

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Федеральное государственное автономное
образовательное учреждение
высшего профессионального образования
«СИБИРСКИЙ ФЕДЕРАЛЬНЫЙ УНИВЕРСИТЕТ»
Институт фундаментальной биологии и биотехнологии
Базовая кафедра биотехнологии
РЕФЕРАТ
Разработка метода оценки физического состояния спортсменов с использованием биолюминесцентной системы светящихся бактерий
Преподаватель
Студент ФБ13-01М
Красноярск 2014
Содержание
Введение 1
1.1 Биохимия слюны 2
1.2 Изменение состава слюны при физических нагрузках 6
2.1 Биолюминесцентная реакция, катализируемая бактериальной люциферазой 8
2.2 Кинетические особенности функционирования биферментной системы НАДН:ФМН-оксидоредуктаза-люцифераза 10
Список литературы 12
Введение
Совершенствование современной теории и методики тренировочного процесса в спорте высших достижений, во многом определяется расширенным внедрением научных методов управления и современной научно-исследовательской аппаратуры в практику спорта.
Оптимальность управления тренировочным процессом реализуется посредством анализа индивидуальных реакций организма на предлагаемые тренирующие воздействия, периодичности процессов адаптации соответствующих функциональных систем, а также стадийности процессов приспособления при генерализованном или локальном утомлении.
Методы индивидуализации тренировок возможны только при использовании объективных критериев оценки реакции организма на физическую нагрузку.
В современном спорте высоких достижений биохимический контроль состояния атлета является практически обязательной составной частью процесса его подготовки. Объектами биохимических исследований обычно являются кровь, моча, реже – выдыхаемый воздух, пот и слюна. Сдвиги, вызываемые физической нагрузкой, наиболее отчетливо выявляются при анализе крови, но этот биосубстрат далеко не всегда доступен. Отсюда существует повышенный интерес к другим биологическим жидкостям таким как слюна и пот.
Преимущества возможности замены в качестве тестобъекта крови на слюну трудно переоценить, учитывая простоту и доступность ее получения и возможность мониторинга как во время тренировки, так и на соревнованиях.
При этом анализ литературы обнаружил отсутствие достоверной информации о зависимости химического состава и свойств слюны от характера нагрузок и уровня тренированности. Не выработан также единый методический подход к сбору и хранению проб слюны. Поэтому одной из задач настоящего исследования являлась разработка подобного унифицированного метода основанном на биолюминесцентном анализе.
1.1 Биохимия слюны
Слюна является одной из важнейших жидкостей организма и представляет собой прозрачную бесцветную жидкость. Слюна выделяется в полость рта тремя парами крупных слюнных желез (подчелюстные, околоушные, подъязычные) и множеством мелких слюнных желез полости рта. В полости рта образуется смешанная слюна или ротовая жидкость, состав которой отличается от состава смеси секретов желез, так как в ротовой жидкости присутствуют микроорганизмы и продукты их жизнедеятельности и различные компоненты пищи, компоненты зубного налета и зубного камня[i].
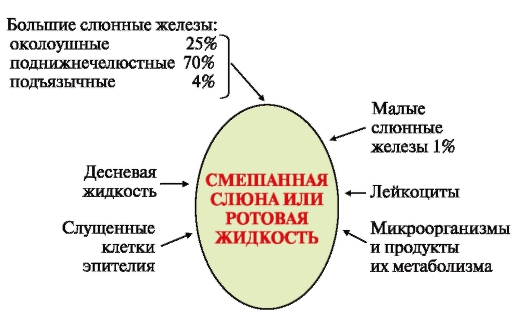
Рисунок 1 – Состав смешанной слюны
Секреция слюны носит циркадный характер – изменяется в течение суток. Средняя скорость не стимулированной секреции слюны составляет ночью во время сна – 0,05-0,1 мл/мин, днем – 0,3-0,4 мл/мин. Во время приема пищи (стимулированная секреция) резко увеличивается до 2-7 мл/мин. Таким образом, суточный объем секретируемой слюны составляет от 600 до 1200мл.
Плотность слюны колеблется в широких пределах и составляет от 1.001 до 1.017.
рН слюны в норме колеблется от 6,5 до 7,4 и зависит от характера пищи, скорости секреции и гигиены полости рта. При низкой секреции рН сдвигается в кислую сторону, при высокой – в щелочную.
Слюна выполняет ряд важных функций:
- принимает участие в очищении полости рта от остатков пищи, налета и бактерий; нейтрализует кислоты и щелочи; обеспечивает поступление ионов, необходимых для поддержания структуры гидроксиапатитов эмали; поддерживает видовой состав микрофлоры полости рта, формирует защитный барьер из муцина, железосодержащих белков и лейкоцитов; проявляет противобактериальные, противогрибковые и противовирусные свойства; участвует в образовании пелликулы зубов, предотвращает осаждение из слюны перенасыщенного раствора фосфата кальция; смачивает и размягчает твердую пищу, способствуя формированию пищевого комка, обеспечивает первый этап гидролиза крахмала пищи; регулирует образование пищеварительных соков в желудочно-кишечном тракте [ii].
Слюна имеет сложный состав. Около 99,5% слюны приходится на воду. Остальное на плотный осадок органического и неорганического происхождения.
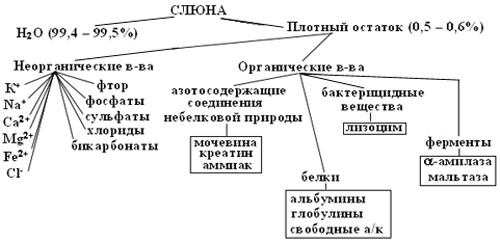
Рисунок 2 – Компоненты слюны
Среди органических компонентов слюны наиболее важными являются разнообразные белки (альбумины, глобулины, муцин, иммуноглобулины, ферменты), липиды (холестерол и его эфиры, свободные жирные кислоты, глицеролипиды), углеводы (моно - и дисахариды, свободные гликозаминогликаны), небелковые азотсодержащие вещества, витамины, циклические нуклеотиды и другие соединения.
Большую часть органических компонентов слюны составляют белковые соединения. Концентрация белка в слюне околоушной железы выше, чем в подчелюстной. Слюна содержит те же белковые фракции, что и сыворотка крови – альбумины, б-, в-, г-глобулины. Однако среди них значительно меньше альбуминов и в 4 раза больше в-глобулинов.
В состав слюны входит большое количество ферментов которые выполняют как пищеварительную, так и защитную функции.
Среди минеральных компонентов слюны преобладают натрий, калий, кальций и магний. Из всех неорганических веществ слюны наиболее важны ионы кальция и неорганического фосфата, т. к. они участвуют в минерализации зубов. Ионы кальция в слюне находятся в двух состояниях – свободном ионизированном и связанном с белками или органическими кислотами. Фосфор в слюне также находится в ионизированной форме – НРО42- и Н2РО4- и в составе белков и других органических соединений[iii,iv].
Слюна содержит три буферных системы: бикарбонатную, фосфатную и белковую.
Основной буферной системой является бикарбонатная, представленная слабой угольной кислотой – донором протонов и ионом бикарбоната – акцептором протонов. Данная буферная система наиболее эффективна при рН 6.1-6.3.
Фосфатная буферная система представлена донором протона – ионом дигидрофосфата и акцептором – ионом моногидрофосфата. Наиболее эффективна при рН 6.8-7.0.
Белковая буферная система представлена кислотными и основными группами радикалов аминокислот, входящих в состав всех белков слюны[2].
1.2 Изменение состава слюны при физических нагрузках
В последние годы проведено большое количество исследований состояния иммунной системы спортсменов в процессе физических нагрузок с использованием слюны в качестве тест-объекта. Так как проявляется преимущество легкой доступности биологической жидкости и возможность получать тест-объект в процессе самой физической нагрузки [v,vi]. Было выявлено существенное изменение секреции иммуноглобулина А (IgA) у бегунов на разные дистанции, у футболистов, у теннисистов, при силовых упражнениях. У участников сверхмарафона (160 км) сначала наблюдалось снижение секреции IgA, причем у 25% супермарафонцев этот пониженный уровень сохранялся в течении 2-х недель. Также уровень IgA в слюне оказался различным в команде пловцов и даже в какой-то мере коррелировал с уровнем их физической подготовки. Отмечается определенное достоверное снижение количества IgA в слюне велосипедистов при нагрузке, с последующим возвращением к норме. Оказалось, что принятие кофеина перед интенсивной нагрузкой вызывает повышенную секрецию IgA во время тренировки. Исследование компонентов иммуной системы оказалось полезным при выявлении наличия в организме алкоголя, а также в процессе допингконтроля у скаковых лошадей[vii,viii,ix] .
Достаточно подробно изучено циклическое изменение спектра стероидных гормонов в слюне регбистов, волейболистов, гандболистов и дзюдоистов как во время соревнования, так и в процессе недельного восстановительного периода. Уровень гормонов в слюне явился тестом в анализе психологического состояния хоккеистов при играх дома и на выезде ("родные стены помогают")[5,x].
Достаточно широко представлены в литературе изменения у спортсменов других биохимических показателей слюны. Здесь наибольший интерес представляет обнаруженная высокая степень корреляции между содержанием лактата в слюне и в крови испытуемых во время бега на различные дистанции (от 400 м до 30 км), во время теннисных соревнований, а также при нагрузках высокой мощности. Важным показателем состояния организма спортсмена во время марафонского забега являются такие биохимические показатели слюны, как минеральный состав, содержание гексозамина и свободной сиаловой кислоты, активность амилазы и пероксидазы. Определение активности амилазы, являющейся одним из основных ферментов слюны, часто проводится при спортивно-биохимическом тестировании. Отмечено, что активность антиоксидантных ферментов слюны (супероксиддисмутазы, пероксидазы, каталазы) коррелирует с уровнем свободных сиаловых кислот в слюне. Также в литературе имеются данные о повышении содержания мочевины в слюне тяжелоатлетов после стандартной тренировки[6].
2.1 Биолюминесцентная реакция, катализируемая бактериальной люциферазой
Биолюминесценция in vivo и in vitro нашла широкое применение в качестве аналитических методов и биотестов для решения проблем охраны окружающей среды, в научных исследованиях, в образовании. Биолюминесцентные методы используются для анализа метаболитов, ключевых ферментов, токсинов, мутагенов и других веществ, воздействующих на живые организмы.
Химической основой свечения бактерий является ферментативное окисление люциферазами восстановленного флавинмононуклеотида ФМН⋅Н2 и длинноцепочечного алифатического альдегида RCHO кислородом воздуха. Все бактериальные люциферазы – флавин-зависимые монооксигеназы. Все они представляют собой фермент бв-гетеродимер, состоящий из двух субъединиц, б и в, молекулярная масса которых соответственно 40 000 и 35 000. Индивидуальные субъеденицы неактивны. Эти две субъединицы гомологичны, но активный центр фермента расположен главным образом на б-субъединице.
Суммарное уравнение процесса может быть записано так:
![]() (1)
(1)
где ФМН – флавинмононуклеотид;
RCOOH – карбоновая кислота.
Ни один из исходных реагентов реакции не может существовать в бактериальной клетке в свободном виде т. к. ФМН⋅Н2 подвергается быстрому автоокислению, тетрадеканаль является ядом и не производится организмами. Поэтому бактерии имеют специальные ферментативные системы, способствующие восстановлению ФМН и карбоновой кислоты исключительно для нужд биолюминесценции. Восстановление ФМН в бактериях происходит в реакции, катализируемой НAД(Ф)H:ФMН-оксидоредуктазой:
![]() (2)
(2)
Время, требуемое для одного каталитического цикла моноферментной биолюминесцентной системы намного больше, чем время жизни одного из субстратов реакции - ФМН∙Н2, который автокаталитически окисляется кислородом менее чем за 1 сек:
ФМН∙Н2 + O2 → ФМН + H2O2 , (3)
Поэтому в условиях запуска реакции одной порцией предварительно восстановленного ФМН фермент успевает совершить всего один оборот. Наблюдается люминесцентная вспышка, затухающая по экспоненте реакция протекает в нестационарном режиме.
Восстановление ФМН можно осуществлять и химически с помощью восстанавливающего реагента НАДН. В этом случае в системе на протяжении довольно большого интервала времени присутствует ФМН∙Н2, в результате чего поддерживается стационарное свечение.
Второй субстрат – RCHO подвержен медленному неферментативному окислению, и скорость окисления зависит от температуры и начальной концентрации. При комнатной температуре раствор альдегида, используемый для измерения биолюминесценции, стабилен в течение 8 часов. Неферментативное окисление альдегида в отличие от ФМН∙Н2 не оказывает влияния на ход люминесцентной реакции, поскольку его скорость значительно меньше скорости ферментативного окисления. Все бактериальные люциферазы проявляют биолюминесцентную активность с альдегидами, длина цепи которых от восьми до шестнадцати углеродов. Существует предположение, что сродство альдегида к люциферазе обусловлено гидрофобными взаимодействиями между каждым участком алифатической цепи альдегида и гидрофобными группами фермента. Поэтому с увеличением длины углеродной цепи альдегид прочнее связывается с люциферазой. Это обеспечивает бульшую эффективность превращения химической энергии в световую. Однако эту гипотезу нельзя считать всеобъемлющей, поскольку не для всех люцифераз соблюдается монотонная связь параметров биолюминесцентной реакции с длиной цепи альдегида. Специфичность люцифераз к альдегидам проявляется в том, что другие алифатические длинноцепочечные соединения (кетоны, кислоты, спирты) не обнаруживают с люциферазой биолюминесцентной активности, хотя не исключено, что они реагируют с ней без излучения. Природным субстратом бактериальной люциферазы считается тетрадеканаль, поскольку в бактериях ферментативная система, восстанавливающая для нужд биолюминесценции карбоновую кислоту, имеет специфичность именно к миристиновой кислоте.
2.2 Кинетические особенности функционирования биферментной системы НАДН:ФМН-оксидоредуктаза-люцифераза
Любые ферменты, катализирующие синтез субстратов люциферазы, могут образовывать с ней сопряженную ферментную систему. Одной из таких является сопряженная ферментная система NAD(P)H:FMN-оксидоредуктаза-люцифераза, осуществляющая следующую цепь ферментативных реакций:
FMN + NAD(P)H → FMN H2 + NAD(P) (4)
FMN Н2 + RCHO + O2 ![]() FMN + RCOOH + Н2О + hн, (5)
FMN + RCOOH + Н2О + hн, (5)
В результате первой реакции, катализируемой NADH:FMN-оксидоредуктазой, происходит восстановление FMN с помощью восстанавливающего реагента NADН.
Вторая реакция, катализируемая люциферазой, является биолюминесцентной. В этой реакции восстановленный флавин и алифатический альдегид окисляются кислородом воздуха. В результате реакции образуется окисленная форма флавина, жирная кислота, а также испускается квант света. При проведении биолюминесцентной реакции с использованием химически восстановленного FМNН2 наблюдается длительное свечение, обусловленное множественными оборотами фермента[xi].
Список литературы
i Durdiakova J., Fabryova H., Koborova I., Ostatnikova D., The effects of saliva collection, handling and storage on salivary testosterone measurement // Steroids 2013. №14. P. 1325-1331.
ii , Биохимия органов полости рта : учеб. пособие для вузов. Полтава: из-во Полтава, 2008. C. 2-21.
iii H. Zauber, S. Mosler, A. Hessberg, W. Schulze Dynamics of salivary proteins and metabolites during extreme endurance sports – a case study // Proteomics 2012. № 12. P. 2221-2235.
iv Вавилова тканей и жидкостей полости рта: учеб. пособие / – 2-е изд., испр. и доп., 2008. 208с.
v , Михайлов как объект биохимического контроля в спорте // Ученые записки университета имени . 2008. №6(40). С. 57-61.
vi Хаустова функциональных резервов организма спортсменов различной специализации на основе анализа состава слюны: дис. … канд. биол. наук: 14.00.51. М., 2012. 167с.
vii Li T., Lin H., Ko M., Chang C., Fang S. Effects of prolonged intensive training on the resting levels of salivary immunoglobulin A and cortisol in adolescent volleyball players // The Journal of Sports Medicine and Physical Fitness. 2012. №5. Р. 569-573.
viii , , Краюшкина местного иммунитета глотки у спортсменов в норме и при хроническом воспалении небных миндалин // Международный журнал прикладных и фундаментальных исследований. 2012. №3. С. 19-20.
ix A. Tanner, B. Nielsen, J. Allgrove Salivary and plasma cortisol and testosterone responses to interval and tempo runs and a bodyweight-only circuit session in endurance-trained men // Jornal of Sports Sciences. 2013.
x R. Toone, O. Peacock, A. Smith. D. Thompson, S. Drawer Measurement of steroid hormones in saliva: Effect of sample storage condition // Scandinavian Journal of Clinical & Laboratory Investigation. 2013. № 73. Р. 615-621.
xi Суковатая методы исследования биологических процессов. 2. Определение кинетических параметров и типов взаимодействия ферментов с эффекторам: метод. указания / , . – Красноярск: Сибирский федеральный университет. 2007. 25с.
