

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
РЕАКЦИИЭЛЕКТРОННОГО ТРАСНФЕРА В МОДИФИЦИРОВАННЫХ МУТАНТАХ ЦИТОХРОМА B562
1, 2, 1
1-Саратовский Государственный Университет
2-Chapman University, USA
*****@***ru
Работа посвящена теоретическому изучению реакций электронного переноса в ковалентно модифицированных металлопротеинах. Явление электронного трансфера (ЭТ) – неотъемлемая часть проходящих повсеместно биологических окслительно-восстановительных реакций (ОВР), которые играют важную роль в таких жизненных биохемических процессах, как фотосинтез, клеточное дыхание, репарации ДНК, биодеградации и биосинтез. Детальное изучение реакций электронного трансфера может стать важной ступенью на пути к созданию биокатодов [1] и солнечных батарей [2].
В реакциях электронного переноса на большие расстояния электронные состояния донора и акцептора взаимодействуют слабо. В таком неадибатическом режиме электронного трансфера система может достигнуть возбужденного состояния (которое характеризуется равными мгновенными энергиями связи электронов донора и акцептора) намного раньше, чем произойдет сам электронный трансфер. В таком случае скорость электронного переноса может быть вычислена по формуле Маркуса
![]()
![]() (1)
(1)
где ![]()
![]() – эффективное взаимодействие донор/акцептор, или электронный каплинг,
– эффективное взаимодействие донор/акцептор, или электронный каплинг, ![]()
![]() – свободная энергия обмена реакции электронного трансфера,
– свободная энергия обмена реакции электронного трансфера, ![]()
![]() – энергия реорганизации, которая характеризует ядерный отклик системы на трансфер электрона от донора к акцептору.
– энергия реорганизации, которая характеризует ядерный отклик системы на трансфер электрона от донора к акцептору.
Целью данной работы был расчет скоростей ЭТ для производных цитохрома b562 с использованием молекулярно-динамического моделирования и оригинального метода вычисления. Мы сосредоточили свое внимание на кофакторе электронных состояний гема, которые сильно влияют на топологию рассматриваемой молекулярной системы, а также на моделировании конформационной мобильности рутениевого комплекса.
Структуры цитохрома b562 были модифицированы по принципу Ru(bpy)2(Im)(HisX) поочередно для гистидинов с номерами 12, 15, 19, 63, 70, 73, 86, 89 и 92 с помощью программного пакета Harlem [2] из рентгеновских структур нативного белка.
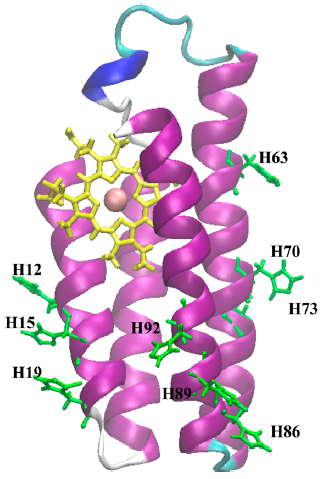
Рис. 1. Производные цитохрома b562
Рисунок 1 показывает позиции аминокислот гистидина, которые были использованы как место прикрепления Ru (2,2′-бипиридин)2(имидазол)2 окислительно-восстановительной группы (RBP). Координаты рутениевого комплекса были взяты из протеиновой базы данных 1BEX для рутениум-модифицированного азурина [3]. Модифицированные структуры белка были помещены в раствор, состоящий из около 9000 молекул воды. Ионы Na+ были добавлены, чтобы нейтрализовать систему. Вначале была проведена минимизация энергии системы, после которой была запущена равновесная молекулярная динамика в течение 1 нс.
Мы вычислили значения донор/акцептор каплингов для 100 молекулярно-динамических точек каждого мутанта b562. С этой целью мы сократили белковый фрагмент, используя процедуру PATHWAYS с критерием 1%. Фрагменты содержали 250-400 атомов. Для каждого фрагмента белка мы провели вычисления методом Хартри-Фока и посчитали донорно-акцепторные каплинги, используя метод функций Грина. С целью сравнения с экспериментальными данными [4] мы посчитали скорости неадибатического электронного переноса, используя в формуле Маркуса (1) квадрат среднего значения HDA по молекулярно-динамическим точкам. Значения ДG0 и л были взяты из экспериментальной работы [5]: −ДG0 ≈л ≈ 0.8 eV.
Сравнение вычисленных нами скоростей электронного переноса с экспериментальными можно посмотреть на Рис. 2. Мы можем увидеть отличное соответствие для мутантов His12, His15, His73, His92 и His86 в пределах множиДля мутантов His19, His63, His70 и His89 согласование можно назвать хорошим – оно находится в пределах множителя 8.
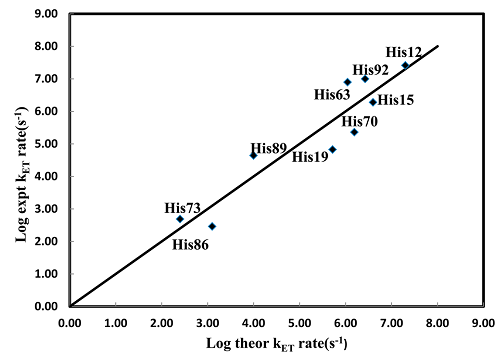
Рис. 2. Сравнение вычисленных скоростей электронного переноса с экспериментальными для девяти производных цитохрома b562
Наши вычисления показывают, что взаимодействие положительно заряженного RBP с отрицательно заряженными аминокислотами на поверхности белка играет важную роль в определении предпочтительной конформации рутениевого комплекса.
Список литературы.
Improved Interfacial Electron Transfer in Modified Bilirubin Oxidase Biocathodes. Ryan J. Lopez1, Dr. Sofia Babanova et al. ChemElectroChem Volume 1, Issue 1, Pp. 241–248, January 3, 2014 Integrating a redox-coupled dye-sensitized photoelectrode into a lithium–oxygen battery for photoassisted charging. M. Yu, Lu Ma et al. Nature Communications 5 Kurnikov, I. V. Harlem: Molecular Modeling Package; University of Pittsburg and Carnegie Mellon University: Pittsburgh, PA, 2005. Gu, J.; Yang, S.; Rajic, A. J.; Kurnikov, I. V.; Prytkova, T. R.; Pletneva, E. V. Control of cytochrome c redox reactivity through off-pathway modifications in protein hydrogen-bonding network. mun. 2014, 50, 5355−5357. Gray, H. B.; Winkler, J. R. Electron tunneling through proteins. Q. Rev. Biophys. 2003, 36, 341−372. Winkler, J. R.; Di Bilio, A. J.; Farrow, N. A.; Richards, J. H.; Gray, H. B. Electron tunneling in biological molecules. Pure Appl. Chem. 1999, 71, 1753−1764