


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Применение диатомовых водорослей в нанотехнологиях
ФИО автора
Учебное заведение Ришельевский лицей, Одесса
Научный руководитель
Одесса, 2011
Оглавление
Постановка задачи 3
Применение диатомовых водорослей в нанотехнологиях 4
Геном диатомовых водорослей 7
Гипотезы об ассимиляции кремния в диатомовых водорослях 12
Цитированная литература 19
Использованная литература 21
Постановка задачи
Данная работа обобщает известные факты о жизнедеятельности диатомовых водорослей, их геноме и ставит перед собой следующие задачи:
Изучить применение диатомовых водорослей в нанотехнологиях. Проанализировать результаты исследований о структуре панцирей диатомовых водорослей и механизмах создания панциря. Предложить возможные исследования, которые необходимо провести для более подробного изучения вопроса генетического контроля синтеза аморфного кремния, что может найти широкое применение в нанотехнологиях.Применение диатомовых водорослей в нанотехнологиях
Диатомовые водоросли применяются во многих отраслях промышленности и науки.
За счет того, что панцири маленьких по линейным размерам видов диатомовых водорослей содержат в себе поры, их используют в качестве фильтров.
Диатомит, который на 60% состоит из панцирей диатомовых водорослей, используется как строительный материал.
Также по активности жизнедеятельности диатомовых водорослей судят об экологическом благополучии водоемов.
Еще одно использование диатомовых водорослей основано на том, что по их отложениям можно судить о палеоклиматических условиях. По этому принципу была прочитана Байкальская «диатомовая летопись».
Однако, наиболее перспективным направлением использования диатомовых водорослей являются нанотехнологии.
Диатомовые водоросли интересуют исследователей, как живые организмы способные структурировать химические элементы в наноструктуры в своем панцире. Известно, что существуют определенные закономерности в расположении пор мельчайших размеров, что доказывается периодичностью узора и наличием у каждого элемента узора геометрического центра.
Технологии на основе аморфоного кремния позволяют решить проблемы низкой подвижности носителей зарядов в электронных приборах, таких, как транзисторы, и значительной утечки тока, обусловленной высокой концентрацией точечных дефектов внутри пленки.
Изготовление тонкопленочных наноструктурных материалов на основе кремния с использованием вакуумно-плазменного осаждения кремния позволяет производить дешевые электронные приборы со сравнительно стабильными рабочими параметрами.
Элементы памяти на основе нанокристаллического кремния могут быть выполнены с использованием схемы тонкопленочного транзистора с тонким слоем SiO2, позволяющим осуществлять процессы записи/считывания/стирания информации. В этом случае основную роль играет химический состав интерфейса Si-SiO2, заряды которого влияют на процессы туннелирования.
Второе возможное устройство элемента памяти основано на наличии в материале пленки атомов кислорода в различном состоянии, которые, с одной стороны, формируют диполи Si-O, а с другой, – оказывают влияние на миграцию точечных дефектов. Такая память, полученная при нанесенной пленке Si на тонкий слой CsO2, позволяет создавать тонкопленочные структуры со стимулированным электрическим полем изменением фазового состава пленки, который обратимо воссоздается при нагревании.
Третий вариант устройства памяти – переключаемая электрическим полем спектральная компонента ВЧ-диапазона от клика пленки, полученная с помощью электронного парамагнитного резонанса. Такое переключение осуществляется за счет присутствия внутри пленки точечных дефектов, таких, как А-дефекты Максимально возможная частота переключений состояний А-дефектов – 1013 Hz. Однако частота переключений состояний памяти ограничивается наличием в пленке границ раздела нанокристаллов, внутри которых присутствуют точечные дефекты, формирующие флуктуации потенциала. Чем меньше размер кристаллов, тем такие флуктуации меньше: Использование более высоких частот для переключения возможно только с применением оптических методов. [1]
Диатомовые водоросли предполагают использовать для создания солнечных батарей. Сначала они образуют колонию, покрывая необходимый участок, затем органический компонент удаляется, а в поры помещают оксид титана. Таким образом, создается n-p полупроводник в тонкой пленке, который под воздействием солнечного света (фотонов) производит энергию (dye-sensitized solar cells). [2, 3]
Панцири диатомовых водорослей без органического компонента можно использовать в литий-ионных аккумуляторах, они будут предавать технологии большую функциональность, поскольку в этом материале множество мелких нанопор, что вызывает нестандартные квантовые эффекты, отражающиеся на уровне энергии, вырабатываемой батареей. Также за счет нанопор достигается увеличение площади взаимодействия между литием и кремнием.
Однако наибольший интерес представляла бы технология направленного выращивания структур пористого кремния. Именно поэтому далее в работе мы рассмотрим механизм образования панциря диатомовых водорослей, обобщим уже сделанные предположения и предложим направления дальнейших исследований.
Геном диатомовых водорослей
Диатомовые водоросли – группа водорослей, представленных одноклеточными и колониальными формами жизни. Широко распространены по всему земному шару, способны выжить в любой влажной среде, за счет кремниевого панциря хорошо приспосабливаются к широкому интервалу давлений и температур.
Диатомовые водоросли разделяются на два класса по типу радиальной симметрии: центрические и пеннатные. Центрические водоросли разделяются на три отряда (пеннатные – на четыре), которые в свою очередь делятся на семейства и виды. [4]
С помощью метода молекулярных часов удалось установить, что диатомовые появились около 230 млн лет назад. Вероятно, первые диатомеи были жгутиковыми клетками (в настоящее время их относят к Heterokonta – разножгутиковым, так как на некоторых стадиях жизненного цикла они имеют один жгутик с тончайшими волосками). [5]
Самые ранние отложения диатомей были найдены в Корее в осадках, сформировавшихся на суше. Была высказана гипотеза, что океан, отступивший около 230 млн лет назад, оставил за собой лужи, где и обитали жгутиковые диатомеи. Они стали использовать кремнезем, чтобы переживать в стадии покоя неблагоприятные условия.
Древние диатомеи потребляли так много кремнезема, что он начал осаждаться в клетках, по-видимому, сначала в виде маленьких чешуек, которые выдавливались из клеток и создавали вокруг них защитную оболочку, препятствующую обезвоживанию при высыхании мелководных водоемов. В конце концов, две чешуйки превратились в створки панциря, а остальные – в поясковые ободки, соединяющие створки.
Древнейшие находки ископаемых центрических диатомей известны из отложений юрского периода (около 180 млн лет назад). Пеннатные диатомеи вдвое моложе: они известны лишь начиная с позднего мелового периода (90 млн лет назад); при этом они гораздо разнообразнее и многочисленнее своих центрических предков. [6,7]
Диатомовые водоросли имеют характерные уникальные особенности, связанные с их эволюционным развитием:
Наличие 4-х мембран вокруг хлоропластов. Две внутренние мембраны соответствуют мембранам, окружающим пластиды высших растений, вторая из них (считая снаружи) происходит от плазматической мембраны гетероконта, а внешняя – продолжение ЭПР хозяйской клетки. Кремнистый панцирь из аморфного кремния. Сложный узор панциря точно воспроизводится в каждом поколении, т. е. является видоспецифическим и используется при таксономической идентификации водорослей. [8] Размножаются диатомовые водоросли делением; каждая дочерняя клетка получает половину материнского панциря, другая вырастает заново, при этом старая половина охватывает своими краями новую. Из-за чего происходит мельчание водорослей, т. е. размеры каждого следующего поколения меньше предыдущего. У диатомей также наблюдается половое размножение с образованием ауксоспор. Согласно наиболее распространенному мнению, достигнув из-за размножения делением минимальных размеров, клетки развивают ауксоспоры, что приводит к восстановлению их размеров. Однако другие исследователи считают, что ауксоспорообразование связано просто со старением клеток, так как его нередко удавалось наблюдать и тогда, когда клетки еще не достигли своих минимальных размеров. Кроме того, есть наблюдения, свидетельствующие о развитии ауксоспор при изменении условий окружающей среды, например при резком понижении температуры воздуха или воды. [9]В 2004 году был расшифрован геном диатомеи Thalassiosira, а в 2008 Phaeodactylum tricornutum, причем первый вид относится к центрическим водорослям, а второй – к пеннатным.
Имея в своем распоряжении два полных генома диатомей, ученые смогли сделать ряд важных выводов об эволюции группы.
Один из основных выводов состоит в том, что эволюция диатомей происходила на удивление быстро. Лишь для 57% генов пеннатной диатомеи нашлись гомологи (близкородственные гены) в геноме центрической диатомеи. Аминокислотные последовательности белков у двух диатомей совпадают только на 54,9%.
Сравнительная таблица геномов водорослей видов Phaeodactylum и Thalassiosira
Пеннатная диатомея Phaeodactylum | Центрическая диатомея Thalassiosira | |
Размер генома | 27,4 млн пар нуклеотидов | 32,4 млн пар нуклеотидов |
Число генов | 10402 | 11776 |
Число генов, встречающихся во многих группах эукариот | 3523 | 4332 |
Число генов, встреченных у обеих диатомей и больше ни у кого (цифры в этой строчке не одинаковы из-за различного числа генных дупликаций у двух видов диатомей) | 1328 | 1407 |
Число генов, уникальных для данного вида | 4366 | 3912 |
Число интронов (некодирующих вставок в генах) | 8169 | 17880 |
Доля мобильных генетических элементов — ретротранспозонов класса LTR | 5,8% | 1,1% |
Особенностью эволюции центрических диатомей было приобретение (по-видимому, сравнительно недавнее) большого количества новых интронов — некодирующих вставок в генах. У пеннатных диатомей массового распространения интронов не было, зато в их геномах бурно размножились мобильные генетические элементы — ретротранспозоны. Проанализировав дополнительные данные по другим видам пеннатных диатомей, авторы пришли к выводу, что активность ретротранспозонов была важным фактором, способствовавшим росту видового разнообразия этой группы. [10] Перепрыгивая с место на место внутри генома, ретротранспозоны могут влиять на активность соседних генов и способствовать росту генетической изменчивости.
Как уже упоминалось, диатомеи вместе с бурыми и золотистыми водорослями и некоторыми другими эукариотами входят в состав группы Heteroconta. Считается, что гетероконты появились около 1 млрд лет назад в результате симбиоза гетеротрофного (нефотосинтезирующего) одноклеточного организма с одноклеточной же красной водорослью. У красных водорослей, как и у зеленых растений, хлоропласты (органеллы, служащие для фотосинтеза) являются первичными, то есть происходят напрямую от симбиотических цианобактерий. Первичные хлоропласты всегда окружены двумя мембранами. Предки гетероконт проглотили одноклеточную красную водоросль и превратили ее в фотосинтезирующего симбионта. Впоследствии от клетки красной водоросли почти ничего не осталось, кроме внешней оболочки и хлоропласта. Гены симбиотической красной водоросли были отчасти утрачены, отчасти — перешли в геном хозяина. Поэтому хлоропласты гетероконт называют «вторичными», и они окружены не двумя, а четырьмя мембранами (из них две внутренние — это оболочка первичного хлоропласта, третья изнутри — бывшая оболочка клетки красной водоросли, а четвертая, внешняя — это оболочка пузырька-вакуоли, в которую была заключена проглоченная красная водоросль).
Для проверки этой теории был предпринят целенаправленный поиск в геномах диатомей генов, похожих на гены красных водорослей. [10] Поиск увенчался успехом: удалось выявить более 170 генов, унаследованных предками диатомей от красных водорослей. Подавляющее большинство этих генов необходимо для работы хлоропластов. Этот результат — очень весомый довод в пользу того, что сложившиеся на сегодняшний день представления о ранней эволюции эукариот в общих чертах верны.
Также в геномах диатомей обнаружилось очень много генов, которые явно были заимствованы диатомеями у различных прокариот: цианобактерий, протеобактерий, архей и других. В геноме Phaeodactylum обнаружено 587 таких заимствованных генов. На сегодняшний день это рекордное количество генов прокариотического происхождения, найденных в эукариотическом геноме. Более половины из этих генов (56%) есть также и у Thalassiosira. Эти гены, скорее всего, были заимствованы диатомеями у бактерий довольно давно — еще до расхождения эволюционных линий центрических и пеннатных диатомей. Остальные 44%, по всей видимости, были заимствованы предками Phaeodactylum уже после этого события, то есть в течение последних 90 млн лет.
Бактериальные гены, по-видимому, значительно расширили биохимические возможности диатомей. Эти гены помогают диатомеям осуществлять ряд биохимических реакций, не свойственных другим эукариотам: они участвуют в ассимиляции кремния, производят сигнальные и светочувствительные белки. [10]
Гипотезы об ассимиляции кремния в диатомовых водорослях
Рассмотрим функции кремния. Прежде всего, отметим, что кремний усваивают не только диатомовые водоросли, но и радиолярии, некоторые виды губок, некоторые виды дрожжей и элементы транспорта кремния есть у некоторых растений, которые создают ребра из кремния (некоторые виды риса).
При помещении диатомовых водорослей в водную среду без кремния у них истончается панцирь, нарушаются процессы размножения. Аналогичный эффект наблюдается пи добавлении в нормальную питательную среду, содержащую германий. [9]
Большинство исследований биохимического состава панцирей выполнено на пеннатном виде Cylindrotheca fusiformis (см работы Kroeger & Sumper) [8]. Эти работы привели к открытию новых пептидов - силаффинов, которые могут участвовать в основном процессе биоминерализации в SDV (Kroeger & Sumper, 1999). Эти катионные пептиды разных размеров синтезируются из полипептидов-предшественников и характеризуются наличием Лиз-Лиз-повторов, к которым в ходе модификации ковалентно присоединены новые химические остатки, такие как олиго-N-метил полиамины (Kroeger et al., 2001). Показано, что силаффины могут запускать формирование кремниевых наносфер in vitro (Kroeger et al., 1999). [8]
Другим важным органическим компонентом биокремнезема диатомей являются длинноцепочечные полиамины, производные путресцина. Путресцин относится к группе биогенных аминов, образуется при дикарбоксилировании аминокислоты орнитин. Как и силаффины, они индуцируют быстрое осаждение кремния in vitro, формируя наносферы размером от 100 нм до 1 мкм в зависимости от использованного полиамина (Kroeger et al., 2000). Разные диатомеи используют разные силаффины и фрустулины, которые определяют видоспецифические отличия осаждения кремния вне клетки. [8]
Другие белковые компоненты диатомовых створок – фрустулины и плевралины (сначала называвшиеся НЕР-белками; Kroeger et al., 1997; Kroeger & Wetherbee, 2000). Как и силаффины, плевралины тесно связаны с кремнием и выделяются из створок только при полном растворении кремния плавиковой кислотой Плевралины кодируются небольшим семейством генов и характеризуются присутствием повторяющихся аминокислотных мотивов.
Локализация плевралина точно ограничена терминальным поясковым ободком (плевралиновым ободком) эпитеки в районе перекрывания с гипотекой. В ходе клеточного деления он оказывается ассоциированным с плевралиновым ободком гипотеки в тот момент, когда родительская гипотека функционально превращается в эпитеку дочерней клетки (Kroeger & Wetherbee, 2000). Это замечательное наблюдение свидетельствует о том, что дифференциция эпи - и гипотеки находится под строгим контролем развития. [8]
Плевралин адресован не в SDV, а секретируется прямо в щель, образующуюся при расхождении дочерних клеток (Kroeger & Wetherbee, 2000).. Связывание с терминальным поясковым ободком гипотеки, следовательно, происходит во внеклеточном пространстве. Секреция, вероятно, идет по общему механизму экзоцитоза, однако является интригующим примером транспорта, направленного в пространство щели расхождения. Интересно определить, сколько еще имеется белков, связанных с панцирем, не секретируемых в пузырьках, депонирующих кремний, однако включающихся в створки. Фрустулины наиболее слабо связаны с клеточной стенкой в сравнении с плевралинами и силаффинами, экстрагируются ЭДТА. К настоящему времени описано 5 фрустулинов, все из которых гликопротеины, способные связываться с Са благодаря присутствию EF-элементов (Kroeger et al., 1994). Они также характеризуются цистеин-богатыми доменами с неизвестной функцией. Фрустулины, по-видимому, не участвуют в отложении кремния, но образуют внешнюю оболочку клетки (van de Poll et al., 1999). [8]
Минимальная концентрация кремнезема, необходимая для роста диатомей, равна 0,5 мг SiO2/л. Как диатомовые водоросли перерабатывают неорганические соединения кремния в органические, точно неизвестно. Возможно, в этом участвуют специальные ферменты, и цепочка реакций предполагает образование кремниевых эфиров.
Известно, что у диатомовых водорослей есть белки-транспортеры органических производных ортокремниевой кислоты - SIT, способные «закачать» в клетку кремния до концентраций в 250 раз превышающих его концентрацию в окружающей среде (Martin-Jezequel et al., 2000). Методом имунноэлектронной микроскопии показали, что доступные для антител детерминаты белка SIT экспонированы в ареалах, отверстиях кремниевого панциря диатомеи с внутренней стороны полисахаридного слоя, расположенного между панцирем и плазматической мембраной (Петрова, 2007). В геноме арабидопсиса гомологов SIT нет, хотя можно предположить их наличие у риса, образующего окремненные ребра на своих листьях (Raven, 1983). [11]
Формирование новых створок подробно изучено на ультраструктурном уровне (Pickett-Heaps et al., 1990), и ключевые моменты этого процесса могут быть представлены следующим образом:
Ядро каждой из дочерних клеток перемещается в ту часть клетки, где будет формироваться новая гипотека. Миктротрубочковый центр позиционируется между ядром и плазматической мембраной, над которым, в итоге, разместится гипотека Между микротрубочковым центром и плазматической мембраной происходит формирование SDV – пузырьков, депонирующих кремний - на участке, который становится центром формирования узора SDV удлиняются в трубочки и затем распространяются в перпендикулярном направлении, формируя огромный везикул, вытянутый вдоль одной стороны створки.В условиях направленного транспорта кремния, белков и полисахаридов внутри SDV формируется новая створка. В ходе процесса содержимое SDV закисляется в результате чего происходит полимеризации кремния (Vrieling etal., 1999). Одни органические компоненты формируют оболочку вокруг кремниевого скелета, другие участвуют в отложении кремния. По завершении биогенеза происходит экзоцитоз створки путем слияния мембраны SDV (силикалеммы) и плазматической мембраны. Считается, что впоследствии внутренняя часть силикалеммы становится новой плазматической мембраной
.Наиболее подробная работа по поводу транспортных SIT-белков с описанием всех использованных методик – «Identification of Proteins from a Cell Wall Fraction of the Diatom Thalassiosira pseudonana. Insights into Silica Structure Formation», написанная Luciano G. Frigeri, Timothy R. Radabaugh, Paul A. Haynes and Mark Hildebrand. [8]
Кратко результаты всех работ до 2010 года можно изложить так:
Сделан биохимический анализ состава панциря Найдена система оптимальных условий для диатомовых водорослей (помогло в исследованиях, поскольку по параметрам, полученным в этой системе, можно было сравнивать результаты разных научных работ) Предложена гипотеза о существовании SDV Выделены белки, отвечающие за активный транспорт кремния (белки SIT) Локализовано месторасположение детерминантов белков SIT Проведен анализ генома диатомей, предположили, какие участки генома за что отвечают. Проведены исследования аксенических культур диатомей. [12, 13, 14]Не решены следующие проблемы:
Цепочка биохимических преобразований кремния Нет доказательств наличия SDV или аналогичной системы Не известны механизмы создания узоров, соответственно не известны причины видовой специфичности узоров Система транспорта кремния в клеткуИтак, поскольку ферменты, участвующие в синтезе кремниевых скелетов или створок через образование кремнийорганических соединений выделить еще не удалось, можно использовать подход от противного: если найти ферменты-антиподы, расщепляющие связь Si-O-С (которая присутствует в сформированных скелетах), то, значит, должны существовать и ферменты, которые синтезируют соединения с такими связями. [11]
Существует такое кремнийорганическое соединение — хпорметилсилатран, больше известное под названием мивал.
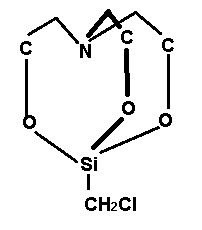
Его биологические свойства изучали академик и его ученики. При изучении дрожжей Rhodotorula mucilaginosa авторы работы «Кремний в биосфере» Ульяновского государственного Университета выделили и охарактеризовали ферменты, гидролизующие или расщепляющие Si-O-С связи - это внеклеточная, образующаяся только в присутствии кремнийорганических соединений, неспецифическая гидролаза, активируемая ионами меди. Как оказалось, такая гидролаза расщепляет Si-O-С связи не только в мивале, но и в других подобных соединениях кремния, когда группа СН2С1 замещена на другие радикалы, причем скорость гидролиза эфиров увеличивается в ряду: СН3 < СН2Сl < CF3CH2CH2 < С6Н5.
Диатомовые водоросли легко гидролизовали все эти силатраны, только гидролизующие ферменты у них так и не удалось пока выделить. В продуктах гидролиза, которые располагались в области слизистой капсулы дрожжей (где находится и сам фермент), обнаружились триэтаноламин и кремнийсодержащий полимер.
При очень большой концентрации мивала вся капсула дрожжей покрывается слоем полисилоксана — продуктом гидролиза дрожжей.
Исходя из биохимического анализа панциря, можно сказать, что не все белки, входящие в его состав транспортируются от SDV к панцирю. Также есть предположения, что активный транспорт с участием SIT-белков запускается благодаря сигналу извне. [11]
Это хорошо сопоставляется с исследованиями 2010 года [14], на которые еще не ссылается ни одна современная работа в англоязычном и русскоязычном интернете. Это исследования аксенических культур диатомовых водорослей, которые показывают, что образование панциря зависит от наличия бактерий в среде, в связи, с чем авторы высказали гипотезу о том, что диатомеи реагируют на некие сигнальные белки бактерий и с ними связана экспрессия соответствующих бактериальных генов.
Также был реализован достаточно интересный подход в компетенции современной биоинформатики - попытка через белки, входящие в состав панциря, выйти на генетический аппарат, определяющий создание панциря. В общем-то, это относится к более глобальной проблеме о связи генотипа и фенотипа. Идея в том, чтобы попытаться соотнести матричное РНК и белки, получаемые в результате. Очевидно, что четкой коллинеарности наблюдаться не будет. Требуется учитывать и мобильные генетические элементы, и полиморфизм, и вырезание интронов при трансляции и другие процессы. Полученные результаты сравнивали с уже имеющимися предположениями о генах, которые это определяют, определенные через поочередную блокировку экспрессии участков генома. [10, 15]
Для дальнейшего изучения диатомовых водорослей необходимо сделать следующее:
Используя данные о том, какие гены отвечают за транспорт кремния, провести генную модификацию другого одноклеточного живого организма и посмотреть, как отразится на метаболизме внедрение системы ассимиляции кремния Попытаться провести аналогичные исследования по анализу генома радиолярий Исследовать поведение диатомовых водорослей в стрессовых ситуациях, меняя концентрацию кремния, воздействовать Ge.Теперь вспомним рассуждения об узорах панцирей. Гипотеза Гордона и Друма (Gordon & Drum, 1994) [8], на которые ссылаются некоторые работы, о том, что начальные этапы биоминерализации происходят через физико-химическое заполнение пространства кремнием, совершенно не объясняет происхождение видоспецифических орнаментов.
Однако, как известно характеристики цитоплазмы, например, интервалы скорости движения цитоплазмы, специфичны для каждого вида. Сама скорость движения цитоплазмы зависит от освещенности, температуры, концентрации питательных веществ. Косвенно о скорости движения цитоплазмы можно судить по электрофоретической подвижности частиц. Электрофоретическая подвижность – это скорость движения частицы (обычно выражаемая в см/c) при напряженности электрического поля в 1 В/см. Эта величина имеет размерность см2 ⋅ с-1⋅ В-1, а её знак совпадает со знаком суммарного заряда макромолекулы Известно, что митохондрия является специфической органеллой, которая способна регулировать заряд на своей поверхности, а значит и заряд в цитоплазме. Очевидно, что направленное движение SIT-белков и других органических производных ортокремниевой кислоты происходит направленно из-за поляризации клетки при размножении и из-за постоянного направления движения цитоплазмы в клетке. О подобных аспектах речь идет в гипотезе Шмидта, 1994 года. Он предполагает, что направленная «укладка » кремния определяется тем, что митохондрии в клетке располагаются на определенной дистанции вдоль пути следования производных ортокремниевой кислоты.
Для анализа узоров можно использовать такие подходы:
Математическое моделирование узоров панциря определенного вида, попытка рассчитать центры геометрической конструкции. Предположение об управлении построением узоров сигнальными белками типа Wnt, способными направлять молекулы Предположение о связи с рецепторной системой, которая может реагировать на сигналы извне в виде химических соединений, вырабатываемых бактериями Смоделировать электрическое поле, способное организовать частицы таким образом, используя корелляцию между созданием структур из химических компонентов и распределению заряда в клетке.