
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Взаимоотношения в системе «хищник–жертва»
Хищничество, т. е. поглощение пищи, в самом широком смысле представляет собой главную силу, обеспечивающую потоки энергии и веществ в экосистемах. Эффективность, с которой хищники находят и схватывают свою добычу, определяет скорость потока энергии от одного трофического уровня к другому. Хищничество является одним из строительных блоков, образующих структуру сообщества и обеспечивающих его стабильность. Хищник и жертва воздействуют друг на друга, но их взаимоотношения, благоприятные для хищника, наносят вред жертве. Примеров того, как хищники подавляют популяции жертвы, снижая их численность до уровня, не достигающего ёмкости среды, очень много и их можно найти в любой группе организмов.
Особенности взаимоотношений между популяциями хищника и жертвы удобно представить графически, путём сопоставления скорости роста численности жертвы в зависимости от её плотности и доли жертвы, выедаемой хищниками, также в зависимости от плотности жертвы (рис. 22) [10]. Одна кривая на рис. 22 отражает скорость прироста новых особей в популяции жертвы (сверх того их числа, которое гибнет не от хищников, а по другим причинам) в результате либо размножения, либо иммиграции, что объединяется под общим термином прирост, в зависимости от плотности популяции жертвы (Pж = Nж / S — численность популяции, приходящаяся на единицу занимаемой территории). Другая кривая определяет скорость выедания жертвы хищниками.
Кривая пополнения популяции жертвы снижается с увеличением плотности жертвы вследствие увеличения внутривидовой конкуренции за ресурсы и падает до нуля, когда плотность популяции жертвы достигает уровня Pжэё, соответствующего экологической ёмкости среды (рис. 22).
В отсутствие хищников численность популяции жертвы регулируется именно экологической ёмкостью среды, определяемой равновесием между их биотическим потенциалом к размножению и ограниченностью ресурсов среды.
В присутствии хищника ситуация существенно изменяется. Пусть начальная плотность популяции жертвы равна pж1 (рис. 22). В данном случае прирост (ДNж) превышает выедание жертвы хищником (ДNх1) и, следовательно, численность популяции жертвы увеличивается, соответственно растёт плотность популяции. Таким образом, система «хищник — жертва» движется слева направо к точке a на рис. 21. Если же начальная плотность популяции жертвы равна pж2, то в этом случае уже выедание превышает прирост и численность популяции жертвы уменьшается, следовательно, система «хищник — жертва» движется справа налево снова к точке a. Таким образом, точка a на рис. 22 оказывается точкой устойчивого равновесия.

Рис. 22. Кривые пополнения и выедания популяции жертвы
в системе «хищник–жертва»
Аналогичным образом можно рассмотреть поведение системы «хищник — жертва» в окрестностях точек пересечения b и с на рис. 22. В результате несложно установить, что точка с также является точкой устойчивого равновесия, а точка b — точкой неустойчивого равновесия (система «хищник — жертва» движется от точки b).
Таким образом, система «хищник — жертва» может находиться в одной из двух возможных точек динамического равновесия численности жертвы или колеблется вблизи этих точек. Точка равновесия, достигаемая системой «хищник — жертва», устанавливается в зависимости от соотношения эффективности хищника как охотника и потенциала роста численности популяции жертвы. Эффективные хищники держат популяцию жертвы на низком уровне плотности (в точке a) вдали от экологической ёмкости среды. Неэффективные хищники держат популяцию жертвы в точке с, уничтожая наиболее уязвимую часть популяции жертвы, незначительно снижая её численность до уровня, близкого к экологической ёмкости среды (Pжэё – точка d на рис. 22).
Графические кривые пополнения и выедания популяции жертвы в общем случае могут иметь лишь одну точку пересечения: либо в точке a, либо в точке c. В таких случаях хищник считается либо абсолютно эффективным (рис. 23,а), либо абсолютно неэффективным (рис. 23,б).
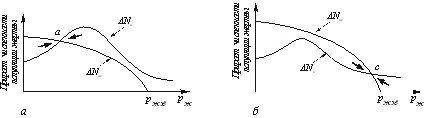
Рис. 23. Кривые пополнения и выедания популяции жертвы для
абсолютно эффективных (а) и абсолютно неэффективных (б) хищников
Воздействие хищника на популяцию жертвы в точке a, довольно ненадёжно. Если сильные морозы или какая-нибудь болезнь понизят численность популяции хищника и удержат её на низком уровне в течение достаточно длительного времени, чтобы популяция жертвы успела перешагнуть за точку b, то рост численности жертвы будет продолжаться независимо от того, произойдёт ли восстановление популяции хищника или нет.
Применительно к сельскому хозяйству это означает, что какой-нибудь вредитель, численность которого обычно удерживается на безопасном для посевов уровне хищниками и паразитами, может внезапно дать угрожающую вспышку роста своей численности. После такой вспышки хищники уже не в состоянии регулировать популяцию вредителя до тех пор, пока какое-нибудь неординарное событие в среде не понизит численность этой популяции до уровня, лежащего левее точки b, т. е. не вернёт её в сферу регулирующего влияния хищника.
Неэффективность хищников повышает стабильность системы хищник — жертва в окрестности точки равновесия c, и регуляция численности популяции при этом целиком зависит от реакции жертвы на пищевые ресурсы и, следовательно, от плотности жертвы. Вблизи точки а эффективность хищника ограничивается тем, что особей жертвы становится меньше, они находятся на больших расстояниях друг от друга и бульшая их часть имеет доступ к надёжным убежищам.
В связи с рассмотренными особенностями хищничества многие экологи считают, что при биологической борьбе с вредителями эффективность используемых паразитов и хищников можно повысить, снабжая их безвредной альтернативной жертвой.
Хищник, выедающий свою жертву так интенсивно, что её численность становится очень низкой, можно сказать, вырывает кусок из собственной пасти. Хищникам легче ловить добычу и, следовательно, увеличивать свою продуктивность, когда их жертвы многочисленны. Небольшая же популяция жертвы может обеспечить существование лишь небольшого числа хищников. Общая скорость пополнения каждой популяции жертвы достигает максимума при некоторой плотности, которая ниже ёмкости среды. Хищники могут, не уменьшая величины популяции жертвы, изымать из этой популяции такое число особей, которое эквивалентно скорости ежегодного прироста; поэтому та популяция жертвы, которая способна к максимальному пополнению, будет также поддерживать и наибольшее число хищников.
