соответственно. Иод не взаимодействует с папаином, вероятно, вследствие стерических препятствий и его малой реакционной способности в этом процессе.
Как уже отмечалось выше, папаин является серосодержащим ферментом класса протеиназ. В структуре папаина присутствуют семь остатков цистеина (Cys). Шесть из семи содержащихся в молекуле остатков Cys участвуют в образовании трех дисульфидных мостиков, которые соединяют остатки полуцистина в положениях 22-63, 56-93 и 153-200 (заштрихованные связи на рисунке). Единственная сульфгидрильная группа принадлежит остатку цистеина, расположенному в положении 25. Этот остаток является частью каталитического центра фермента [24].
Молекула фермента имеет близкую к сферической форму и состоит из двух доменов, обозначенных как L - и R-домены. Домены разделены глубоким "желобом", на поверхности которого находится активный центр фермента. Оба домена содержат практически одинаковое число аминокислотных остатков. Положение одного домена относительно другого поддерживается с помощью трех отрезков полипептидных "хребтов", пересекающих основание "желоба", и множеством водородных связей, а также электростатическими и гидрофобными взаимодействиями боковых групп аминокислотных остатков полипептидной цепи. Центральная область R-домена представляет собой гидрофобное ядро [26, 27].
![]()
![]()
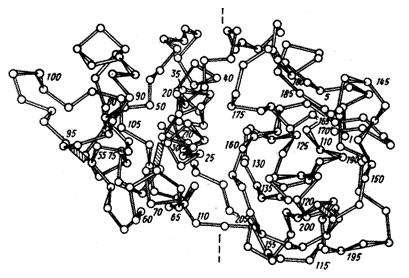
Рисунок. Домены в структуре протеолитического фермента папайи - папаина.
Прерывистыми линиями намечено направление впадины между двумя доменами, в которой связывается субстрат и размещен каталитический центр. Дисульфидные связи заштрихованы.
Исходя из результатов кулонометрического титрования, можно предположить, что в реакцию с электрогенерированными хлором и бромом вступает тиольная группа каталитического центра фермента, расположенного на поверхности "желоба" между двумя доменами. Она окисляется до сульфоновой группы с участием 6 электронов. Электрогенерированный хлор взаимодействует и с дисульфидным фрагментом в положении 22-63. При этом происходит окисление дисульфида до сульфоксида, а электрогенерированный бром не реагирует вследствие стерических затруднений. Два других дисульфидных мостика не вступают в реакцию с электрогенерированными галогенами, поскольку один находится в гидрофобной области молекулы папаина (R-домена), а второй – на значительном расстоянии от активного центра в перпендикулярной плоскости.
Результаты кулонометрического определения папаина в модельных растворах представлены в таблице.
Таблица. Результаты кулонометрического определения папаина в модельных растворах (n=5, Р=0,95).
Титрант | Введено, мкг | Найдено, мкг | Sr |
Cl2 | 2.2 | 2.2±0.2 | 0.08 |
2.6 | 2.6±0.1 | 0.03 | |
3.3 | 3.2±0.2 | 0.05 | |
4.4 | 4.4±0.2 | 0.04 | |
5.5 | 5.5±0.2 | 0.03 | |
Br2 | 2.2 | 2.2±0.1 | 0.04 |
3.3 | 3.2±0.1 | 0.03 | |
4.4 | 4.2±0.2 | 0.03 | |
5.5 | 5.4±0.2 | 0.03 | |
6.6 | 6.5±0.3 | 0.03 |
Экспериментальная часть
Электрогенерацию галогенов осуществляли на потенциостате П-5827 М при постоянной силе тока 5.0 мА из водных 0.2 М KBr в 0.1 М Н2SO4 и из 0.1 М раствора KI в тартратном буферном растворе с рН 3.56. Конечную точку титрования устанавливали амперометрически с двумя поляризованными платиновыми электродами (DЕ=300 мВ). Рабочим электродом служила гладкая платиновая пластинка площадью 1 см2, вспомогательным электродом - платиновая спираль, отделенная полупроницаемой перегородкой от анодного пространства ячейки.
Кулонометрическое определение проводили следующим образом. В ячейку объемом 50.0 мл вводили 20.0 мл фонового раствора, опускали электроды и включали генераторную цепь. По достижении определенного значения индикаторного тока в ячейку вносили аликвоту исследуемого раствора (10-60 мкл) и одновременно включали секундомер. Конечную точку титрования фиксировали по достижению первоначального значения индикаторного тока. При этом выключали секундомер и отключали генераторную цепь.
В работе использовали лиофилизированный папаин от Ferak (Berlin, Germany). Стандартные растворы готовили по точной навеске (около 1.0 г), которую растворяли в 50 мл дистиллированной воды.
Выводы
Найдены стехиометрические коэффициенты реакций папаина с электрогенерированными галогенами, которые составили 1:5 и 1:3 для хлора и брома, соответственно. Электрогенерированный иод не взаимодействует с папаином в условиях гальваностатической кулонометрии. На основе полученных данных обсуждено участие серосодержащих центров молекулы фермента в реакциях с галогенами и разработан способ определения папаина.
Литература
1. Blocklehurst K., Baines B. S., Kierstan M. P.J. Papain and other constituents of Cartica papaya L. In: Topics on enzyme and fermentation biotechnology ( E. Wiseman, E. Horwood). Chichester: Ellis Horwood Ltd, 1981. V.5. P.262-335.
2. Протеолитические ферменты. М.: Наука, 1971. 414с.
3. Glazer A. N., Smith E. L. Papain and other plant sulfhydryl proteolytic enzymes. In: The enzymes. Academic press, 1971. V.3. P.501-546.
4. Drenth J. // Recueil des Travaux Chimiques des Pays-Bas. 1980. V.99. № 6. P.185-190.
5. Polgar L., Csoma C. Characterization of active site of cysteine proteases. In: Molecular and cellular regulation of enzyme activity ( A. Barth, R. L. Schwen, H. Possin). Berlin:Verlag, 1986. P.1-10.
6. Beker E. N., Drenth J. The thiol proteases: structure and mechanism. In: Biological macromolecules and assemblies ( F. A. Iurnak, A. McPherson). N. Y.: J. Wiley&Sons, 1987. V.3. P.314-368.
7. Klein I. B., J. F. Kirsch // Biochem. Biophys. muns. 1969. V.34. № 5. P.575-581.
8. Основы биохимии растений. 5 изд. М., 1971. 460с.
