
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Энергетика и продуктивность экосистем
После того как пища съедена, заключенная в ней энергия направляется в организме по ряду различных путей (рис. 7). Пища не переваривается и не ассимилируется вся целиком. Большинство животных не может переварить шерсть, перья, скелеты насекомых, хрящи и кости, содержащиеся в животной пище, целлюлозу и лигнин растительной пищи. Эти материалы либо выводятся наружу в экскрементах, либо отрыгиваются в виде непереваренных остатков. Некоторые из выделяемых непереваренных остатков, проходя по организму, остаются почти неизмененными по химическому составу, хотя почти все они подвергаются механическому разрушению при пережевывании пищи, что облегчает их потребление детритоядными организмами.
Организмы используют бульшую часть ассимилируемой с пищей энергии для удовлетворения своих потребностей: для выполнения разнообразной работы, для роста и размножения. Ввиду низкой эффективности биологического преобразования энергии значительная её часть, получаемая в результате метаболизма, не используется, а теряется, рассеиваясь в виде тепла. В этом отношении живые организмы мало отличаются от машин, созданных человеком. Например, в автомобильном двигателе бульшая часть заключённой в бензине энергии теряется в виде тепла, а не превращается в энергию движения.
В естественных сообществах энергия, использованная на совершение работы или рассеянная в виде тепла, не может быть потреблена другими организмами и совершенно утрачивается для экосистемы.
Ассимилированная энергия, которая не теряется в процессах дыхания и выделения, может быть использована для синтеза новой биомассы в результате роста и размножения. Популяции теряют некоторую часть своей биомассы вследствие гибели особей, болезней или ежегодного опада листвы; эта биомасса включается затем в детритные участки пищевой цепи. Остальная биомасса, в конечном счёте, потребляется растительноядными или хищниками, и заключённая в ней энергия передается на следующий, более высокий трофический уровень.
Таким образом, трофические цепи и сети в экосистемах одновременно являются и энергетическими цепями и могут быть представлены как упорядоченный процесс передачи энергии Солнца растениям — продуцентам и далее всем последующим звеньям консументов. Упрощённая схема передачи энергии в простой трофической цепи (рис. 5) представлена на рис. 8 [9].
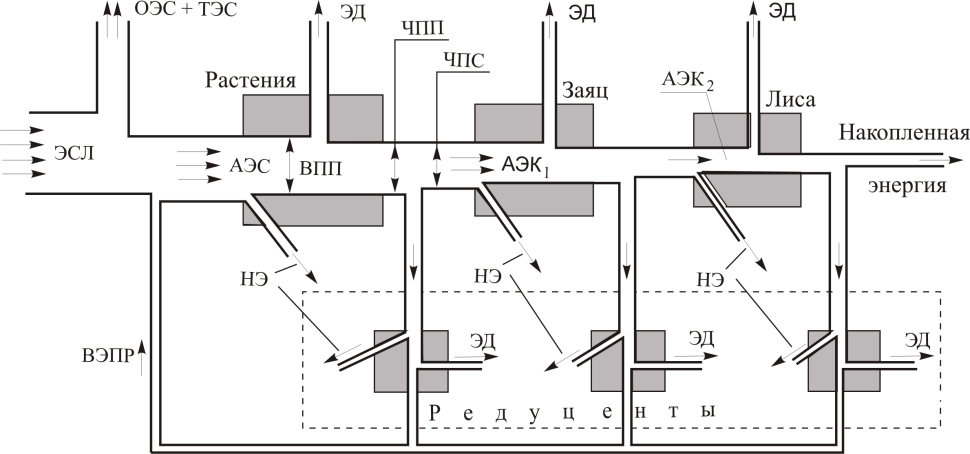
Рис. 8. Упрощённая схема потоков энергии в простой пищевой цепи
ЭСЛ — энергия солнца; ОЭС — отраженная энергия солнца; ТЭС — тепловая энергия солнца; АЭС — ассимилированная (поглощённая) энергия солнца; АЭК1,2 — энергия, ассимилированная консументами различных уровней; ЭД — энергия дыхания (теряется в виде тепла и недоступна на следующих трофических уровнях: это может быть энергия, расходуемая на процессы дыхания, кровообращения, передвижения, добычи пищи т. п.); НЭ — неиспользуемая энергия (например, часть света проходит через растения, не усваиваясь, часть пищи проходит через пищеварительный тракт, не подвергаясь энергетическим превращениям, теплоотдача и т. д.); ВЭПР — возвращаемая продуцентам энергия продуктов разложения в почве в результате жизнедеятельности организмов-редуцентов; ВПП — валовая первичная продуктивность экосистемы; ЧПП — чистая первичная продуктивность экосистемы; ЧПС — чистая первичная продуктивность сообщества
Движение энергии в экосистемах происходит в двух взаимосвязанных трофических сетях: пастбищной и детритной. В пастбищной трофической сети живые растения поедаются фитофагами (консументами 1-го уровня), а сами фитофаги являются пищей для хищников и паразитов. В детритных трофических цепях отходы жизнедеятельности живых организмов и их трупы разлагаются микроорганизмами-детритофагами до простых соединений, которые вновь используются растениями, а затем часть накопленной в организмах детритофагов энергии передаётся хищникам детритофагов (остаток энергии рассеивается в виде тепла).
При движении питательных веществ от одного трофического уровня к другому в пределах трофической цепи совершается работа и в окружающую среду выделяется низкокачественная тепловая энергия, а количество высококачественной энергии (например, химической энергии пищи), используемой организмами следующего трофического уровня, снижается. Такое снижение используемой на каждом следующем трофическом уровне энергии является следствием второго начала термодинамики.
Количество высококачественной энергии, передаваемой на следующий трофический уровень, колеблется от 2 до 30 % в зависимости от типа экосистемы и видов организмов, вовлечённых в пищевую сеть. В дикой природе, по оценкам экологов, около 10 % доступной высококачественной энергии одного трофического уровня трансформируется в доступную химическую энергию для организмов следующего трофического уровня. Оставшаяся энергия используется на поддержание процессов жизнедеятельности в организмах (пищеварение, кровообращение, передвижение, добыча и поедание пищи и т. п.), но бьльшая часть её теряется в окружающей среде в виде тепла. Редуцентами используется лишь незначительная часть энергии, проходящей по пищевой сети экосистемы, и также частично рассеивается в виде тепла. Физика утверждает, что любое вещество эквивалентно некоторому количеству энергии (формула Эйнштейна E = m∙c2). В частности, 1 г сухого органического вещества растений в среднем эквивалентен 23,5 кДж, поэтому энергию, ассимилированную растениями или животными (консументами), можно считать эквивалентной их биомассе. Простой пример на рис. 6 показывает, что в экосистемах имеет место непрерывный поток энергии, заключающийся в её передаче от одного пищевого уровня к другому. Согласно второму началу термодинамики этот процесс сопровождается рассеиванием энергии на каждом последующем звене (возрастание энтропии). Ясно, что рассеиваемая энергия всё время компенсируется энергией Солнца. В процессе функционирования экосистемы постоянно создаётся и расходуется органическое вещество. В связи с этим имеет место понятие продуктивности экологических систем.
Под продуктивностью экосистемы понимают скорость образования биомассы.
Основная или первичная продуктивность экосистемы или любой её части определяется как скорость, с которой энергия Солнца усваивается организмами-продуцентами (растениями) в процессе фотосинтеза или хемосинтеза. В процессе производства органического вещества в экосистемах выделяют четыре уровня.
Валовая первичная продуктивность — это общая скорость фотосинтеза, создающего любые разновидности органического вещества, включая органику, расходуемую на дыхание.
Чистая первичная продуктивность — скорость накопления органического вещества в растениях за вычетом вещества, использованного на дыхание.
Чистая продуктивность сообщества — скорость накопления органического вещества, не потребленного гетеротрофами, т. е. чистая первичная продуктивность минус потребление гетеротрофов.
Вторичная продуктивность — скорость накопления органического вещества на уровнях консументов.
Консументы тоже создают органическое вещество, но сами синтезировать его из неорганического вещества не могут, поэтому их продуктивность называется вторичной, независимо от их порядкового уровня. В среднем 1 га леса воспринимает 2,1∙109 кДж солнечной энергии, но если сжечь всё запасённое лесом за год органическое вещество, то в результате высвободится 1,1∙106 кДж или 0,5 % от полученной дозы солнечной энергии. Это значит, что первичная продуктивность растений не превышает 0,5 %. При передаче от одного звена трофической цепи к последующему теряется 90–99 % энергии. Например, для получения 1 кг говядины требуется 70–90 кг свежей травы.
В любой экологической системе создание биомассы осуществляется только растениями-продуцентами. Все остальные организмы только потребляют уже созданное растениями органическое вещество, и, следовательно, общая продуктивность экологической системы от них не зависит.
Так как физически энергия и масса эквивалентны, то продуктивность можно оценивать не только в энергетических единицах, но и в единицах биомассы. Продуктивность экологических систем часто выражают в виде пирамид, отражающих соотношение продуктивности различных трофических уровней. Пример экологической пирамиды [9] приведён на рис. 9. Экологические пирамиды используют в практических расчётах при обосновании, например, наиболее выгодных районов, отводимых под кормовые сельскохозяйственные культуры и обеспечивающих наибольшую продуктивность скота.

Рис. 9. Экологическая пирамида пищевой цепи:
«люцерна — теленок — человек»
В зелёных тканях растений осуществляются два противоположных процесса — фотосинтез и дыхание. При фотосинтезе создаётся органическое вещество и накапливается энергия (потенциальная), а при дыхании часть накопленного органического вещества и энергии, наоборот, расходуется. Очевидно, если в экосистеме процессы накопления вещества преобладают над процессами дыхания, то происходит накопление биомассы и энергии. Например, в водных экосистемах с глубиной уменьшается количество поступающего солнечного света и соответственно процесс фотосинтеза ослабевает и на некоторой глубине уравнивается с процессом дыхания. На больших глубинах уже доминируют процессы дыхания, т. е. происходит не накопление биомассы, а преимущественно её потребление.
Продуктивность наземных экосистем быстро уменьшается по мере перемещения от влажных тропиков к умеренным областям, особенно резко это снижение выражено в тех местах, где климат слишком сухой или слишком холодный для того, чтобы могли расти леса. Экосистемы болот и низменных побережий морей, затопляемых высокими приливами и ветровыми нагонами воды — маршей находятся на грани между наземными и водными местообитаниями, а по продуктивности растений соответствуют тропическим лесам. Максимальные уровни продукции для маршей достигают в год 4000 г/м2 в умеренных областях и 7000 г/м2 в тропиках. Растения, обитающие на маршах, высокопродуктивны, т. к. их корни постоянно находятся под водой, а листья — на свету и в воздухе; тем самым эти растения одновременно пользуются благами как водной, так и наземной сред. Кроме того, они в изобилии снабжаются питательными веществами, потому что смываемый в марши детрит быстро разлагается бактериями.
Продуктивность возделываемых земель обычно несколько ниже продуктивности естественной растительности в той же области, т. к. поля ежегодно перепахиваются и бывают оголены в начале и в конце вегетационного сезона, в то время как естественные ненарушенные местообитания продолжают создавать продукцию.
Продуктивность водных экосистем. Открытый океан — это, в сущности, пустыня, где недостаток питательных минеральных веществ (но не воды) ограничивает продуктивность до 10 % или даже менее продуктивности лесов умеренной зоны. В зонах подъёма вод из глубины к поверхности водоёма (апвеллинг), где питательные вещества выносятся на поверхность из глубины моря вертикальными течениями, и континентального шельфа, где происходит активный обмен между донными осадками и поверхностными водами, продукция выше, составляя в среднем соответственно 500 и 360 г/м2 в год. Продукция мелководных эстуариев, коралловых рифов и прибрежных зарослей водорослей приближается к продукции соседних наземных местообитаний, достигая в среднем 2000 г/м2 в год. Первичная продукция пресноводных местообитаний сходна с таковой аналогичных морских местообитаний.
Различия в продуктивности водных экосистем в значительной степени определяются доступностью питательных веществ. У отдельных растений интенсивность фотосинтеза может быть подавлена низкими температурами, тем не менее, морские водоросли достигают в холодной воде такой значительной плотности, что арктические моря столь же продуктивны, как и тёплые тропические моря.
