

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
5. Моделирование процесса эвтрофирования водоемов
Решение проблемы антропогенного эвтрофирования водоемов невозможно без глубокого знания закономерностей функционирования их экосистем. Все природные экосистемы очень сложны, а когда приходится иметь дело с высокоорганизованными системами, то суждения и интуиция могут приводить к ошибочным решениям и необратимому экологическому ущербу.
Достаточное количество наблюдений для перехода от вполне оформившихся качественных представлений о механизме эвтрофирования к попыткам количественной его оценки было накоплено к концу 60-ых годов нынешнего столетия. Таким образом, была создана необходимая основа для разработки математических моделей эвтрофирования [Modelling…, 1974].
Специфика всех моделей эвтрофирования, с формальной точки зрения, заключается в попытках адекватно описать связи изменения запаса в воде биогенных веществ с трофическим состоянием водоема, отражаемым, в первую очередь, величиной его первичной продукции. На этом пути условно выделяют две группы моделей: качественно-аналитические и эмпирические; имитационные (портретные)
Для первой группы характерно:
- небольшое количество моделируемых параметров состояния, объединяющих характеристики со сходной структурой, таксономию или другие характеристики. Примером может служить количественная характеристика общего фосфора (
,
Робщ), , встречающегося в природных и сточных водах, в виде суммарной массы элемента фосфора во всех его формах, или биомассы многочисленных видов фитопланктона по концентрации хлорофилла “а”(Chl. a) в нефильтрованной воде водного объекта. (В лимнологической литературе стало общепринятым обозначать содержание хлорофилла “а” — символом Chl. a.).
- небольшое число элементарных связей, что вытекает из объединения параметров,
- исследование средних для водоема стационарных условий протекания процесса эвтрофирования.
Вторая группа моделей предназначена для воспроизведения взаимосвязанных изменений отдельных форм элемента. Эти модели детализированы, многокомпонентны и требуют больших объемов исходной информации.
5.1. Модели экологического состояния водоемов
Попытки количественных оценок процесса эвтрофирования начались в середине ХХ века, когда первые серьезные лимнологические исследования этого процесса показали тесную зависимость трофического состояния озера от комплекса гидрологических и гидрохимических факторов [Sakamoto, 1966, Vollenweider, 1968]. Обобщив данные по сравнительно небольшому количеству североамериканских озер, Раусон показал наличие связи между биомассой фитопланткона в озере с его глубиной, а несколько позже Сакамото установил тесную зависимость содержания хлорофилла с концентрацией фосфора в японских озерах [Sakamoto, 1966]. Эти исследования предвосхитили классическую работу Фолленвейдера, разработавшего первые диаграммы для определения трофического состояния озер в зависимости от глубины и фосфорной нагрузки на озеро [Vollenweider, 1968].
Примерно в это же время для расчета средней концентрации фосфора в озерах им были применены дифференциальные балансовые уравнения, основанные на допущении полного мгновенного перемешивания воды в водоеме, при этом Фолленвейдер рассматривал скорость внутриводоемной седиментации фосфора как функцию его общего содержания в озере. Предложенная им эмпирическая зависимость коэффициента седиментации фосфора от интенсивности водообмена водоема завершила формирование достаточно стройной концепции количественного описания процесса эвтрофирования озер, основанной на стационарных балансовых моделях [Vollenweider, 1968]. Принципиальную систему связей балансовых моделей можно представить схематически следующим образом (рис. 5.1).
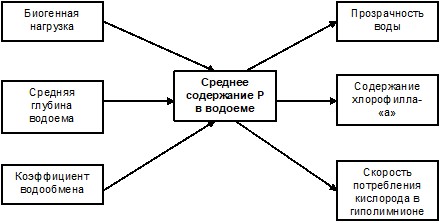
Рис. 5.1. Система связей в простых стационарных моделях эвтрофирования водоема.
Основное балансовое уравнение Фолленвейдера имеет вид
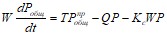 , (5.1)
, (5.1)
где 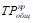 — приток общего фосфора с водосбора за определенный промежуток времени t (объемная нагрузка водоема общим фосфором), Pобщ - концентрация фосфора в водоеме, W - объем воды в водоеме,
— приток общего фосфора с водосбора за определенный промежуток времени t (объемная нагрузка водоема общим фосфором), Pобщ - концентрация фосфора в водоеме, W - объем воды в водоеме, ![]() — расход воды, вытекающей из водоема за то же время t, Kс - коэффициент седиментации (осаждения) фосфора
— расход воды, вытекающей из водоема за то же время t, Kс - коэффициент седиментации (осаждения) фосфора
Аналитическое решение этого уравнения имеет вид
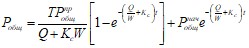 , (5.2)
, (5.2)
где ![]() - начальная концентрация общего фосфора в воде водоема.
- начальная концентрация общего фосфора в воде водоема.
При установившемся процессе
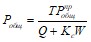
Альтернативный подход разработан П. Диллоном и Ф. Риглером [1975], использовавшим вместо коэффициента седиментации коэффициент удержания фосфора в водоеме (КР), сравнительно легко определяемый по составляющим внешнего баланса фосфора
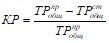 ,
,
Где ![]() — количество вытекающего с водой из озера или сбрасываемого из водохранилища общего фосфора.
— количество вытекающего с водой из озера или сбрасываемого из водохранилища общего фосфора.
Стационарная модель Диллона-Риглера имеет вид
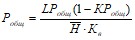
Где LPобщ – нагрузка водоема общим фосфором по площади, Н – средняя глубина, Кв – коэффициент водообмена водоема
Предложенные балансовые модели интегрально описывают комплекс процессов, определяющих удержание фосфора в водоеме и вовлечение его во внутренний круговорот. Теоретическая обоснованность их применения, подтверждаемая получаемыми адекватными результатами, основана на том, что скорость изменения результирующей внешнего баланса фосфора в водоеме замедленного водообмена, т. е. запаса общего фосфора, значительно меньше, чем скорость основных биологических процессов в ходе круговорота фосфора – первичной продукции и бактериальной деструкции (месяцы-годы по сравнению с сутками-часами). Эта особенность позволяет рассматривать задачу прогнозирования концентраций общего фосфора в водоеме независимо от оценки состояния его экосистемы при данном уровне содержания фосфора. Таким образом, стационарные модели определяют биогенный фон экосистемы и тем самым могут успешно служить для косвенных ориентировочных оценок изменения трофического состояния экосистемы. Их теоретическую обоснованность подтвердил , получив такую же как и Фолленвейдер зависимость содержания хлорофилла в озере от концентрации фосфора с помощью аналитической, но сильно детализированной экологической модели [Вавилин, 1980].
С момента появления стационарных балансовых моделей усилия исследователей процесса эвтрофирования были сосредоточены на поисках эмпирических связей коэффициента седиментации и коэффициента удержания фосфора с гидрологическими и морфометрическими характеристиками водоемов. В 70-х годах появилось большое количество работ с предложениями подобных эмпирических зависимостей, результаты которых обобщены К. Рэкхау [1979]. При этом в большинстве работ водоемы не дифференцируются по типам и не выделяются границы применимости этих зависимостей. Использование предложенных связей коэффициента седиментации от определяющих его факторов привело к многочисленным модификациям балансовой модели эвтрофирования.
В общем случае процесс седиментации фосфора зависит не только от гидрологических факторов, но также и от гидрохимического режима и состояния экосистемы водоема. В частности, содержание и изменчивость форм некоторых элементов (кальция, железа и алюминия) может оказывать прямое воздействие на величину удержания фосфора в водоеме, контролируя растворимость его соединений и процессы сорбции на минеральной взвеси. Осаждение фосфора также зависит от продукционных процессов в водоеме как прямо (сорбция фосфатов органической взвесью), так и косвенно (изменение физико-химических характеристик среды, определяющих процесс сорбции и растворимости фосфатов). Недостаточно еще изучена в континентальных водоемах и роль биоседиментации в осаждении фосфора, которая в океанах имеет исключительно важное значение [Савенко, 1988].
Большинство водохранилищ относится к долинному типу и характеризуется длинной и вытянутой формой в плане. Для описания удержания фосфора и расчета средней концентрации фосфора в таких водохранилищах Д. Хиггинсом и Б. Кимом [1981] была рассмотрена модель, основанная на гипотезе полного вытеснения, примененная ими к водохранилищам каскада Тенесси. Расчет средней концентрации общего фосфора по этой стационарной модели проводится по формуле
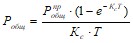
Где ![]() - концентрация общего фосфора в притоке, Кс – коэффициент седиментации общего фосфора, Т – период водообмена водоема
- концентрация общего фосфора в притоке, Кс – коэффициент седиментации общего фосфора, Т – период водообмена водоема
Коэффициенты седиментации фосфора в водохранилищах определяется с помощью коэффициентов водообмена и удержания по формуле
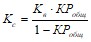
Где Кв – коэффициент водообмена водохранилища, КPобщ – коэффициент удержания общего фосфора в водохранилища.
Из дальнейших предложений по совершенствованию стационарных балансовых моделей фосфора заслуживают внимания работы Т. Фриска [1989], С. Чапры и К. Рэкхау [Chapra, Reckhow, 1983], попытавшихся подкорректировать один из главных недостатков моделей Фолленвейдера – допущение о полном мгновенном перемешивании водоема. Эта корректировка основана на введении дополнительного параметра, формально характеризующего долю объема водоема, соответствующего условиям полного перемешивания. Параметр D предлагается определять на эмпирической основе (Frisk, 1989). В этом случае средняя концентрация общего фосфора в водоеме вычисляется с учетом расчетных концентраций общего фосфора по моделям полного мгновенного перемешивания и вытеснения

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
