
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
1 U = (1*10-6М)/60 с = 1,67*10-8 М с-1 = 1, 67 * 10-8кат = 16,7 нкат.
Часто определяют удельную активность препаратов фермента делением активности навески препарата фермента, выраженной в (U), на массу навески в миллиграммах:
Ауд = U/масса препарата (мг)
При очистке ферментов удельная активность увеличивается. По возрастанию удельной активности можно судить об эффективности стадий очистки и чистоте ферментного препарата.
Для оценки активности высокоочищенных, гомогенных препаратов ферментов делением числа международных единиц (U) фермента в образце на количество вещества фермента (мкмоль) в этом образце рассчитывают молярную активность (число оборотов). По физическому смыслу молярная активность - это число молекул субстрата, подвергающихся превращению на одной молекуле фермента за 1 минуту или за 1секунду. Например: для уреазы, ускоряющей гидролиз мочевины, молярная активность составляет 30000, трипсина - 102, глюкозоксидазы - 17000 циклов в секунду.
4. Свойства ферментов
4.1. Механизм действия. Ферменты не смещают равновесие катализируемых реакций в сторону образования продуктов, таким образом, константа равновесия реакции остается постоянной. Как и все катализаторы, ферменты лишь уменьшают время достижения этого равновесия. В большинстве случаев ферменты ускоряют реакции в 107 - 1014 раз. В основе эффективности ферментативного катализа лежит сильное снижение энергии активации реакции за счет превращения субстрата в продукт через переходные состояния.
4.2. Специфичность действия. Специфичность связывания с субстратом и пути протекания ферментативной реакции определяются апоферментом. Специфичность действия ферментов определяет направленный обмен веществ в организме.
О ферментах говорят, что они имеют узкую субстратную специфичность, если они действуют на очень небольшой круг субстратов. Иногда можно говорить об абсолютной субстратной специфичности, например, каталаза катализирует только одну реакцию - разложение пероксида водорода:
![]()
Для большинства ферментов характерна относительная (широкая, групповая) субстратная специфичность, когда они катализируют группу однотипных реакций. Например, алкогольдегидрогеназа катализирует превращения спиртов в альдегиды, причем в качестве субстратов могут выступать метанол, этанол, пропанол и другие спирты. Интересным является тот факт, что алкогольдегидрогеназа может окислять и нелинейные спирты, а также спиртовую группу, входящую в состав сложных молекул, в частности, этот фермент может катализировать превращение ретинола в ретиналь. Естественно, ферменты, наделенные широкой субстратной специфичностью, катализируют превращения субстратов с различной эффективностью.
Ферменты наделены также стереохимической специфичностью: их активный центр распознает молекулы субстратов по пространственной конфигурации. Например, оксидазы L-аминокислот активны только в отношении L-аминокислот и совершенно не действуют на их D-аналоги. Для окислительного дезаминирования D-аминокислот в живых организмах имеются оксидазы D-аминокислот, не действующие на L-аминокислоты. Именно способность активного центра связываться с определенными стереоизомерами субстрата лежит в основе функционирования таких ферментов, как рацемазы, которые превращают одни стереоизомеры в другие.
Специфичность путей превращения заключается в том, что один субстрат под действием разных ферментов может превращаться в продукты, различающиеся по структуре и роли в метаболизме.
Приведем пример: оксидазы L-аминокислот действуют на L-аминокислоты, превращая их в альфа-кетокислоты с образованием аммиака и пероксида водорода.
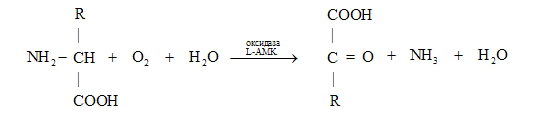
Декарбоксилазы L-аминокислот связываются с теми же субстратами, но катализируют другую реакцию: декарбоксилирование с образованием биогенных аминов и выделением углекислого газа.
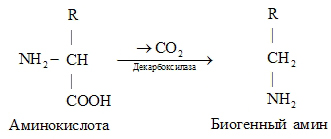
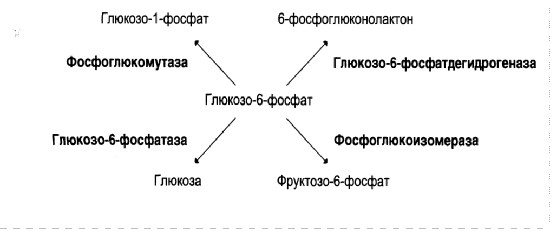
Еще одним примером является возможность превращения глюкозо-6 фосфата под действием различных ферментов, по одному из возможных метаболических путей:

4.3. Термолабильность.
Как и многие белки, при повышении температуры ферменты подвергаются термической денатурации, что приводит к нарушению нативной конформации фермента и изменению структуры активного центра. Ферменты млекопитающих начинают заметно денатурировать при температурах выше 40оС.
В связи с вышесказанным, ферментные препараты желательно хранить при пониженных температурах. Одним из лучших путей сохранения ферментов является их лиофилизация (высушивание при температуре ниже -70оС в вакууме), переведение в частично денатурированное состояние с помощью солей аммония и помещение в холодильник.
4.4. Зависимость скорости реакции от температуры. Скорость ферментативных реакций, как и любых химических реакций, зависит от температуры. При повышении температуры на 10оС скорость реакции увеличивается в 2-4 раза согласно правилу Вант-Гоффа. Однако при температурах выше 40оС существенной становится денатурация ферментов, что приводит к уменьшению суммарной активности (рис. 2):
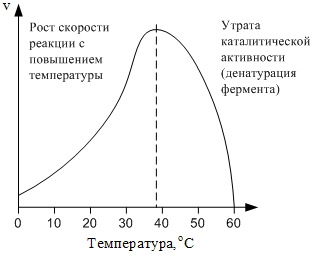
Рис. 2. Зависимость скорости ферментативной реакции от температуры.
4.5. Зависимость скорости реакции от рН. Зависимость скорости ферментативной реакции от рН имеет колоколообразный вид (рис. 3). Значения рН, при которых наблюдается наиболее высокая скорость ферментативной реакции, называют оптимальными (рН-оптимум). Характер кривых и значение рН-оптимума зависит от природы заряженных групп субстрата и заряженных групп фермента (особенно тех, которые входят в активный центр). Оптимум рН для большинства ферментов лежит в пределах от 6,0 до 8,0 (рис. 3).
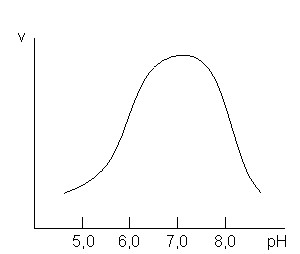
Рис. 3. Зависимость скорости ферментативной реакции от рН.
Однако, есть и исключения, например, пепсин наиболее активен при рН 1,5 – 2,0, а щелочная фосфатаза при рН 10,0 – 10,5 (рис. 4)
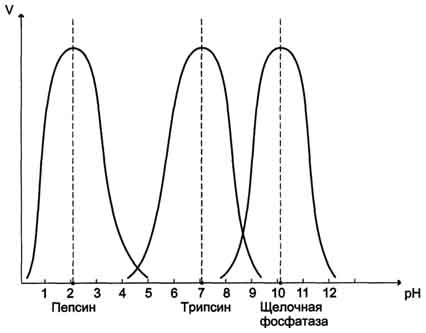
Рис. 4. Зависимости скорости ферментативной реакции (v) от рН среды.
При экстремальных (очень низких или очень высоких) значениях рН происходит нарушение третичной структуры молекулы фермента, приводящее к потере ферментативной активности.
5. Кинетика ферментативных реакций
Кинетика ферментативных реакций изучает скорости катализируемых ферментами реакций и влияние на них различных факторов.
В простейшем случае ферментативную реакцию можно представить как двухстадийный процесс. К настоящему времени многочисленными экспериментами показано, что первая стадия - это образование фермент-субстратного комплекса (ЕS). Вторая стадия - это превращение фермент-субстратного комплекса через ряд промежуточных состояний в комплекс фермент-продукт и распад этого комплекса на свободный фермент и продукт реакции. Наиболее коротким схематическим способом записи этих стадий является:
![]()
Зависимость скорости реакции v от концентрации субстрата Сs описывается уравнением Михаэлиса-Ментен, где Км (константа Михаэлиса), а Vмакс (max) - максимальная скорость ферментативной реакции при полном насыщении фермента субстратом:
Графически зависимость скорости реакции от концентрации субстрата представляет полуветвь гиперболы (рис.5):
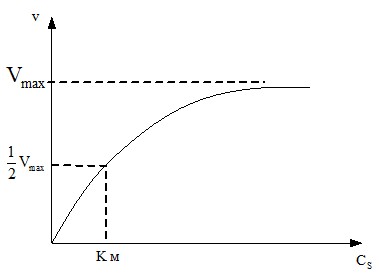
Рис. 5. Зависимость скорости реакции от концентрации субстрата.
При полном насыщении фермента субстратом наблюдается ситуация, когда Cs >> Kм, тогда значением Км в знаменателе можно пренебречь, и скорость реакции становится максимальной:
v = Vмакс
На графике (рис.5) при этом наблюдается плато при высоких концентрациях субстрата.
При полунасыщении фермента субстратом v = Vмакс/2. Подстановка этого значения скорости в уравнение Михаэлиса приводит к значению Км = Cs. Поэтому можно дать следующее определение Км: константа Михаэлиса - это концентрация субстрата, при которой скорость ферментативной реакции равна половине максимальной скорости.
Поскольку в ходе ферментативной реакции наблюдается расходование субстрата и накопление продукта, зависимость их концентраций от времени можно выразить следующим образом (рис. 7):
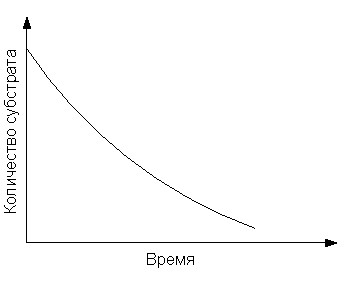
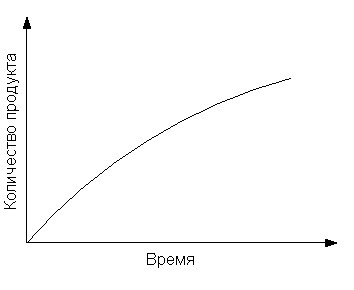
Рис. 7. Графики зависимости концентрации субстрата и продукта ферментативной реакции от времени.
Для более удобного графического представления экспериментальных данных Г. Лайнуивер и Д. Бэрк предложили использовать двойные обратные величины 1/v и 1/Cs, поскольку если существует равенство между двумя какими-либо величинами, то и обратные величины также равны.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
