
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Содержание
Введение……………………………………………...………………………...…3
Глава 1 Структура и функции белков…………………………………......….5
Глава 2 Различие в синтезе пуриновых и пиримидиновых оснований....17
Список литературы………………………………………….…………...…….33
Введение
Живой организм характеризуется высшей степенью упорядоченности составляющих его ингредиентов и уникальной структурной организацией, обеспечивающей как его фенотипические признаки, так и многообразие биологических функций. В этом структурно-функциональном единстве организмов, составляющем сущность жизни, белки играют важнейшую роль, не заменяемую другими органическими соединениями (Березов, 1998).
Белки имеют сложную трёх или четырёх уровневую структуру, которая задаётся последовательностью аминокислот. Аминокислоты благодаря неоднородности и разным характеристикам своих радикалов формируют свойства того или иного белка.
Но аминокислоты важны не только как основное составляющее белковых молекул, они также участвуют в формировании некоторых других веществ, например пуриновых и пиримидиновых оснований в составе пуриновых и пиримидиновых нуклеотидов.
Возвращаясь к белкам, следует отметить, что каждый уровень белковой организации имеет характерные для него связи, которые помогают удерживать определенную пространственную структуру. Пространственная структура (конформация) является важной характеристикой белка, которая отвечает за его способность к разным взаимодействиям.
Исходя из всего, что было сказано ранее, становится ясна причина разнообразия белковых функций. Например, они участвуют в качестве ферментов в специфических реакциях. Белки, могут состоять из нескольких ферментов, формируя целые комплексы, которые также участвуют, например, в тех же реакциях синтеза пуриновых и пиримидиновых оснований.
Синтез пуриновых и пиримидиновых оснований является чрезвычайно важным для организма. Эти основания вместе с пентозой (рибозой или дезоксирибозой) и фосфорной кислотой входят в состав нуклеотидов (Николаев, 2004). Все превращения нуклеотидов начинаются только после синтеза первых пуринового (ИМФ) и пиримидинового (УМФ) нуклеотида. Как известно, полимерами нуклеотидов являются нуклеиновые кислоты (ДНК и РНК), которые имеют первостепенное значение для хранения и реализации генетической информации и для синтеза белка. Таким образом, мы возвращаемся к структурно-функциональному единству живых организмов.
Глава 2. Различия в синтезе пуриновых и пиримидиновых оснований
Основными пуриновыми основаниями входящими в состав ДНК и РНК являются Аденин (6-аминопурин) и Гуанин (2-амино-6-оксипурин), а общим для ДНК и РНК пиримидиновым основанием является только Цитозин (2-окси-4-аминопиримидин), для ДНК специфично пиримидиновое основание Тимин (5-метил-2,4-диоксипиримидин), а для РНК Урацил (2,4-диоксипиримидин) (рис. 1). Тимин и Урацил отличаются только одной метильной группой, которая присутствует в Тимине и отсутствует в Урациле (Берёзов, 1998).
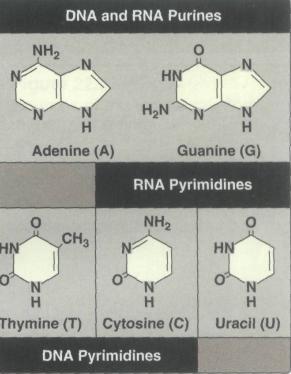
Рис. 1 Пуриновые и пиримидиновые основания, входящие в состав ДНК и РНК.
Азотистые основания – это ароматические гетероциклические соединения, производные пурина и пиримидина (рис. 2). Пиримидин – это шестичленный гетероцикл с двумя атомами азота. Пурин – это гетероцикл, включающий два сочленённых цикла: пиримидиновый и имидазольный (пятичленный цикл, содержащий два атома азота). Молекулы пиримидинов имеют плоское кольцо, а молекулы пуринов – почти плоское, с небольшой складкой (Жеребцов, 2002).
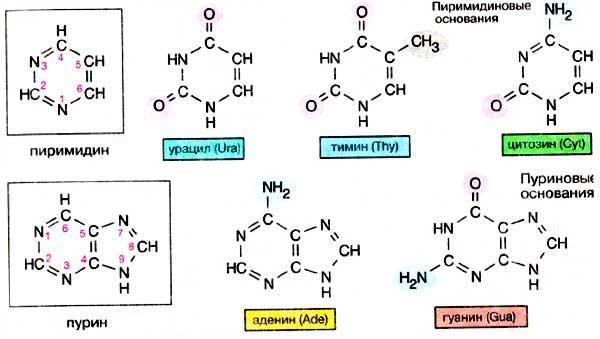
Рис. 2 Пиримидин, пурин и их производные.
Азотистые основания, соединяясь ковалентной связью с 1' атомом рибозы или дезоксирибозы, образуют N-гликозиды, которые называют нуклеозиды. Нуклеозиды, в которых к 5'-гидроксильной группе сахара присоединены одна или несколько фосфатных групп, называются нуклеотидами (рис. 3) (Кольман, Рём, 2004).
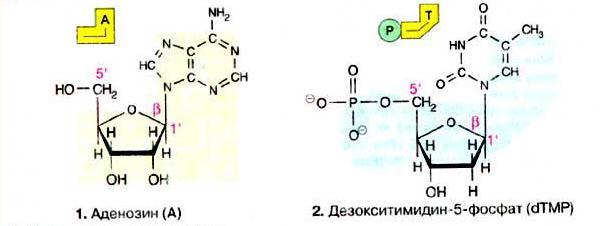
Рис. 3 Нуклеозид (1) и нуклеотид (2).
Путь биосинтеза нуклеиновых оснований довольно сложен, однако этот процесс жизненно необходим почти для всех клеток. И этот путь тесно связан с синтезом нуклеотидов.
Синтез пуриновых оснований
Пуриновые основания, образующиеся в процессе переваривания нуклеиновых кислот в кишечнике, в дальнейшем практически не используются, поэтому их синтез осуществляется из низкомолекулярных предшественников, продуктов обмена углеводов и белков. Нуклеотиды, синтезирующиеся за счёт повторного использования азотистых оснований или нуклеозидов, составляют не более 10-20% общего фонда этих соединений (Николаев, 2004).
Впервые работами Дж. Бьюкенена, Дж. Гринберга экспериментально доказано включение ряда меченых атомов в пуриновое кольцо мочевой кислоты. Скармливая птицам меченые соединения, Дж. Бьюкенен анализировал места включения метки в пуриновое кольцо; полученные данные были в дальнейшем уточнены и подтверждены рядом других исследователей.
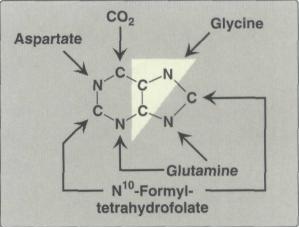
Рис. 4 Происхождение атомов в пуриновом кольце.
Было установлено, что в формировании кольца принимают участие аминокислоты Глицин, Аспарагиновая кислота, Глутамин, СО2 и два одноуглеродных производных тетрагидрофолата: метенил-Н4-фолат и формил-Н4-фолат (рис. 4).
На рисунке показано что 4-й и 5-й атомы углерода и 7-й атом азота в ядре имеют своим источником глицин. Два атома азота (N-3 и N-9) происходят из амидной группы глутамина, один атом азота (N-1) – из азота аспарагиновой кислоты; углеродный атом (С-2) происходит из углерода N10-формил-ТГФК, атом углерода в 8-м положении – из N5,N10-метенил-ТГФК и, наконец, углерод С-6 имеет своим источником СО2. Пуриновый гетероцикл, как видно, собирается в основном из отдельных атомов углерода и азота. Единственным блочным элементом, используемом в этом процессе является молекула глицина (Кнорре, Мызина, 2000).
В настоящее время благодаря исследованиям Дж. Бьюкенена, Дж. Гринберга, А. Корнберга и сотр. полностью расшифрована последовательность включения перечисленных веществ в пуриновое кольцо, установлена природа всех промежуточных соединений и ферментных систем, катализирующих химические реакции синтеза. Интересным оказался факт почти полного совпадения путей синтеза пуриновых оснований в печени животных и у микроорганизмов, в частности у Е. coli и Neurospora crassa. Следует, однако, отметить, что конечным результатом синтеза оказалось не свободное пуриновое основание, а рибонуклеотид – инозиновая кислота (ИМФ), из которой далее синтезируются АМФ и ГМФ. Синтез пуринового кольца состоит из 11 химических реакций, которые будут рассмотрены далее.
Образование 5-фосфорибозил-1-дифосфата (ФРДФ), или фосфорибозилпирофосфата (ФРПФ) занимает центральное место в синтезе как пуриновых, так и пиримидиновых нуклеотидов. Он образуется за счёт переноса в, г-пирофосфатного остатка АТФ на рибозо-5-фосфат в реакции, катализируемой ФРДФ-синтетазой (рис. 5).
Источниками рибозо-5-фосфата могут быть: пентозофосфатный путь превращения глюкозы или катаболизм нуклеозидов, в ходе которого под действием нуклеозидфосфорилазы первоначально образуется рибозо-1-фосфат, а затем с помощью соответствующей мутазы фосфатный остаток переносится в 5-положение.
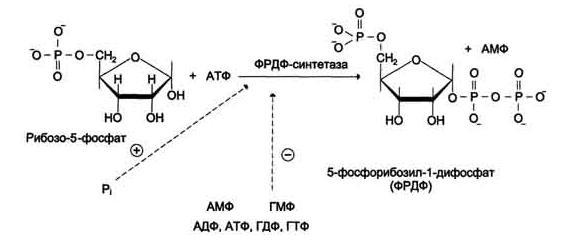
Рис. 5 Образование 5-фосфорибозил-1-дифосфата (ФРДФ).
ФРДФ участвует не только в синтезе пуриновых и пиримидиновых нуклеотидов из простых предшественников (т. е. de novo), но используется на образование пуриновых нуклеотидов по "запасному" пути и в синтезе нуклеотидных коферментов (Северин, 2003).
Образовавшийся 5-фосфорибозил-1-пирофосфат (ФРПФ) взаимодействует с глутамином, являющимся донором NH2-группы, в результате чего образуется в-5-фосфорибозиламин. В реакции наряду с освобождением пирофосфата и свободной глутаминовой кислоты происходит изменение конфигурации 5-фосфорибозиламина (из б - в в-). Таким образом, данная стадия становится ключевой реакцией в синтезе пуринов (рис. 6).
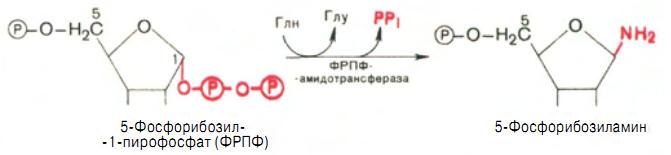
Рис. 6 Образование 5-фосфорибозиламина.
На следующей стадии присоединяется вся молекула глицина к свободной NH2-группе в-5-фосфорибозиламина (реакция нуждается в доставке энергии АТФ) с образованием глицинамидрибонуклеотида (рис. 7).
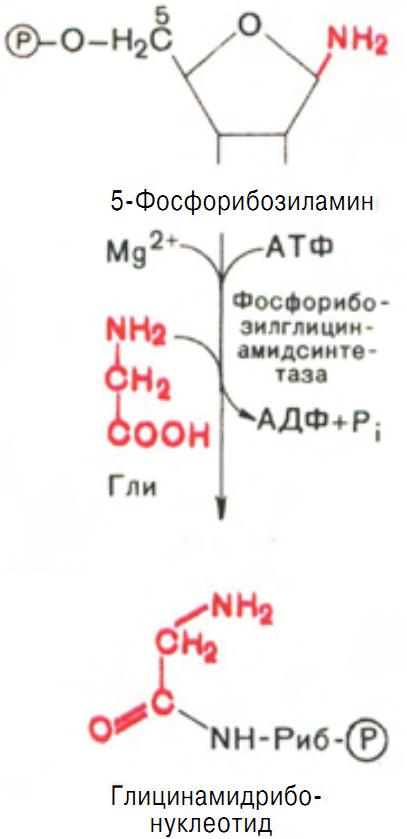
Рис. 7 Образование глицинамидрибонуклеотида.
Затем, на следующей стадии, цепь удлиняется за счет присоединения формильной группы из N5,N10-метенил-ТГФК с образованием формилглицинамидрибонуклеотида (рис. 8).
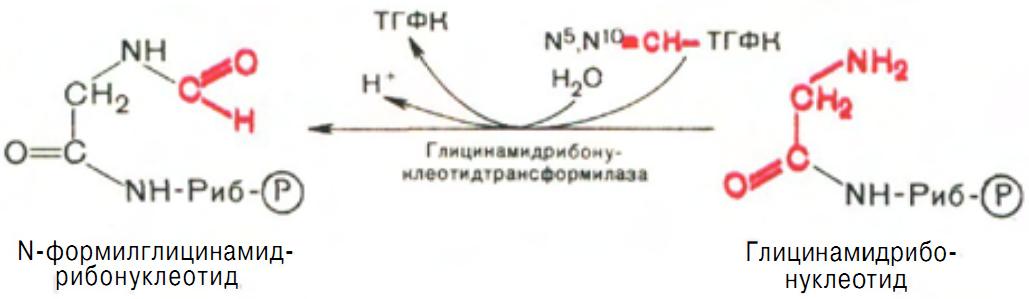
Рис. 8 Образование N-формилглицинамидрибонуклеотида.
На формильную группу последнего переносится далее амидная группа глутамина и синтезируется формилглицинамидинрибонуклеотид (реакция также идет с потреблением энергии АТФ) (рис. 9).
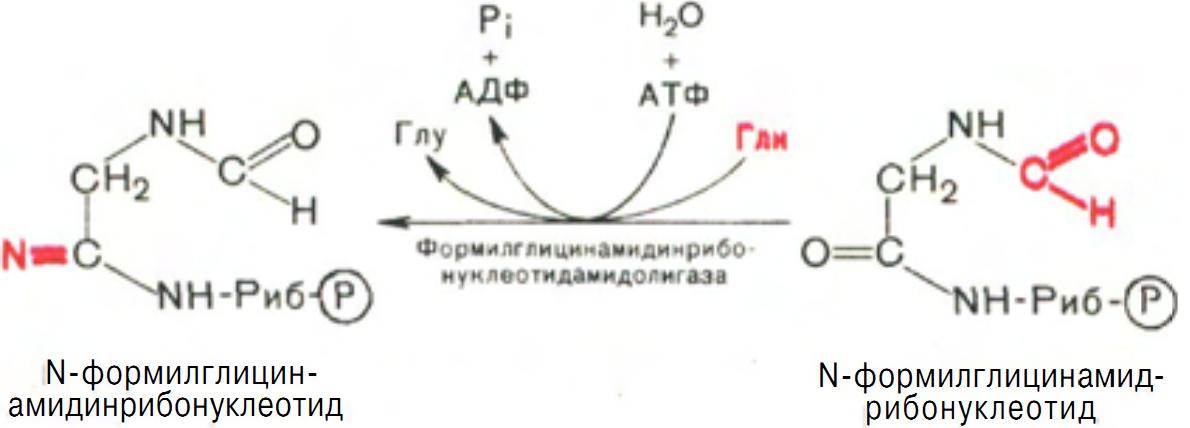
Рис. 9 Образование N-формилглицинамидинрибонуклеотида.
На следующей стадии замыкается пятичленное имидазольное кольцо и образуется 5-аминоимидазолрибонуклеотид (рис. 10).
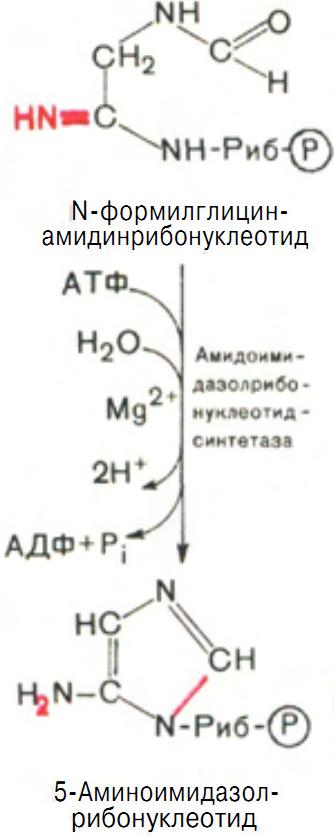
Рис. 10 Образование 5-Аминоимидазолрибонуклеотида.
5-аминоимидазолрибонуклеотид способен акцептировать СО2 с образованием рибонуклеотида 5-аминоимидазол-4-карбоновой кислоты (рис. 11).
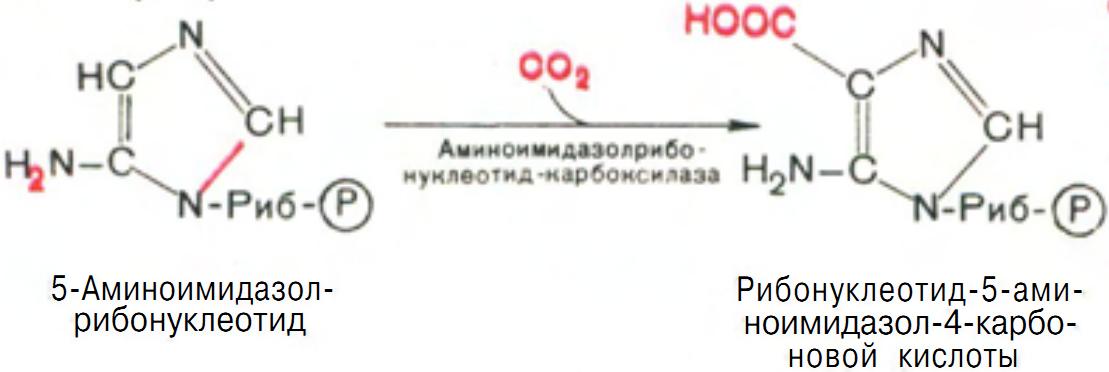
Рис. 11 Образование 5-аминоимидазол-4-карбоновой кислоты.
В последующем двухступенчатом процессе, в котором участвуют аспа-рагиновая кислота и АТФ, сначала азот аспарагиновой кислоты включается в 1-е положение будущего пуринового ядра и образуется 5-аминоимидазол-4-N-сукцинокарбоксамидрибонуклеотид (рис. 12).
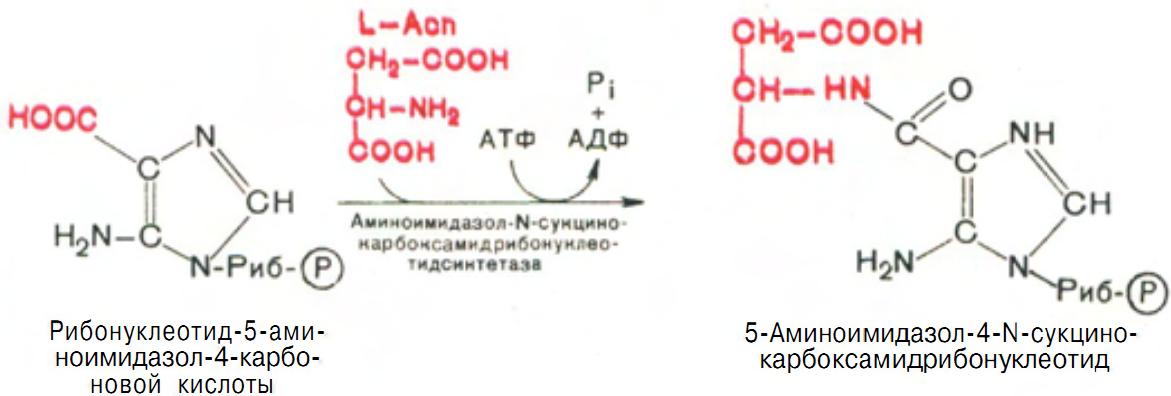
Рис. 12 Образование 5-аминоимидазол-4-N-сукцинокарбоксамидрибонуклеотида
Затем, на второй ступени этого процесса, образуется 5-аминоимидазол-4-карбоксамид-рибонуклеотид и освобождается фумаровая кислота (рис. 13).
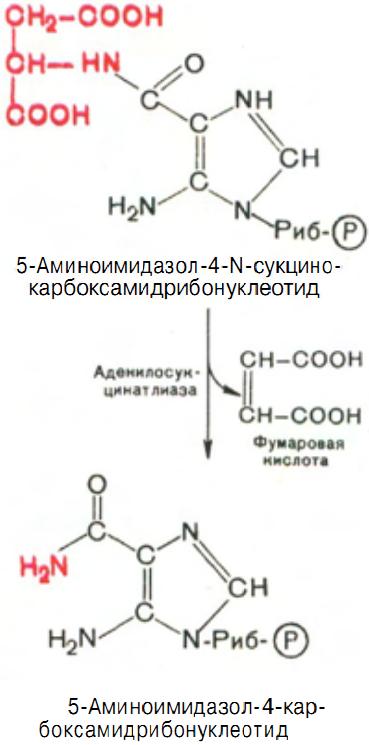
Рис. 13 Образование 5-аминоимидазол-4-карбоксамид-рибонуклеотида.
Последний углеродный атом пиримидинового остатка кольца пурина вводится в виде формильного остатка (источник N10-формил-ТГФК), который присоединяется к 5-NH2-группе. В результате этой реакции образуется 5-формамидоимидазол-4-карбоксамидрибонуклеотид (рис. 14).
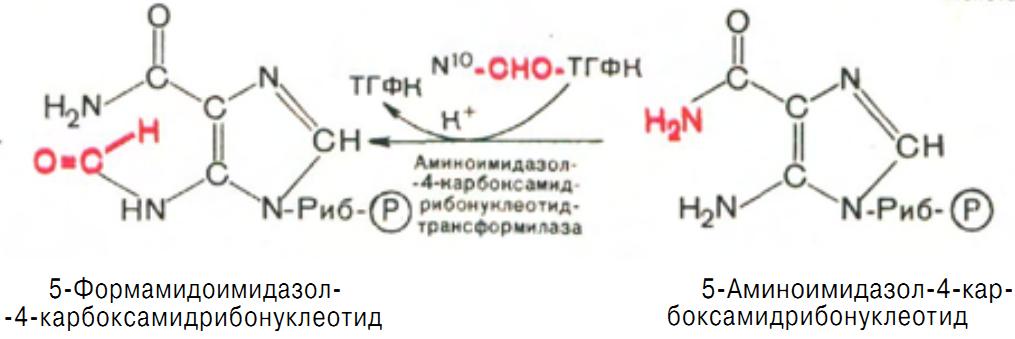
Рис. 14 Образование 5-формамидоимидазол-4-карбоксамидрибонуклеотида.
После этого отщепляется молекула воды и второе кольцо замыкается. В результате образуется первое пуриновое основание в составе первого пуринового нуклеотида – инозина-5'-монофосфата, который является предшественником пуриновых нуклеотидов в составе нуклеиновых кислот (рис. 15).
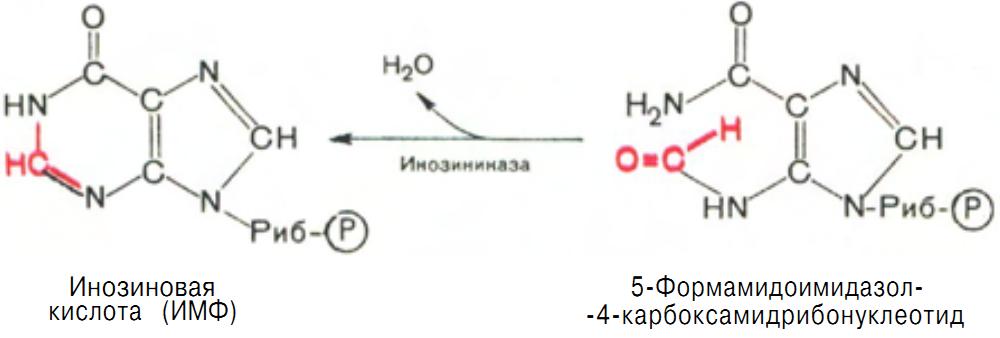
Рис. 15 Образование Инозиновой кислоты (ИМФ).
На синтез инозин-5'-монофосфата (ИМФ) затрачивается не менее шести молекул АТФ. В отличие от прокариотов, у которых каждую стадию этого процесса катализирует отдельный фермент, у эукариотов за счёт слияния генов возникли полифункциональные ферменты, каждый из которых катализирует несколько реакций. В синтезе пуриновых нуклеотидов de novo это реакции 3,4 и 6, 7-8 и 10-11 соответственно.
ИМФ в основном используется на синтез АМФ или ГМФ. Небольшое количество этого продукта обнаруживается также в тРНК в качестве одного из минорных нуклеотидов (Северин, 2003).
