
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Публикация доступна для обсуждения в рамках функционирования постоянно
действующей интернет-конференции “Бутлеровские чтения”. http:///readings/
УДК: 544.42+577.112,4+577.114.3.
Кинетический анализ ингибирования глутатионом процесса неферментативного гликозилирования in vitro генноинженерного инсулина человека
© ,1 ,1+
,1 ,1
1 и 2*
1 Химико-технологический институт. Уральский федеральный университет им. первого Президента России . Ул. Мира, 19.
г. Екатеринбург, 620002. Россия. Тел.: (343) 375-47-94. E-mail: esavateeva@gmail.com
2 Институт иммунологии и физиологии УрО РАН. Ул. Первомайская, 106.
г. Екатеринбург, 620049. Россия. Тел.: (343) 374-00-70.
________________________________________________
*Ведущий направление; +Поддерживающий переписку
Ключевые слова: кинетика, неферментативное гликозилирование белков, инсулин, глутатион.
Аннотация
Рассмотрены кинетические особенности ингибирования восстановленным глутатионом процесса неферментативного гликозилирования инсулина in vitro на его начальном этапе, где он состоит из двух последовательных стадий s=1,2 и протекает в квазиравновесном режиме по стадии 1. Установлено, что добавки глутатиона не нарушают двухстадийный механизм данного процесса, но снижают его темп и выход конечного продукта – фруктозамина – на стадии 2 за счёт связывания глутатионом (Y) инсулина (I) как реагента стадии 1 в химическое соединение типа IαY по маршрутной реакции р=3: αI + Y = IαY, где α – модуль стехиометрического коэффициента инсулина. Термодинамические расчёты равновесных концентраций всех компонентов в рабочих растворах по их начальным концентрациям с использованием ранее найденных констант равновесия стадии 1 и 2 показывают, что маршрутная реакция 3 реализуется при α=4 и имеет константу равновесия ![]() =733 (Т=277К,
=733 (Т=277К, ![]() = 1 моль/м3).
= 1 моль/м3).
Введение
Неферментативное гликозилирование белков (НГБ) – взаимодействие аминогрупп белков и карбонильных групп редуцирующих сахаров – является одной из значимых посттрансляционных модификаций белков в организме. Присоединение остатков моносахаридов нарушает ионные взаимодействия в белковой молекуле, изменяет ее конформацию, растворимость, а вследствие этого – функциональные свойства и чувствительность к действию протеаз [1,2,3]. Содержание в крови и тканях неферментативно гликозилированных белков многократно повышается при сахарном диабете, характеризующемся длительной и выраженной гипергликемией [1,2].
Одним из белков, подвергающихся гликозилированию при сахарном диабете, является гормон инсулин. В литературе имеется подробная характеристика структуры гликозилированного инсулина животных и человека [4-6]. Большинство исследователей сходятся во мнении, что присоединение глюкозы происходит прежде всего по аминогруппе концевого фенилаланина в В-цепи инсулина. Наши исследования показывают, что процесс гликозилирования инсулина на начальном этапе его развития отчетливо проявляется в виде совокупности двух стадий, имеющих двусторонний характер, причем течение процесса происходит в квазиравновесном режиме по первой стадии [7].
Гликозилирование приводит к частичной утрате биологической активности гормона. Согласно данным [8], гликозилирование инсулина и его предшественника проинсулина может происходить еще в β-клетках, которые затем секретируют в кровь неполноценный гормон, что усугубляет нарушения обмена веществ при сахарном диабете. В связи с изложенным, представляет интерес выяснить, можно ли повлиять на процесс гликозилирования инсулина с помощью каких-либо фармакологических агентов.
В качестве возможного ингибитора неферментативного гликозилирования инсулина привлекает внимание восстановленный глутатион (G-SH). Это соединение является основным внутриклеточным тиольным антиоксидантом, G-SH обнаруживается в клетках в концентрациях 0,5 – 10,0 ммоль/л [1]. В литературе имеется немало сведений о способности гетерофункциональных природных соединений, содержащих амино - и тиольную группу, взаимодействовать с карбонильными соединениями, тем самым конкурировать с белком и ингибировать НГБ [9-13]. Однако нет единого мнения о механизме такой активности. Рассматривается возможность образования аддуктов глутатиона, цистеина с карбонильными соединениями по амино - и тиольной группам с образованием шиффовых оснований и полутиоацеталей, соответственно, а также по обеим группам с образованием циклических производных [9-13]. Возможность ингибирования неферментативного гликозилирования инсулина при помощи G-SH, а также механизм такого действия ранее не подвергались специальному исследованию.
Экспериментальная часть
Исследование ингибирующего действия глутатиона на процесс гликозилирования инсулина проводилось на базе водных растворов, которые по начальному составу, если исключить добавки ингибиторов, были близки к водным растворам, используемым ранее при изучении кинетических закономерностей процесса гликозилирования инсулина in vitro без каких-либо добавок [7]. В тех и других растворах роль исходных веществ играли:
Ø инсулин человеческий генноинженерный (символ I) под торговым названием «Биосулин Р» производства ЗАО «Фармстандарт»;
Ø D-глюкоза одноводная (символ G) квалификации х. ч. производства «НПО ЭКРОС»;
Ø вода дистиллированная (символ H2O);
Ø глутатион восстановленный, производства «Reanal» (символ Y)
При постановке опытов рабочие растворы готовились при комнатной температуре смешением исходных веществ в определенных соотношениях и добавлением к смеси определённого количества ингибитора. Объём каждого раствора составлял около 10 см3. Во всех растворах начальные концентрации воды ![]() , инсулина и глюкозы были одинаковы (
, инсулина и глюкозы были одинаковы (![]() = 55400 моль/м3, = 0.875 моль/м3, = 50.0 моль/м3), тогда как начальные концентрации глутатиона – разные ( = 0; 0.05; 0.50; 5.0; 50.0 моль/м3). Приготовленные растворы тотчас инкубировались при заданной температуре Т и давлении Р (Т = 277 К, Р = 1 атм) в течение одного и того же интервала времени ( = 11 недель= 1848 часа).
= 55400 моль/м3, = 0.875 моль/м3, = 50.0 моль/м3), тогда как начальные концентрации глутатиона – разные ( = 0; 0.05; 0.50; 5.0; 50.0 моль/м3). Приготовленные растворы тотчас инкубировались при заданной температуре Т и давлении Р (Т = 277 К, Р = 1 атм) в течение одного и того же интервала времени ( = 11 недель= 1848 часа).
Контроль за ходом процесса гликозилирования осуществлялся путем отбора проб из рабочих растворов в определенные моменты времени t (t = 1; 2,5; 5; 11 недель) и определения в них концентрации ![]() фруктозамина колориметрическим методом по реакции с тиобарбитуровой кислотой [14]. Оптическая плотность стандартных образцов и препарированных проб измерялась с помощью фотоэлектроколориметра КФК-3-01-ЗОМЗ. Каждое найденное значение является средним из трех параллельных определений.
фруктозамина колориметрическим методом по реакции с тиобарбитуровой кислотой [14]. Оптическая плотность стандартных образцов и препарированных проб измерялась с помощью фотоэлектроколориметра КФК-3-01-ЗОМЗ. Каждое найденное значение является средним из трех параллельных определений.
Результаты и их обсуждение
На рис. 1 представлены экспериментальные данные по изменению концентрации ![]() в рабочих растворах с течением времени t при одинаковых начальных концентрациях
в рабочих растворах с течением времени t при одинаковых начальных концентрациях ![]() , , , но разных концентрациях . Они свидетельствуют, что кинетические кривые при всех [0;50.0] моль/м3, обладают общим характерным признаком, а именно: каждая кривая ведёт себя как монотонно возрастающая функция, стремящаяся к некоторому пределу
, , , но разных концентрациях . Они свидетельствуют, что кинетические кривые при всех [0;50.0] моль/м3, обладают общим характерным признаком, а именно: каждая кривая ведёт себя как монотонно возрастающая функция, стремящаяся к некоторому пределу ![]() (
(![]() < ), когда t →
< ), когда t → ![]() (
(![]() = ∞ условно). Из рис. 1 видно также, что по мере повышения предел
= ∞ условно). Из рис. 1 видно также, что по мере повышения предел ![]() неуклонно снижается.
неуклонно снижается.
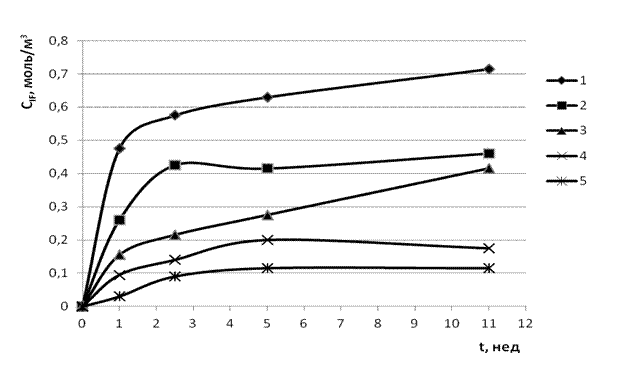
Рис. 1. Кинетические кривые ![]() (n = 1, 2, 3, 4, 5) при
(n = 1, 2, 3, 4, 5) при ![]() = 55400 моль/м3,
= 55400 моль/м3, ![]() = 0.872 моль/м3,
= 0.872 моль/м3, ![]() =50.0 моль/м3 и разных : 1 – 0; 2 – 0.05; 3 – 0.50; 4 – 5.0; 5 – 50.0. Температура Т = 277 К.
=50.0 моль/м3 и разных : 1 – 0; 2 – 0.05; 3 – 0.50; 4 – 5.0; 5 – 50.0. Температура Т = 277 К.
Как было показано ранее [7], процесс неферментативного гликозилирования инсулина in vitro без каких-либо добавок глутатиона на его начальном этапе состоит из двух последовательных стадий s (s = 1 и 2), первая из которых представляет собой стадию образования промежуточного соединения – основания Шиффа (символ IG) в результате взаимодействия инсулина I с глюкозой G, а вторая – стадию превращения IG во фруктозамин (символ IF):
1. I + G = IG + H2O, (1)
2. IG = IF, (2)
Обе стадии имеют двусторонний характер, но стадия 1 существенно превосходит стадию 2 по быстроте достижения равновесия, обеспечивая тем самым течение процесса гликозилирования на его начальном этапе в квазиравновесном режиме по стадии 1. Что касается кинетики данного процесса, то она определяется интегральным уравнением вида
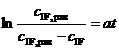 , (3)
, (3)
где величины ![]() и
и ![]() выступают в качестве параметров, зависящих от константы равновесия
выступают в качестве параметров, зависящих от константы равновесия ![]() стадии 1, констант скоростей
стадии 1, констант скоростей ![]() (Т) и
(Т) и ![]() (Т) прямой и обратной элементарных реакций стадии 2, а также от начальных концентраций
(Т) прямой и обратной элементарных реакций стадии 2, а также от начальных концентраций ![]() , , .
, , .
Мы полагаем, что добавки глутатиона не нарушают двухстадийный механизм процесса гликозилирования инсулина на его начальном этапе и, следовательно, не изменяют логарифмический характер интегрального кинетического уравнения (3), но они оказывают ингибирующее действие на темп и степень продвижения процесса через параметры ![]() и
и ![]() в уравнении (3). Это возможно, если только глутатион частично связывает собой один или оба реагента стадии 1 в химические соединения типа IαY (α=1,2,…) и GβY (β=1,2,…) по двум новым независимым и двусторонним реакциям 3 и 4:
в уравнении (3). Это возможно, если только глутатион частично связывает собой один или оба реагента стадии 1 в химические соединения типа IαY (α=1,2,…) и GβY (β=1,2,…) по двум новым независимым и двусторонним реакциям 3 и 4:
3. αI + Y = IαY, (4)
4. βG + Y = GβY, (5)
где целые и положительные числа α и β играют роль модулей стехиометрических коэффициентов инсулина I и глюкозы G. При α>2 и β>2 обе реакции 3 и 4, скорее всего, являются представителями так называемых маршрутных реакций [15], имеющих многостадийный характер и обозначаемых обычно символом р (от англ. path – тропа, маршрут).
Если, кроме того, допустить, что реакции 3 и 4, подобно стадии 1, многократно превосходят стадию 2 по быстроте достижения своих равновесных состояний, то можно сказать, что процесс гликозилирования инсулина в присутствии глутатиона протекает в квазиравновесном режиме не только по стадии 1, но и по реакциям 3 и 4.
Для проверки и подтверждения данной гипотезы мы выполнили обработку всех кинетических кривых , изображённых на рис. 1, по интегральному уравнению (3), которое в координатах t и ![]() является прямой линией с угловым коэффициентом
является прямой линией с угловым коэффициентом ![]() . Выбор положения предельной (равновесной) точки
. Выбор положения предельной (равновесной) точки ![]() на каждой кинетической кривой проводился, как и в случае работы [7], при помощи компьютерной программы Excel с таким расчётом, чтобы обеспечить наименьшие отклонения остальных точек относительно прямой линии.
на каждой кинетической кривой проводился, как и в случае работы [7], при помощи компьютерной программы Excel с таким расчётом, чтобы обеспечить наименьшие отклонения остальных точек относительно прямой линии.
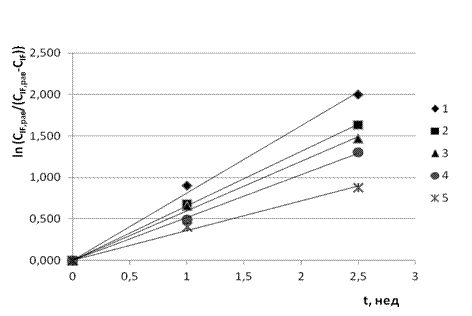
Рис. 2. Линейные образы кинетических кривых ![]() (n = 1, 2, 3, 4, 5)
(n = 1, 2, 3, 4, 5)
в координатах t и 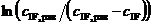 при следующем выборе
при следующем выборе ![]() , моль/м3: 1 – 0.637; 2 – 0.528; 3 – 0.342; 4 – 0.200; 5 – 0.120.
, моль/м3: 1 – 0.637; 2 – 0.528; 3 – 0.342; 4 – 0.200; 5 – 0.120.
Полученные результаты представлены на рис. 2, из которого видно, что уравнение (3) действительно выполняется при всех [0;50,0] моль/м3. Данные табл. 1 свидетельствуют, что параметр ![]() , подобно параметру
, подобно параметру ![]() , неуклонно уменьшается с увеличением .
, неуклонно уменьшается с увеличением .
Табл. 1. Влияние начальной концентрации на параметры ![]() и
и ![]() (в уравнении (9)) по данным рис. 1 и 2.
(в уравнении (9)) по данным рис. 1 и 2.
, моль/м3 | 0 | 0.05 | 0.50 | 5.0 | 50.0 |
| 0.637 | 0.528 | 0.343 | 0.200 | 0.120 |
| 6.616 | 3.911 | 3.548 | 3.168 | 2.130 |
Графики функций ![]() ( ) и
( ) и ![]() ( ), изображённые на рис. 3, отчётливо показывают, что снижение наибольшей функции происходит во много раз быстрее при низких значениях аргумента , чем при высоких.
( ), изображённые на рис. 3, отчётливо показывают, что снижение наибольшей функции происходит во много раз быстрее при низких значениях аргумента , чем при высоких.
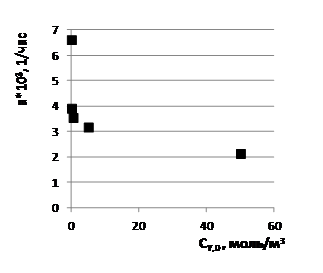
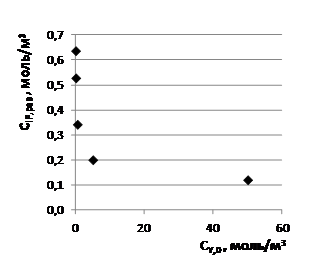
Рис. 3. Графики зависимостей ![]() ( )(1) и
( )(1) и ![]() ( )(2) по данным табл. 1.
( )(2) по данным табл. 1.
Принимая во внимание, что существенные изменения параметров ![]() и
и ![]() наблюдаются уже при небольших значениях ≤1 моль/м3 (см. рис. 3) на фоне сопоставимых значений
наблюдаются уже при небольших значениях ≤1 моль/м3 (см. рис. 3) на фоне сопоставимых значений ![]() = 0,872 моль/м3 и гораздо более высоких значений
= 0,872 моль/м3 и гораздо более высоких значений ![]() =50 моль/м3, естественно допустить, что ингибирующее действие глутатиона на параметры
=50 моль/м3, естественно допустить, что ингибирующее действие глутатиона на параметры ![]() и
и ![]() обусловлено его способностью связывать собой преимущественно инсулин I по реакции 3. Чтобы убедиться в этом, необходимо провести термодинамический анализ текущих и равновесных состояний равновесных растворов при условии, что химическое превращение в них включает три независимых реакции r (r = 1, 2, 3): две стадии s (s = 1,2) и одну маршрутную реакцию р (р = 3) [16].
обусловлено его способностью связывать собой преимущественно инсулин I по реакции 3. Чтобы убедиться в этом, необходимо провести термодинамический анализ текущих и равновесных состояний равновесных растворов при условии, что химическое превращение в них включает три независимых реакции r (r = 1, 2, 3): две стадии s (s = 1,2) и одну маршрутную реакцию р (р = 3) [16].
Известно, что в любом растворе концентрация каждого компонента k, выраженная в шкале объёмных плотностей чисел молей, ![]() -шкале, являются линейными функциями независимых
-шкале, являются линейными функциями независимых ![]() – переменных
– переменных ![]() , имеющих смысл объёмных плотностей глубин реакций r:
, имеющих смысл объёмных плотностей глубин реакций r:
![]() (8)
(8)
где ![]() – начальная концентрация компонента k,
– начальная концентрация компонента k, ![]() – стехиометрический коэффициент того же компонента k в реакции r (
– стехиометрический коэффициент того же компонента k в реакции r (![]() <0, >0 или = 0, если компонент k является соответственно реагентом, продуктом или неучастником реакции r). Конкретизация (8) с привлечением стехиометрических уравнений (1)-(3) даёт
<0, >0 или = 0, если компонент k является соответственно реагентом, продуктом или неучастником реакции r). Конкретизация (8) с привлечением стехиометрических уравнений (1)-(3) даёт
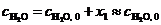 , (9)
, (9)
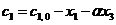 , (10)
, (10)
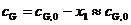 , (11)
, (11)
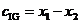 , (12)
, (12)
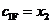 , (13)
, (13)
![]() (14)
(14)
![]() (15)
(15)
Где дополнительно учтено, что компоненты IG, IF, IαY в начальный момент времени ![]() отсутствуют (
отсутствуют (![]() =0,
=0, ![]() =0,
=0, ![]() =0), и принято во внимание, что слагаемое
=0), и принято во внимание, что слагаемое ![]() в уравнениях (9) и (11) пренебрежимо мало по сравнению с
в уравнениях (9) и (11) пренебрежимо мало по сравнению с ![]() и
и ![]() (
(![]() <<
<< ![]() и
и ![]() <<
<< ![]() ), так как согласно уравнению (10)
), так как согласно уравнению (10) ![]() не может превышать
не может превышать ![]() (
(![]() ≤
≤ ![]() ), а во всех рабочих растворах
), а во всех рабочих растворах ![]() и
и ![]() .
.
С термодинамической точки зрения рабочие растворы близки к модели разбавленных идеальных растворов, где коэффициенты активностей ![]() компонентов k в
компонентов k в ![]() -шкале равны единице (
-шкале равны единице (![]() = 1 для всех k). Это позволяет использовать следующую общую формулу выражения законов равенства реакций r:
= 1 для всех k). Это позволяет использовать следующую общую формулу выражения законов равенства реакций r:
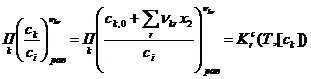 , (16)
, (16)
где [![]() ] – единица измерения
] – единица измерения ![]() ,
, ![]() – константа равновесия реакции r в
– константа равновесия реакции r в ![]() -шкале, зависящая только от T и [cк] (её зависимость от давления является весьма слабой из-за малой сжимаемости жидкой среды). Опираясь на (16) и привлекая стехиометрические уравнения (1)-(3) вместе с балансовыми уравнениями (9)-(15), получаем конкретные формы выражения законов равновесия стадий s и реакции р (s=1, 2; p=3):
-шкале, зависящая только от T и [cк] (её зависимость от давления является весьма слабой из-за малой сжимаемости жидкой среды). Опираясь на (16) и привлекая стехиометрические уравнения (1)-(3) вместе с балансовыми уравнениями (9)-(15), получаем конкретные формы выражения законов равновесия стадий s и реакции р (s=1, 2; p=3):
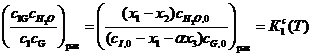 (17)
(17)
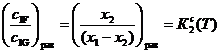 , (18)
, (18)
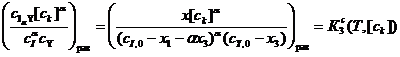 (19)
(19)
Система из двух уравнений (17) и (18) при заданных ![]() ,
, ![]() ,
, ![]() ,
, ![]() позволяет сначала рассчитать неизвестные переменные
позволяет сначала рассчитать неизвестные переменные ![]() и
и ![]() , а затем (привлекая дополнительное балансовое уравнение (10) и (14) неизвестные переменные
, а затем (привлекая дополнительное балансовое уравнение (10) и (14) неизвестные переменные ![]() и
и ![]() , если переменная
, если переменная ![]() известна.
известна.
Мы провели такой расчёт, используя найденные ранее [7] числовые значения констант равновесия стадий 1 и 2 при Т = 277 К: ![]() = 134,2;
= 134,2; ![]() = 21,0, а также полученные в настоящей работе числовые значения
= 21,0, а также полученные в настоящей работе числовые значения ![]() при постоянных
при постоянных ![]() = 55400 моль/м3 и
= 55400 моль/м3 и ![]() =50 моль/м3, но переменных
=50 моль/м3, но переменных ![]() (см. табл. 1), имея в виду, что
(см. табл. 1), имея в виду, что ![]() =
=![]() , как это следует из балансового уравнения (13). Результаты расчёта в форме функций
, как это следует из балансового уравнения (13). Результаты расчёта в форме функций ![]() ,
, 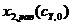 ,
, 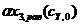 ,
, 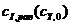 и
и 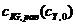 сведены в табл. 2. Их графические изображения для наглядности даны на рис. 4.
сведены в табл. 2. Их графические изображения для наглядности даны на рис. 4.
Табл. 2. Результаты расчёта функций 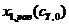 , , , и на основе законов равновесия (17) и (18) с учётом балансовых уравнений (10), (12) и (13).
, , , и на основе законов равновесия (17) и (18) с учётом балансовых уравнений (10), (12) и (13).
| 0 | 0.05 | 0.50 | 5.0 | 50.0 |
| 0.668 | 0.553 | 0.359 | 0.210 | 0.126 |
| 0.637 | 0.528 | 0.343 | 0.200 | 0.120 |
| 0 | 0.114 | 0.381 | 0.587 | 0.702 |
| 0.250 | 0.208 | 0.135 | 0.079 | 0.047 |
| 0.031 | 0.025 | 0.016 | 0.010 | 0.006 |
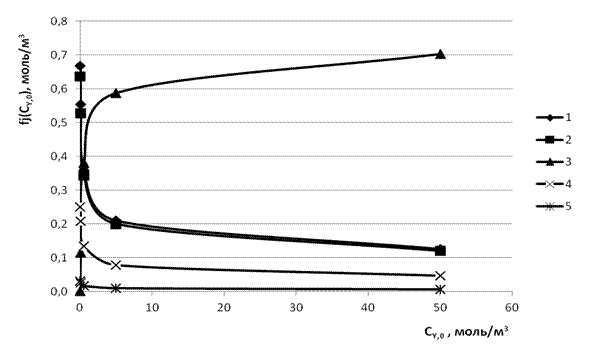
Рис. 4. Графики функций fj(![]() )(j = 1,2,3,4,5): 1-
)(j = 1,2,3,4,5): 1-![]() , 2-
, 2-![]() , 3-
, 3-![]() , 4-
, 4-![]() , 5-
, 5-![]() по данным табл. 2.
по данным табл. 2.
Располагая зависимостью ![]() (см. табл.2), нетрудно получить расчётным путём серию зависимостей типа
(см. табл.2), нетрудно получить расчётным путём серию зависимостей типа ![]() с разными значениями α (α=1, 2, 3 и т. д.), а также серию зависимостей типа
с разными значениями α (α=1, 2, 3 и т. д.), а также серию зависимостей типа ![]() с теми же α, подключив к расчёту в последнем случае балансовое уравнение (14). Обе серии размещены в табл. 3. Этих сведений
с теми же α, подключив к расчёту в последнем случае балансовое уравнение (14). Обе серии размещены в табл. 3. Этих сведений
Табл. 3. Результаты расчёта функций ![]() и
и ![]() при разных α с учётом балансового уравнения (14)
при разных α с учётом балансового уравнения (14)
α |
| 0 | 0.05 | 0.50 | 5.0 | 50.0 |
| 0 | 0.114 | 0.381 | 0.587 | 0.702 | |
1 |
| 0 | 0.114 | 0.381 | 0.587 | 0.702 |
| 0 | -0.064 | 0.119 | 4.413 | 49.298 | |
2 |
| -0.021 | 0.057 | 0.190 | 0.293 | 0.351 |
| 0.021 | -0.007 | 0.310 | 4.707 | 49.649 | |
3 |
| -0.014 | 0.038 | 0.127 | 0.196 | 0.234 |
| 0.014 | 0.012 | 0.373 | 4.804 | 49.766 | |
4 |
| -0.011 | 0.029 | 0.095 | 0.147 | 0.176 |
| 0.011 | 0.021 | 0.405 | 4.853 | 49.824 | |
5 |
| -0.009 | 0.023 | 0.076 | 0.117 | 0.140 |
| 0.009 | 0.027 | 0.424 | 4.883 | 49.860 |
вместе со сведениями по зависимости в табл. 1 достаточно, чтобы с помощью уравнения (19) рассчитать числовые значения константы равновесия ![]() при разных
при разных ![]() и α для решения вопроса о том, при каком α данная константа действительно ведёт себя как сохраняемая величина с изменением
и α для решения вопроса о том, при каком α данная константа действительно ведёт себя как сохраняемая величина с изменением ![]() . Полученные здесь результаты расчёта представлены в табл. 4, из которой следует, что требование сохраняемости величины
. Полученные здесь результаты расчёта представлены в табл. 4, из которой следует, что требование сохраняемости величины ![]() лучше всего выполняется при α=4, поскольку в этом случае с изменением,
лучше всего выполняется при α=4, поскольку в этом случае с изменением, ![]() величина
величина ![]() представляет сравнительно небольшие отклонения от её среднего значения:
представляет сравнительно небольшие отклонения от её среднего значения: ![]() =733 ±29.
=733 ±29.
Табл. 4. Результаты расчёта ![]() на основе закона равновесия (19) при разных α и
на основе закона равновесия (19) при разных α и ![]() . Температура Т=277 К, [ck] = 1 моль/м3.
. Температура Т=277 К, [ck] = 1 моль/м3.
| |||||
α |
| Среднее | |||
0.05 | 0.50 | 5.0 | 50.0 | ||
3 | 356 | 139 | 84 | 45 | 156±100 |
4 | 717 | 712 | 792 | 712 | 733±29 |
5 | 2184 | 4037 | 8012 | 12073 | 6576±3466 |
Этот результат является довольно неожиданным из-за большого значения α=4, поэтому он нуждается в дополнительном обсуждении с точки зрения механизма взаимодействия инсулина с глутатионом. Вероятно, глутатион за счет имеющихся в его составе полярных функциональных групп способен нековалентно взаимодействовать с инсулином, за счет чего изменяется конформация гормона. Также известно, что в растворе инсулин образует агрегаты – тетра и гексамеры. Диссоциация данных комплексов на мономеры может увеличивать доступность аминогрупп для гликозилирования, поэтому замедление диссоциации в присутствии глутатиона способно ингибировать процесс.
Выводы:
1. Установлено, на начальном этапе процесса гликозилирования инсулина in vitro, где он выступает в виде совокупности двух последовательных стадий s=1,2, участниками которых являются инсулин I, глюкоза G, основание Шиффа IG, вода H2O и фруктозамин IF:
1. I + G = IG + H2O,
2. IG = IF,
добавки глутатиона Y не изменяют двухстадийный механизм данного процесса, протекающего в квазиравновесном режиме по стадии 1, но они существенно снижают его темп и степень продвижения за счёт связывания инсулина глутатионом в химическое соединение типа IαY по маршрутной реакции р=3:
3. αI + Y = IαY,
где α – модуль стехиометрического коэффициента инсулина.
2. Показано, что в присутствии глутатиона течение процесса гликозилирования инсулина на его начальном этапе происходит в квазиравновесном режиме не только по стадии 1, но и по маршрутной реакции 3, поэтому его ход во времени t по прежнему подчиняется интегральному кинетическому уравнению стадии 2, выраженному в логарифмической форме:  , где параметры
, где параметры ![]() и
и ![]() зависят от начальной концентрации глутатиона
зависят от начальной концентрации глутатиона ![]() .
.
3. Выполнены термодинамические расчёты равновесных концентраций всех компонентов рабочих растворов по их начальным концентрациям и ранее найденным константам равновесия стадии 1 и 2, что позволило определить для маршрутной реакции 3 величину α=4 и константу равновесия 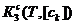 =733 (Т=277К,
=733 (Т=277К, ![]() = 1 моль/м3).
= 1 моль/м3).
Литература
[1] , , Черешнев гликозилирование белков: химия, патофизиология, перспективы коррекции. Вопр. биол., мед. и фарм. химии. 2010. №1. C.3-15.
[2] , , Ромашкан протеины: биохимия неферментативного гликозилирования протеинов. Новосибирск: Наука. Сиб. отделение. 1989. 258с.
[3] A. M. McKillop, M. H. Mooney, P. Harriott, P. R. Flatt, F. P.M. O’Harte. Еvaluation of glycated insulin in diabetic animals using immunocytochemistry and radioimmunoassay. Biochemical and Biophysical Research Communications. 2011. Vol.286. Р.524-528.
[4] S. W. Vetter, V. S.K. Indurthi. Moderate glycation of serum albumin affects folding, stability, and ligand binding. Clinica Chimica Acta. 2011. Vol.12. Р.2105-2116.
[5] A. C. Boyd, Y. H.A. Abdel-Wahab, A. M. McKillop, H. McNulty, C. R. Barnett, F. P.M. O'Harte, P. R. Flatt. Impaired ability of glycated insulin to regulate plasma glucose and stimulate glucose transport and metabolism in mouse abdominal muscle. Biochimica et Biophysica Acta. 2000. Vol.1523. Р.128-134.
[6] F. P.M. O’Harte, A. C. Boyda, A. M. McKillop, Y. H.A. Abdel-Wahab, H. McNulty, C. R. Barnett, J. M. Conlon, P. Højrupb, P. R. Flatt. Structure, antihyperglycemic activity and cellular actions of a novel diglycated human insulin. Peptides. 2000. Vol.21. Р.1519-1526.
[7] , , Черешнев анализ реакции неферментативного гликозилирования in vitro генноинженерного инсулина человека // Бутлеровские сообщения. 2012.Т.30, №6. С.94 – 102.
[8] A. M. McKillop, A. Meade, P. R. Flatt, F. P.M. O’Harte. Evaluation of the site(s) of glycation in human proinsulin by ion-trap LCO electrospray ionisation mass spectrometry. Regulatory peptides. 2003. Vol.113. Р.1-8.
[9] J. M. Acimovic, B. D. Stanimirovic, N. Todorovic, V. B. Jovanovic, L. M. Mandic. Influence of the microenvironment of thiol groups in low molecular mass thiols and serum albumin on the reaction with methylglyoxal. Chemico-Biological Interactions. 2010. Vоl.188. P. 21–30.
[10] L. Wlodek, J. Chubak. Formation of 2-methyl-2,4-thiazolidinedicarbonxylic acid from L-cysteine in rat tissues. Acta Biochimika Polonika. 1984. Vol.31. No 3. P. 279-288.
[11] L. Wlodek. The reaction of sulfhydryl groups with carbonyl compounds. Acta Biochimika Polonika. 1988. Vol.35. No 4. P. 307-317.
[12] M. D. Linetsky, E. V. Shipova, O. O. Argirov. Influence of glutathione fructosylation on its properties. Archives of Biochemistry and Biophysics. 2006. Vol.449. P. 34–46.
[13] M. D. Linetsky, E. V. Shipova, R. D. Legrand, O. O. Argirov. Glucose-derived Amadori compounds of glutathione. Biochimica et Biophysica Acta. 2005. Vol.1724. Р.181– 193.
[14] , Городецкий метод определения неферментативно гликозилированного альбумина и гемоглобина. Лаб. дело. 1990. №5. С.15-18.
[15] , К оценке числа внутренних стационарных состояний многомаршрутных каталитических реакций. Бутлеровские сообщения. 2009. Т.16. №3. С.30-34.
[16] , Лундин необратимых физико-химических процессов. М.: Химия, 1984. 336 с.
