

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Кинетический механизм взаимодействия гемопротеинов с кофактором in vitro на примере провоспалительного фермента простагландин-H-синтазы
Автор:
Международный учебно-научный биотехнологический центр Московского государственного университета им.
Одной из важнейших для организма белковых групп являются гемопротеины. Они выполняют самые разнообразные функции: связывают и переносят кислород (гемоглобин, миоглобин), катализируют окислительно-восстановительные реакции (ферменты цитохромной системы, каталаза, пероксидазы), и т. п. Их объединяет наличие простетической гемовой группы. Наиболее распространённым является гем B (Fe-протопорфирин IX).
Гем может синтезироваться во всех клетках, за исключением эритроцитов. С наибольшей скоростью он образуется в костном мозге и печени.
Железо, необходимое для синтеза гема, поступает с пищей или освобождается при распаде гема. Всасывание Fe происходит в тонком кишечнике с помощью энтероцитов. Абсорбированное железо может накапливаться в виде молекул ферритина, являющегося основным внутриклеточным депо Fe, либо экспортироваться в плазму крови через ферропортин. В плазме транспортную функцию выполняет трансферрин – гликопротеин, синтезирующийся в печени и связывающий только окисленное железо (Fe3+). Поступающее в кровь железо окисляет фермент ферроксидаза.
Нарушения метаболизма железа приводят к серьёзным последствиям для организма. При дефиците железа возникает железодефицитная анемия (ЖДА), характеризующаяся нарушением синтеза гемоглобина, а также уменьшением содержания трансферрина и ферритина. ЖДА часто наблюдается при кровотечениях, беременности, родах и опухолях ЖКТ. С другой стороны, такое наследственное заболевание, как гемохроматоз, характеризуется чрезмерным накоплением железа в тканях и органах, что может спровоцировать развитие цирроза печени, сердечной недостаточности, сахарного диабета, артрита.
В 2001 г. был выделен и описан пептидный гормон гепсидин ([1]), обладающий ярко выраженными антибактериальными свойствами, а также являющийся медиатором воспаления и одним из ключевых регуляторов обмена железа. Повышенная экспрессия гепсидина приводит к анемии хронических заболеваний (АХЗ), которая часто сопровождает различные воспалительные процессы: ревматоидный артрит, пиелонефрит и т. п.
Было обнаружено, что синтез гепсидина может увеличиваться вне зависимости от обмена железа и активности эритропоэза, а именно – под действием бактериальных липополисахаридов и провоспалительных цитокинов. Тем самым, ограничивается доступ Fe к патогенным микроорганизмам.
Одним из важнейших ферментов, участвующих в воспалительных процессах, является простагландин-H-синтаза (PGHS; К. Ф.1.14.99.1), осуществляющая синтез простагландинов из арахидоновой кислоты. Простагландины являются медиаторами, ответственными за воспаление и другие важные физиологические процессы: свёртывание крови, репродуктивные функции, работа желудочно-кишечного тракта. PGHS катализирует две последовательные реакции: циклооксигеназную, в ходе которой арахидоновая кислота окисляется двумя молекулами кислорода и превращается в простагландин G2 (PGG2), и пероксидазную, в результате которой PGG2 преобразуется в PGH2 в присутствии донора электронов. В ходе обеих реакций фермент подвергается необратимой инактивации.
PGHS – мембранный димерный гемопротеин, состоящий из двух структурно идентичных мономеров. Катализ обеих реакций способна проводить только холоформа PGHS (комплекс белка с кофактором гемом).
Исследование взаимодействия фермента с гемом является актуальным для понимания механизма функционирования гемопротеинов и для оценки участия PGHS в процессах метаболизма Fe. Кроме того, открытие гепсидина связывает воедино обмен Fe и воспалительные процессы, в развитии которых PGHS принимает активное участие.
Одним из ключевых моментов в исследовании взаимодействия PGHS и гема является выяснение кинетического механизма образования и распада комплекса фермент-гем. В научной литературе постулируются две возможные кинетические модели:
1) Молекулы гема соединяются с мономерами фермента в соотношении 1:1 ([2], [3], [4]).
Подразумевается, что гем взаимодействует с ферментом по простейшей одностадийной модели:
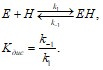
Здесь EH – холоформа мономера фермента, E – апоформа, H – свободный гем, k1 – константа скорости присоединения гема к апоферменту, k-1 – константа скорости отщепления гема от холофермента, Kдис – равновесная константа диссоциации комплекса PGHS-гемин.
2) Мономеры значительно отличаются друг от друга по силе связывания гема, и он в первую очередь присоединяется к мономеру, обладающему более высоким сродством ([5], [6], [7]).
По этой модели PGHS является функциональным гетеродимером и проявляет отрицательную кооперативность по отношению к гему.
В данных работах проводили титрование апофермента гемом, измеряли оптическое поглощение в полосе Соре и строили график зависимости поглощения от концентрации
гема. Выводы о разном сродстве мономеров к гему делали по наличию двух участков графика со значительно отличающимся наклоном.
Мы воспроизвели эти эксперименты, измерив поглощение комплекса фермент-гем на 402 нм относительно поглощения свободного гема.
Апоформу фермента получали методом восстановления гемового железа дитионитом натрия в анаэробных условиях и дальнейшего окисления системы кислородом.
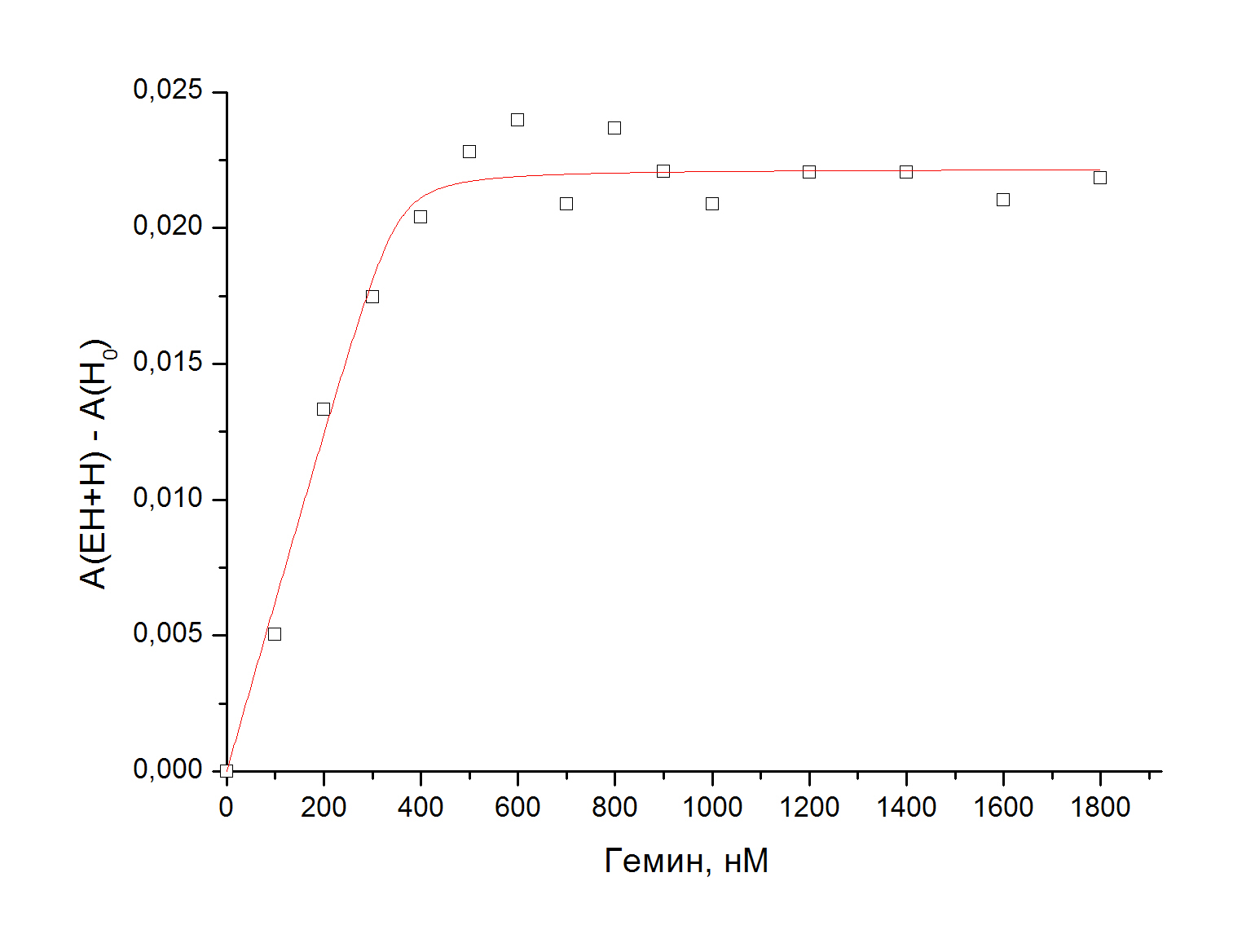
Видно, что есть только один участок связывания гема, и предположение о кооперативной модели связывания не выполняется.
Непосредственное определение констант скорости прямой и обратной реакций для комплекса PGHS-гем, а также равновесной константы диссоциации по измерению поглощения в полосе Соре оказалось весьма затруднительным, поскольку концентрация фермента и гема в системе существенно выше, чем предполагаемый диапазон значений равновесной константы. Ранее в работах [2] и [6] оценивали равновесную константу диссоциации, однако в [2] она была измерена косвенным путём в присутствии другого гемопротеина – человеческого альбумина, а в [6] – по титрованию апофермента гемом в полосе Соре, с существенной погрешностью. Индивидуальные константы скорости реакций процесса комплексообразования не определялись.
В настоящем исследовании кинетику взаимодействия PGHS с гемом изучали с помощью детекции скорости циклооксигеназной реакции при 25°C для ассоциации и диссоциации холоформы PGHS в зависимости от времени инкубации. Зависимости, полученные для 27 нМ фермента и 30 нМ гема, показаны ниже.
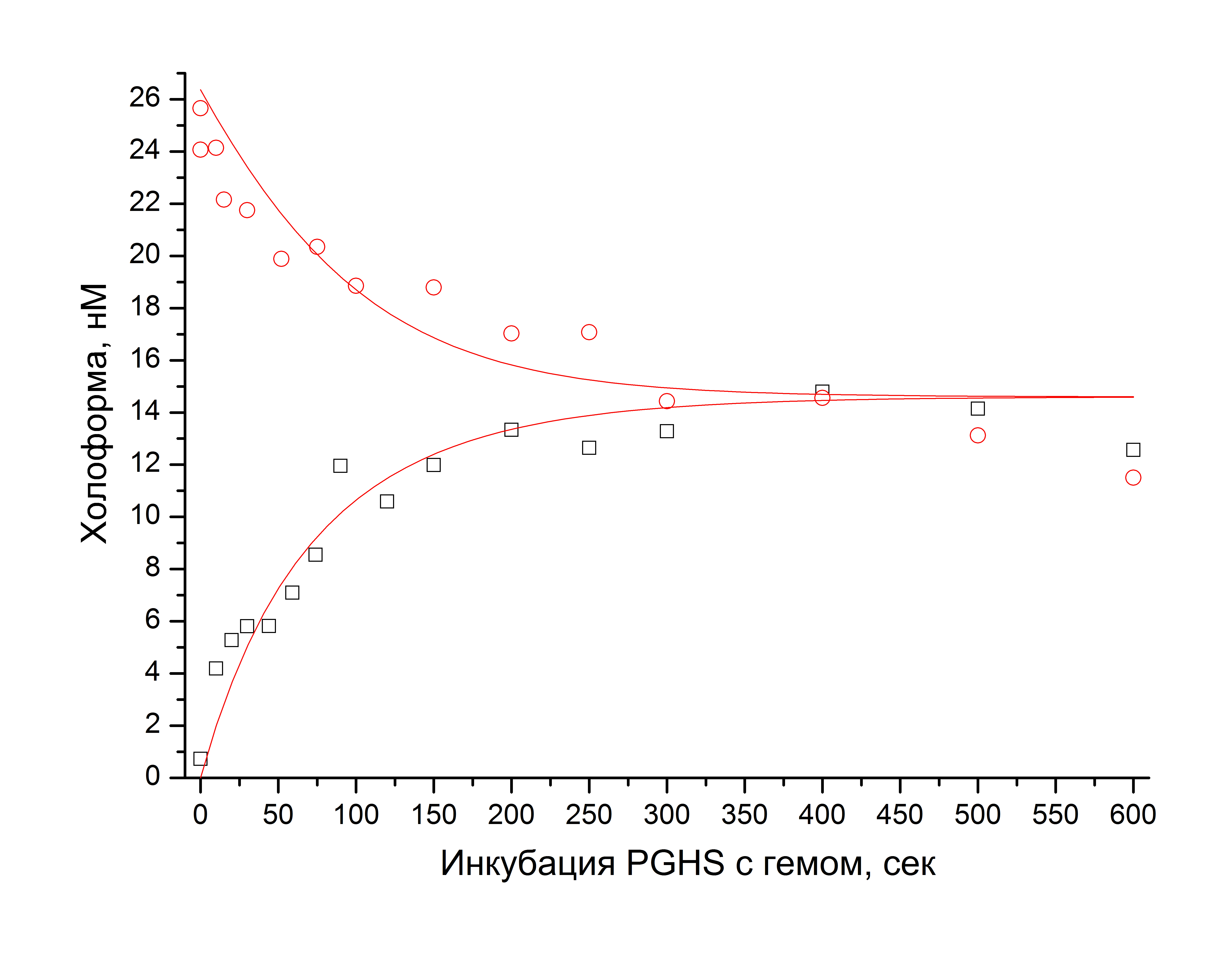
Данные были аппроксимированы зависимостью вида
![]()
выведенной для простейшей одностадийной модели. В параметры A, B, C, D, E заложены значения концентрации фермента и гема, начальной доли холофермента, коэффициента разбавления комплекса фермент-гем в реакционной ячейке, а также константы скорости прямой и обратной реакций.
Для простейшей модели найдены значения константы скорости присоединения гема к апоферменту (3*105 М-1*сек-1) и константы скорости отщепления гема от холофермента (3,9*10-3 сек-1). Соответственно, равновесная константа диссоциации составляет 1,3*10-8 М.
Выводы:
1) Гипотеза о кооперативных взаимодействиях, проявляемых простагландин-H-синтазой при связывании гема, не подтверждается.
2) Для простейшей одностадийной модели найдены значения констант скорости присоединения гема к апоферменту и его отщепления от холофермента, а также значение равновесной константы диссоциации комплекса PGHS-гем.
Литература:
1) Park C. H., Valore E. V., Waring A. J. et al. Hepcidin: a urinary antibacterial peptide synthesized in the liver. J. Biol. Chem. 2001; 276: 7806-7810.
2) Ueno R., Shimizu T., Kondo K., Hayaishi O. J Biol Chem. 1982; 257(10): 5584-5588.
3) Roth G. J., Machuga E. T., Strittmatter P. J Biol Chem. 1981; 256(19): 10018-10022.
4) Garavito R. M., DeWitt D. L. Biochim Biophys Acta. 1999; 1441(2-3): 278-287.
5) Kulmacz R. J., Lands W. E. J Biol Chem. 1984; 259(10): 6358-6363.
6) Dong L., Vecchio A. J., Sharma N. P., Jurban B. J., Malkowski M. G., Smith W. L. J Biol Chem. 2011; 286(21): 19035-19046.
7) Zou H., Yuan C., Dong L., Sidhu R. S., Hong Y. H., Kuklev D. V., Smith W. L. J Lipid Res. 2012; 53(7): 1336-13347.
