

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Основные аспекты использования молочнокислых бактерий
в молочной промышленности:
Часть 1. Мониторинг штаммов молочнокислых бактерий на
способность к образованию низкомолекулярных полипептидов
Одегов Нинолай Иванович, к. т.н., ст. научный сотрудник,
, к. т.н., ст. научный сотрудник,
, к. т.н., ст. научный сотрудник, зав. лабораторией
ГНУ Сибирский научно-исследовательский институт сыроделия Россельхозакадемии, г. Барнаул
Поскольку молочная отрасль занимает одно из ведущих мест в переработке сельскохозяйственной продукции, то важным будет рассмотреть применение различных штаммов молочнокислых бактерий в процессе производства в молочной промышленности, в частности для проектирования пищи с функциональными ингредиентами геродиетического назначения. Это является возможным благодаря гидролизу молочных белков до низкомолекулярных полипептидов под воздействием протеолитических ферментных систем молочнокислых бактерий.
Функциональные продукты, к которым в частности относятся геродиетические, при систематическом употреблении стимулируют жизнедеятельность организма в целом или оказывают определенное регулирующее действие на определенные системы, органы или функции организма.
Анализ ситуации промышленного производства функциональных продуктов в России показывает, что в настоящее время в их ассортиментной структуре недостаточно специализированных геродиетических продуктов. Большое значение при этом имеет их биологическая и пищевая ценность, характеризуемая наличием и рациональным содержанием основных ингредиентов, в частности незаменимых аминокислот, насыщенных жирных кислот, витаминов, минеральных веществ и др. [1].
Следует отметить, что молоко и, как следствие, молочные продукты, изготовляемые из молочного сырья по традиционным технологиям, не полностью удовлетворяют требованиям геродиетики, в частности, по липидному и белковому составам. Кроме того, для получения высококачественных продуктов в т. ч. и геродиетических необходимо использовать лишь кондиционное сырье.
В соответствии с современными требованиями геронтологии содержание алиментарного белка в организме человека должно быть повышено до 1,00 – 1,25 г на 1 кг массы тела. При этом необходимо сочетание животных и растительных белков, имеющих в своем составе достаточное количество низкомолекулярных полипептидов (в т. ч. олигопептидов).
Олигопептиды являются эффективными биорегуляторами. Они, напрямую воздействуя на метаболизм клетки, повышают ее толерантность к гипоксии, действию токсинов и иных негативных факторов, нормализуют процессы усвоения тканями питательных веществ и выведения продуктов обмена, позитивно влияют на многие функции организма в нормальных и патологических условиях [2].
Одной из главных задач при создании геропродутов является проблема обеспечения их биологической и пищевой ценности, характеризуемой наличием и обоснованным содержанием нутриентов, в частности, белковых структур различной иерархии сложности (полипептидов, олигопептидов и аминокислот), ненасыщенных жирных кислот, витаминов, минеральных веществ, пребиотиков и др.
При этом спектр и доля содержания этих ингредиентов должны адекватно соответствовать возрастным изменениям метаболизма и потребностям организма пожилых людей. Нормализация липидно-белкового состава геропродуктов обычно осуществляется за счет использования соответствующих биологически активных веществ (БАВ), получаемых различными способами.
В частности, в качестве белковых структурных обогатителей применяются гидролизаты растительной и животной этиологии.
Однако такая коррекция химического состава путем механического добавления каких-либо компонентов требует предварительного осуществления специальных технологических операций (например, ферментация, ультрафильтрация, структурная дифференциация и др.). Это затрудняет процесс получения корректирующих БАВ.
При этом сам процесс гидролиза белковых молекул технологически сложно контролировать вследствие труднорегулируемой активности специфических пептидаз, неопределенности уровня деструкции и выхода пептидов с заданной аминокислотной цепью.
Предполагается, что перспективным способом обогащения молочных продуктов олигопептидами является контролируемая ферментация молочных белков с помощью комплекса протеаз ферментных препаратов и специально селектированных микроорганизмов, применяемых в биотехнологиях молочных продуктов, например в сыроделии.
Особое место занимает продукция молочной отрасли, обладающая высокой биологической и пищевой ценностью, особенно ее сыродельной промышленности.
Известно, что сыр, как продукт направленной биоконверсии молочных ингредиентов, обладает определенной геродиетической ценностью. В процессе созревания осуществляется гидролиз казеина под воздействием молокосвертывающих ферментов с образованием пептидов, которые в свою очередь гидролизуются пептидазами заквасочной микрофлоры. Кроме того, под воздействием липолитических ферментов происходит частичное гидролитическое расщепление основных липидных фракций в сыре с образованием диацилглицеринов и моноацилглицеринов, жирных кислот и др. продуктов реакции. Такое «упрощение» снижает порог энергетической и ферментативной доступности нутриентов для их метаболизации в организме пожилого человека. Последнее и определяет уровень героадекватности пищевых продуктов [3, 4, 5].
Литические системы ферментирующих бактерий также осуществляют неспецифический гидролиз пептидов, как нативной этиологии, так уже и частично деструктурированных химозином и пепсином.
Таким образом, можно сделать предположение о том, что суммарный спектр продуктов протеолиза МФП является определителем матрицы ферментного отклика клеток заквасочных культур, используемых в данной биотехнологии.
Естественно предположить, что установление параметров этой корреляции позволит сделать процесс протеолиза прогнозируемым и управляемым как по «глубине», так и по «ширине».
Реализация этого предположения возможна путем применения в биотехнологиях молочных продуктов определенных заявленных композиций МФП и заквасочных культур, обеспечивающих оптимальное соотношение в протеиназно-пептидазном литическом пуле агентов, как МФП - так и бактериальной этиологии. Такое соотношение, в свою очередь, предопределяет требуемую точность дозировки глубины, интенсивности и направленности процесса протеолиза, заданную степень деструкции белковой молекулы до уровня олигопептидов и достижение высокого содержания последних в готовом продукте.
Метод проведения исследований. Для определения в исследуемых субстратах, обработанных протеолитическими ферментами микробного происхождения, концентрации низкомолекулярных полипептидов (НМПП) с молекулярной массой (ММ) менее 16 Kd, использовали оригинальный мембранный метод. Метод основан на выделении фракций полипептидов с различной ММ способом ультрафильтрации (УФ) прямым спектрофотометрированием при л = 216 нм).
Материалы: Ячейки для УФ с отсечкой по ММ = 3,0; 5,0; 10,0 кДа ("VivaScience", США), Спектрофотометр СФ-46 (Россия), Центрифуга лабораторная напольная ОС-6м, производства Росси (диаметр ротора 28 см, угловая скорость не менее 18000 об/мин), Центрифуга Type MPW-310, производства Польши (диаметр ротора 15 см, угловая скорость не менее 15000 об/мин), обеззоленные бумажные фильтры (белая лента, d = 11 см), протеазы молочнокислых бактерий, казеин по Гаммерстену, ч (по ТУ 6-09-3574-82), в качестве субстрата использовали восстановленное обезжиренное молоко.
Методика получения гидролизатов
Выделение НМПП методом УФ. В качестве субстрата применяли восстановленное обезжиренное молоко, причем с целью устранения возможных колебаний его состава, в течение всего периода исследований использовали одну и туже партию сухого молока. В исследуемый субстрат объемом 100 мл, вносили 1,0 М НСl до рН=4,6, перемешивали в течение 10 мин и центрифугировали 15 минут при v = 2,5 тыс. об./мин (диаметр ротора 28 см, угловая скорость не менее 18000 об/мин). Супернатант (0,5 мл) вносили в ячейку для УФ и центрифугировали в течение 30 мин при v = 12 тыс. об./мин (диаметр ротора 15 см, угловая скорость не менее 15000 об/мин). Для дальнейшего исследования использовали пермеат.
Определение концентрации НМПП биуретовым методом. 0,1 мл пермеата или фильтрата доводили до 10 мл 3%-ным раствором NaOH. Полученную смесь в объеме 4 см3 переносили в пробирку (опыт). В качестве контроля использовали 4 мл раствора NaOH 3%. В контрольную и опытную пробу добавляли по 0,2 мл реактива Бенедикта, тщательно перемешивали, выдерживали 15 минут при комнатной температуре и спектрофотометрировали при л=330 нм. Концентрацию НМПП, в мг%, определяли по калибровочному графику.
Определение концентрации НМПП прямым спектрофотометрированием. Определяли оптическую плотность пермеата при л=216 нм. Концентрацию НМПП, в мг%, определяли по калибровочному графику.
Для построения калибровочного графика, из стандартного щелочного раствора казеина готовили ряд растворов с известной концентрацией белка: 2,5; 12,5; 20,0; 37,5; 50,0 мг в 100 мл раствора NaOH 3%, с таким расчетом, чтобы искомое содержание НМПП входило в диапазон, охваченный вышеназванным рядом.
При определении концентрации НМПП, необходимо учитывать все разведения, которые претерпевает анализируемая проба в ходе всего процесса тестирования.
Результаты исследований. В работе использовали 62 производственно-ценных культуры мезофильных молочнокислых стрептококов (лактококков) видов Lactococcus lactis subsp. cremoris (Lc. cremoris), Lactococcus lactis subsp. lactis (Lc. lactis), Lactococcus lactis subsp. lactis biovar. diacetylactis (Lc. diacetylactis), Lactobacterium plantarum (L. plantarum), Lactobacterium lactis (L. lactis), Lactobacterium acidophilus (L. acidophilus), Lactobacterium helveticus (L. helveticus) и Streptococcus termophilus (S. termophilus) из коллекций лаборатории биохимических и микробилогических исследований и биофабрики.
В качестве объекта для последующей ферментации (субстрата) применяли восстановленное обезжиренное молоко.
Исследовались 10-суточные молочнокислые культуры тестируемых штаммов.
Установлено, что межштаммовые различия в интенсивности продуцирования НМПП были наибольшими для культур вида Lc. cremoris. Коэффициент вариации массовой доли прироста НМПП для этого вида лактококков составил 218%.
Для культур Lc. diacetylactis данный коэффициент был значительно меньше и составил 136%.
Термофильные виды молочнокислой микрофлоры в порядке увеличения прироста продуцирования НМПП с ММ менее 16 Kd располагались в следующем порядке S. thermophilus (диапазон прироста от 46 мг% до 116 мг%), L. lactis (диапазон прироста от 112 мг% до 308 мг%), L. helveticus (диапазон прироста от 218 мг% до 364 мг%) и L. acidophilus (диапазон прироста от 328 мг% до 372 мг%). При этом в таком же порядке возрастала и массовая доля данных НМПП в общем растворимом белке. Диапазоны изменения ее величины составляли от 53,9 % до59,5 %; от 79,4 % до90,0 %; от 78,7 % до 92,1 и от 95,0 % до99,5 % соответственно вышеназванным видам.
Внутривидовая дифференциация уровня прироста ПП с ММ менее 10 Кd иллюстрируется рисунками 1 и 2.
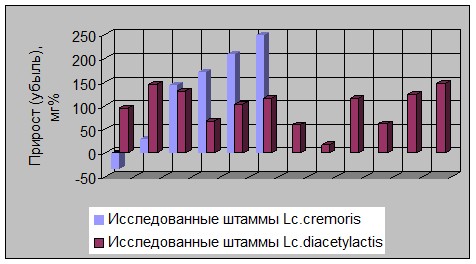
Рисунок 1 - Изменение прироста содержания полипептидов разной ММ
в молочных субстратах в зависимости от вида ферментирующих мезофильных
молочнокислых бактерий
Из гистограммы (см. рис 1) видно, что штаммы вида Lc. cremoris были более вариабельны по уровню данного прироста, чем культуры Lc. diacetylactis.
Межштаммовые различия по этому признаку для термофильных видов были наиболее выраженными для культур L. helveticus и L. lactis, а наименее - для S. thermophilus (см. рис. 2).
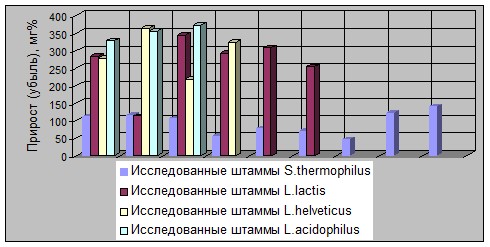
Рисунок 2 - Изменение прироста содержания полипептидов разной ММ
в молочных субстратах в зависимости от вида ферментирующих термофильных
молочнокислых бактерий
Выявлен факт о более высоком приросте содержания низкомолекулярных полипептидов в субстратах, ферментированных культурами L. helveticus и L. acidophilus по сравнению с лактобактериями.
Полученные в вышеназванных «биуретовых» экспериментах мониторинговые данные по исследованным видам свидетельствуют о достаточно высокой протеолитической активности тестированных культур молочнокислых лактококков и палочек в направлении образования полипептидов.
Данные экспериментов по применению «мембранного метода разделения фракций ПП» представлены в таблице 1.
Таблица 1 - Содержание общего растворимого азота и полипептидов с разной ММ в молочных субстратах, ферментированных молочнокислыми бактериями
№ п/п | Се-рия | Штамм молочнокислых бактерий | Прирост (убыль) содержания полипептидов, мг% | |||
общего азота растворимого | поли-пептидов с ММ10 Kd | поли-пептидов с ММ5 Kd | поли-пептидов с ММ3 Kd | |||
1 | 1 | Контроль (содержание) | 0 (866) | 0 (413) | 0 (360) | 0 (326) |
2 | 1 | S. thermophilus 125/4 | -3 | 103 | 105 | 120 |
3 | 1 | S. thermophilus 32/ар | 28 | 112 | 155 | 64 |
4 | 1 | S. thermophilus 33д | 34 | 109 | 150 | 143 |
5 | 1 | L. lactis 11-Е | 109 | 228 | 263 | 293 |
6 | 1 | L. lactis 2512 | 53 | 225 | 263 | 293 |
7 | 1 | L. lactis 1638 | 371 | 468 | 309 | 280 |
8 | 1 | L. lactis 126 | 84 | 198 | 203 | 207 |
9 | 1 | L. lactis 2970 | 459 | 506 | 459 | 312 |
10 | 1 | L. helveticus 18-13 | 472 | 650 | 553 | 349 |
11 | 2 | Контроль (содержание) | 0 (1050) | 0 (488) | 0 (425) | 0 (338) |
12 | 2 | L. helveticus 6014 | 50 | 112 | 75 | 150 |
13 | 2 | L. acidophilus 6/3 | 80 | 205 | 263 | 308 |
14 | 2 | L. acidophilus 3c | 110 | 449 | 475 | 412 |
15 | 2 | L. acidophilus 20т | 150 | 512 | 500 | 582 |
16 | 2 | L. casei 21ст | 112 | 250 | 238 | 135 |
17 | 2 | L. plantarum 23ст | -10 | 6 | 56 | 122 |
18 | 2 | Lc. lactis 593/4 | 0 | 108 | 166 | 181 |
19 | 2 | Lc. lactis 833/7 | 60 | 288 | 68 | 137 |
20 | 2 | Lc. lactis 996/4-3 | 40 | 215 | 200 | 252 |
Как свидетельствуют результаты (см. табл. 1), массовая доля низкомолекулярных ПП с разной ММ (3, 5 и 10 Kd) в общем растворимом белке составляла для наиболее репрезентативных выборок видов S. thermophilus, L. lactis, Lc. lactis и L. acidophilus, соответственно, от 43% до 58%; от 48% до 71%; от 42% до 64% и от 57% до 83%. Необходимо отметить, что данные диапазоны перекрываются, однако тенденция увеличения рассматриваемой доли в «палочковых» гидролизатах по сравнению с «кокковыми» просматривается вполне отчетливо.
На рисунках 3, 4, 5 и 6 графически представлена дифференциация исследованных штаммов 4 видов молочнокислых бактерий по величине доли содержания ПП разной ММ в общем растворимом азоте.
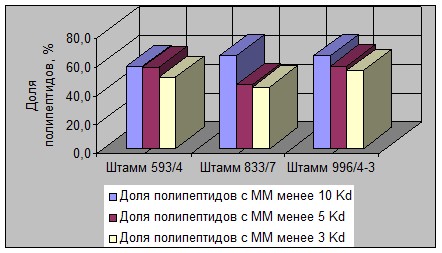
Рисунок 3 - Изменение массовой доли ПП разной ММ в растворимом белке
в гидролизатах штаммов Lc. lactis
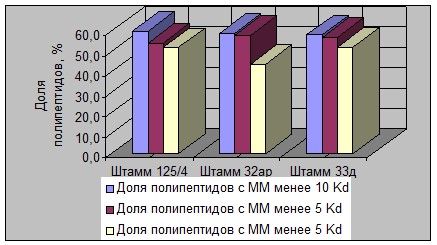
Рисунок 4 - Изменение массовой доли ПП разной ММ в растворимом белке
в гидролизатах штаммов S. thermophilus
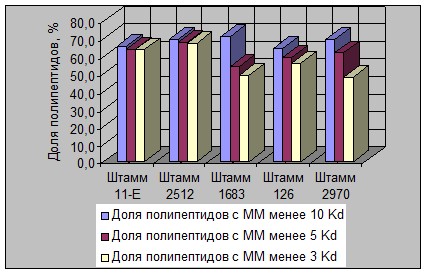
Рисунок 5 - Изменение массовой доли ПП разной ММ в растворимом белке
в гидролизатах штаммов L. lactis
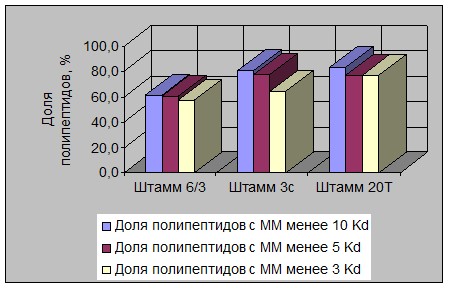
Рисунок 6 - Изменение массовой доли ПП разной ММ в растворимом
белке в гидролизатах штаммов L. acidophilus
Таким образом, по виду гистограмм можно косвенно судить о параметрах процесса гидролиза белков, осуществляемого конкретным штаммом.
В связи с этим надо отметить, что ряд штаммов палочковых видов молочнокислых бактерий характеризовался большей долей накопления ПП фракции «ММ ≤ 3 Kd» (например штаммы L. lactis 11, L. lactis 2512, L. acidophilus 6/3, L. acidophilus 20Т и др.).
Полученные результаты вполне согласуются с литературными данными о большей активности палочек в отношении накопления более «легких» продуктов гидролиза белков по сравнению с кокковыми формами лактобактерий [4, 5].
Примерно такой же вывод можно сделать и по данным, усредненным по видовым выборкам исследованных культур, которые представлены в таблице 2.
Таблица 2 - Усредненные показатели уровни прироста полипептидов (ПП) с различной молекулярной массой в молочных субстратах, ферментированных различными видами молочнокислых бактерий
Вид ПП | Показатель | Lc. lactis | S. termophilus | L. lactis | L. acidophilus |
Общий раствор. азот | Средний прирост; мг/% | 33,3 | 19,7 | 215,2 | 113,3 |
Диапазон варьиро-вания; мг/% | От 0 до 60 | 0т -3 до 34 | От 53 до 459 | От 80 до 150 | |
Коэффициент. вариации; % | 180,2 | 187,8 | 188,7 | 61,8 | |
ППс ММ ≤ 10 Kd | Средний прирост; мг/% | 203,7 | 108,0 | 325,0 | 388,7 |
Диапазон варьиро-вания; мг/% | От 108 до 288 | От 103 до 112 | От 198 до 506 | От 205 до 512 | |
Коэффициент вариации; % | 88,4 | 8,3 | 94,8 | 79,0 | |
ПП с ММ ≤ 5 Kd | Средний прирост; мг/% | 144,7 | 136,7 | 299,4 | 412,7 |
Диапазон варьиро-вания; мг/% | От 68 до 200 | От 105 до 155 | От 203 до 459 | От 263 до 500 | |
Коэффициент вариации; % | 91,2 | 36,6 | 85,5 | 57,4 | |
ПП с ММ ≤ 3 Kd | Средний прирост; мг/% | 190,0 | 109,0 | 277,0 | 434,0 |
Диапазон варьиро-вания; мг/% | От 137 до 252 | От 64 до 143 | От 207 до 312 | От 308 до 582 | |
Коэффициент вариации; % | 60,5 | 72,5 | 37,9 | 63,1 |
Как свидетельствуют данные (см. таблицу 2), средний (по изученным выборкам культур) прирост выхода ПП с ММ ≤ 10 Kd составил 108,0мг%; 325,0мг%; 203,7мг%; и 388,7мг%, а коэффициент вариации уровня прироста – 8,3%; 94,8%; 88% и 79 %, соответственно видам S. thermophilus, L. lactis, Lc. lactis и L. acidophilus. Видно, что кокки «отставали» по этому приросту от палочек.
Динамика изменения усредненного содержания ПП для вышеотмеченных видов молочнокислых бактерий в осях «уровень прироста» - «молекулярная масса» представлена на рисунке 7.
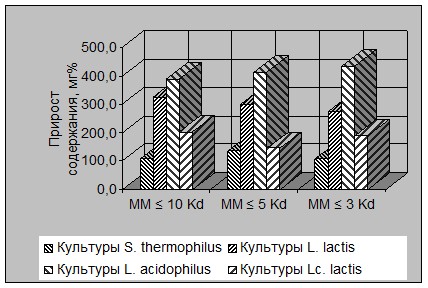
Рисунок 7 - Изменение прироста содержания полипептидов разной ММ в молочных
субстратах в зависимости от вида ферментирующих молочнокислых бактерий
Как свидетельствуют данные (см. рис. 7), гистограммы для всех 4 видов бактерий имеют выраженную ступенчатость для каждого исследованного значения ММ., что говорит об уровне межвидовых различий по средним значениям прироста ПП с разной ММ.
При этом межвидовая линия тренда практически идентична для точек «ММ ≤ 3 Kd» «ММ ≤ 5 Kd» и «ММ ≤ 10 Kd», что свидетельствует о сохранении характера этих различий для всех фракций ПП.
Таким образом, по результатам данных тестов наименее вариабельны по уровню прироста выхода низкомолекулярных ПП являются (из числа изученных) штаммы S. thermophilus, но они и менее продуктивны по рассматриваемым показателям (уровню прироста и содержанию низкомолекулярных ПП). Наибольший средний прирост выхода ПП с ММ менее 10 Kd был выявлен для штаммов L. acidophilus, несколько меньший последних - для L. lactis. Культуры Lc. lactis характеризовались промежуточным среди рассматриваемых видов средним уровнем прироста полипептидов данной молекулярной массы.
Резюмируя обсуждение результатов исследований, можно выделить ряд культур наиболее продуктивных по приросту низкомолекулярных ПП разной ММ.
Например, с точки зрения прироста выхода наиболее интересной в контексте данной НИР фракции «ММ ≤ 10 Kd» к таковым относятся штаммы L. lactis 2970 (прирост 506 мг%); L. lactis (468 мг%); L. helveticus 18-30 (650 мг%); L. acidophilus 20Т (512 мг%); L. acidophilus 3с (449 мг%); Lc. lactis 833/7 (288 и Lc. lactis 996/4-3 (215 мг%).
Для фракции «ММ ≤ 5 Kd» это будут штаммы S. thermophilus 32ар (155 мг%); S. thermophilus 33д (150 мг%); L. lactis 2970 (459 мг%); L. lactis 1638 (309 мг%); L. acidophilus 20Т (500 мг%); L. acidophilus 3с (475 мг%); Lc. lactis 996/4-3 (200 мг%); Lc. lactis 593/4 (166 мг%).
И, наконец, для фракции «ММ ≤ 3 Kd» - S. thermophilus 33д (143 мг%); S. thermophilus 125/4 (120 мг%); L. lactis 2970 (312 мг%); L. lactis 2512 (293 мг%); L. acidophilus 20Т (582 мг%); L. acidophilus 3с (412 мг%); Lc. lactis 996/4-3 (252 и Lc. lactis 593/4 (181 мг%).
На вышеуказанных «мембранных» гистограммах не представлены важные с точки зрения сыроделия культуры вида L. helveticus (вследствие малой репрезентативности выборки). В связи с этим необходимо отметить, что штамм L. helveticus 18-13 обеспечивал прирост выхода всех трех фракций низкомолекулярных ПП на уровне самого продуктивного из всех тестированных культур штамма L. acidophilus 20Т, уступая последнему лишь в продуцировании фракции «ММ ≤ 3 Kd» (в 1,7 раза).
Напротив, другой штамм L. helveticus 6014 давал очень низкий прирост по всем фракциям. Данный прирост составлял 112 мг%; 75 мг%; и 150 мг% соответственно фракциям «ММ ≤ 10 Kd», «ММ ≤ 5 Kd» и «ММ ≤ 3 Kd», т. е. был намного ниже, чем практически у всех исследованных штаммов (за исключением S. thermophilus 125/4 и L. plantarum 23ст, также не представленного на этих гистограммах).
Известно, что направленность и глубина протеолитических процессов в рассматриваемом случае определяется в первую очередь геномом штамма. Естественно, что его фенотипическая реализация детерминируется конкретными условиями ферментации (составом субстрата, наличия нативных протеиназ молока эндо - и экзоферментов остаточной микрофлоры и др.).
ВЫВОДЫ
В молочной промышленности большое значение имеет применение молочнокислых бактерий, поскольку от уровня молочнокислого брожения и активности ферментных систем заквасочных культур в целом, зависит качество изготавливаемого продукта.
1. Установлено, что межштаммовые различия в интенсивности продуцирования олигопептидов были наибольшими для культур вида Lc. сremoris.
2. Термофильные виды молочнокислой микрофлоры в порядке увеличения прироста продуцирования олигопептидов с ММ менее 16 Kd располагались в следующем порядке: S. thermophilus (диапазон прироста от 46 мг% до 116 мг%), L. lactis (диапазон прироста от 112 мг% до 308 мг%), L. helveticus (диапазон прироста от 218 мг% до 364 мг%) и L. acidophilus (диапазон прироста от 328 мг% до 372 мг%).
3. Штаммы S. thermophilus менее продуктивны по уровню прироста и содержанию низкомолекулярных полипептидов. Наибольший средний прирост выхода полипептидов с ММ менее 10 Kd был выявлен для штаммов L. acidophilus, несколько меньший последних - для L. lactis. Культуры Lc. lactis характеризовались промежуточным среди рассматриваемых видов средним уровнем прироста полипептидов данной молекулярной массы.
4. Выявлен факт о более высоком приросте содержания низкомолекулярных полипептидов в субстратах, ферментированных культурами L. helveticus и L. acidophilus по сравнению с кокковыми культурами.
5. В целом, по результатам исследований культуры вида L. lactis детерминировали наиболее широкий по спектру и достаточно глубокий протеолиз, а культуры вида L. acidophilus отличались наибольшей интенсивностью и глубиной этого процесса.
Проведенные исследования позволяют для разных условий ферментации выбрать среди тестированных культур виды и штаммы, наиболее перспективные с точки зрения увеличения выхода низкомолекулярных полипептидов.
Для обеспечения такого мягкого управления процессом протеолиза предлагается использование, в частности, эндопротеазной способности селектированных молочнокислых бактерий и применение их в биотехнологиях молочных продуктов.
Список используемой литературы:
1. Гаврилова продукт для функционального питания / , // Сыроделие и маслоделие. – 2011. - №6. - С. 32-33.
2. Замятнин. Блистающий мир белков и пептидов. (http: // www. inbi. ras. ru/, http: // erop. inbi. ras. ru /.
3. Белов активность молочнокислых бактерий применяемых при выработке сыров / // Сборник материалов V специализированного конгресса «Молочная промышленность Сибири». – Барнаул, 2006. – с.58-30.
4. Белов аспекты протеолитических процессов при производстве сыров, Актуальные проблемы техники и технологии переработки молока: сборник трудов. Вып. 3 / ГНУ Сибирский НИИ сыроделия СО РАСХН. Барнаул, 2006. С. 76-86
5. Шергина технологии в области сыроделия / , , // Сыроделие и маслоделие. - 2008. - №2. – С. 14-15.
Реферат статьи:
Использование продуктов специфического гидролиза молочных белков для проектирования пищи с функциональными ингредиентами геродиетического назначения:
Часть 1. Мониторинг штаммов молочнокислых бактерий на
способность к образованию низкомолекулярных полипептидов
, к. т.н., ст. научный сотрудник,
, к. т.н., ст. научный сотрудник,
, к. т.н., ст. научный сотрудник, зав. лабораторией
ГНУ Сибирский научно-исследовательский институт сыроделия Россельхозакадемии, г. Барнаул
Ключевые слова: гидролиз молочных белков, проектирование пищи, функциональные ингредиенты, геродиетическое назначение, низкомолекулярные полипептиды, мониторинг молочнокислых бактерий
Главная задача при создании геропродутов - обеспечение их биологической и пищевой ценности, характеризуемой наличием олигопептидов и аминокислот, витаминов и др.
Перспективный способ обогащения молочных продуктов олигопептидами - ферментация молочных белков протеазами ферментных препаратов и селектированных микроорганизмов.
Исследовали интенсивность продуцирования низкомолекулярных полипептидов различными культурами молочнокислых бактерий. Проведенные исследования позволяют выбрать виды и штаммы культур, перспективные с точки зрения увеличения выхода низкомолекулярных полипептидов.
