

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Предполагаемые рубрики: биохимия, иммунология, иммунохимия.
Очистка высокомолекулярной субфракции из кожи свиньи, ингибирующей пролиферацию эпидермоидных клеток карциномы человека А431
*, *, *, *, *, *, *, **, **, *, *
* – Федеральное государственное бюджетное учреждение науки «Научно-исследовательский институт физико-химической медицины» ФМБА России, директор – академик РАМН , Москва; ** - Федеральное государственное бюджетное учреждение науки «Институт биоорганической химии им. академиков и » РАН, директор – академик РАН , Москва.
Реферат
Была проведена очистка субфракции с молекулярной массой более 250 кДа, полученной из кожи свиньи и ингибирующей пролиферацию эпидермоидных клеток карциномы человека А431, методом анионообменной хроматографии на смоле ДЭАЭ 32 с использованием ступенчатого градиента концентрации NaCl. Изучали влияние исходной субфракции и фракций после разделения на ДЭАЭ 32 на пролиферацию эпидермоидных клеток карциномы человека А431 ин витро в двух тестах: МТТ-тесте и флуоресцентном тесте. С помощью более специфичного флуоресцентного теста было выявлено, что наибольшей ингибирующей активностью обладала фракция №2, которая смывается с колонки при концентрации NaCl 0,15М. Методом вертикального электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия в этой фракции был выявлен один мажорный белковый компонент и ряд минорных белковых компонентов.
Ключевые слова: кожа, высокомолекулярные белки, пролиферация, регуляция, эпидермоидные клетки, карцинома человека А431.
Адрес для корреспонденции:
119435, а.
ФГБУН НИИ ФХМ ФМБА России,
Тел.: (499) 246-43-42; (499) 246-42-96
E-mail: *****@***ru
Регуляция пролиферации клеток кожи находится под контролем множества факторов, которые оказывают как стимулирующее, так и ингибирующее действие [14,9,13,11]. Нарушение регуляторных механизмов пролиферации и апоптоза лежит в основе развития онкологических заболеваний. Показано, что один и тот же фактор по-разному влияет на пролиферацию нормальных и злокачественных клеток [8].
Ранее авторами статьи были разработаны способы получения веществ из кожи свиньи, влияющих на пролиферацию и дифференцировку клеток кожи человека [3,5,6,7]. С помощью одного из способов [3] из кожи свиньи был выделен препарат с молекулярной массой (м. м.) более 15 кДа, который стимулировал пролиферацию и дифференцировку кератиноцитов человека [4], стимулировал В-систему иммунитета [2] и ингибировал пролиферацию эпидермоидных клеток карциномы человека (ЭККЧ) А431 [1]. Из этого препарата была выделена высокомолекулярная субфракция №1 (СФ1), которая вызывала более сильное ингибирование пролиферации ЭККЧ А431 по сравнению с исходным препаратом [1].
Целью данной работы была дальнейшая очистка субфракции №1, ингибирующей пролиферацию ЭККЧ А431, методом анионообменной хроматографии на смоле ДЭАЭ 32 и характеристика очищенной фракции методом электрофореза.
Методика исследования
Из кожи свиньи с помощью метода, запатентованного ранее [3], был выделен препарат с м. м. более 15 кДа (П>15кДа). Затем он был разделен на колонке с сефадексом G-100 на четыре субфракции с различными молекулярными массами [1]. Была взята СФ1 с м. м. более 250 кДа, обессолена на колонке с сефадексом G-15 и лиофилизирована. СФ1 была использована в настоящей работе для дальнейшей очистки методом анионообменной хроматографии на колонке со смолой ДЭАЭ 32 с использованием ступенчатого градиента концентрации NaCl в 0,01М трис-НСl буфере рН 7,0. Влияние СФ1 и фракций после ДЭАЭ 32 на пролиферацию ЭККЧ А431 ин витро определялось в двух тестах: МТТ-тесте и флуоресцентном тесте.
МТТ-тест [10]. ЭККЧ A431 выращивали по методике в среде RPMI-1640 с добавлением 10 % эмбриональной телячьей сыворотки, 1мМ пирувата, 2мМ L-глутамина и 50 мкг/мл гентамицина. Клетки культивировали во флаконах, пересев осуществляли с помощью трипсина (0,25%) и ЭДТА (0,02%). Затем клетки в количестве 5х103 вносили в лунки плоскодонного 96-луночного планшета, культивировали в течение 7 ч., после чего в культуры добавляли исследуемые препараты до конечной концентрации 300, 200, 100 и 10 мкг/мл. Затем клетки культивировали еще в течение 48 часов. Пролиферацию клеток определяли по тому количеству живых клеток, которое оказывалось в культуре после культивирования, за вычетом исходного количества клеток. Оценку количества живых клеток проводили с использованием MTT-теста. Измерение оптической плотности содержимого каждой лунки проводили на приборе "Multiscan MCC/340" при длине волны 540 нм. Количество живых клеток оценивали по величине оптической плотности раствора формазана. Статистическую обработку результатов проводили по t-критерию Стьюдента.
Флуоресцентный тест [1]. ЭККЧ A431 выращивали по методике на среде DMEM с добавлением 10 % эмбриональной телячьей сыворотки и стандартным набором антибиотиков. Клетки культивировали во флаконах, пересев осуществляли с помощью трипсина (0,25%) и ЭДТА (0,02%). Определение количества клеток в процессе культивирования проводили в камере Горяева с красителем трипановым синим при одновременном подсчете процента жизнеспособных клеток. На последнем этапе аликвоты суспензии клеток в среде DMEM с сывороткой разливали в 24-луночный планшет с покровными стёклами по 300 мкл в лунку. Количество клеток в лунке было ~2х105. Клетки инкубировались в течение 7 часов, после чего в культуры добавляли исследуемые препараты до конечной концентрации 200 и 100 мкг/мл. После этого клетки культивировали в течение еще 48 часов. Окраска осуществлялась йодидом пропидия. Съёмка образцов проводилась на конфокальном флуоресцентном микроскопе Nikon Eclipse E800. Анализ количества клеток в образцах проводился в программе ImageJ 1.36. В контроле проанализировано 4 образца, в каждом опыте – по 2 образца. В каждом образце оценено в среднем около 3 тысяч клеток.
СФ1 и фракции после разделения на смоле ДЭАЭ 32 были проанализированы методом вертикального ДСН-электрофореза в 12,5% ПААГ с использованием реактивов Fisher's EZ-RunTM Protein Gel Solution, а также методом вертикального двуступенчатого ДСН-электрофореза в 6-12% ПААГ и 6-20% ПААГ по Лэммли [12]. После проведения электрофореза препараты окрашивались на белок красителем Кумасси R-350 и сканировались.
Результаты исследования
В данной работе субфракция №1 (СФ1) была разделена на 5 фракций методом анионообменной хроматографии на колонке 1,5х10 см со смолой ДЭАЭ 32 с использованием ступенчатого градиента концентрации NaCl в 0,01М трис-НСl буфере рН 7,0. Фракция №1 (0M NaCl) не адсорбировалась на колонке. Фракция №2 смывалась 0,05М NaCl. Фракция №3 смывалась 0,15М NaCl. Фракция №4 смывалась 0,2М NaCl. Фракция №5 смывалась 2М NaCl. После сбора фракции были лиофилизированы и обессолены на колонке с сефадексом G-15. Взвешивали каждую фракцию и определяли, какой процент по весу она составляет от суммарного веса всех фракций. Выход фракции №1 составил 5%, фракции №2 – 3%, фракции №3 – 28%, фракции №4 – 20% и фракции №5 -44%.
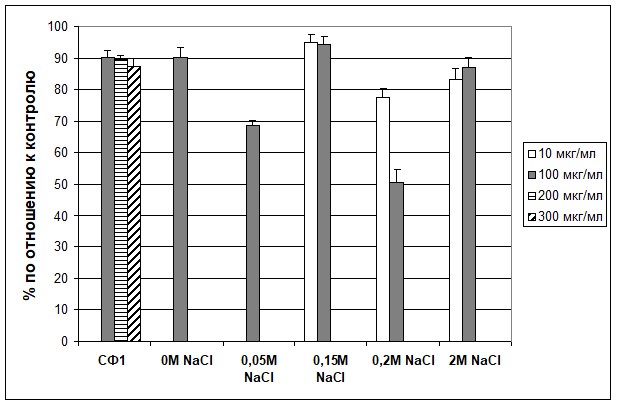
Изучали влияние СФ1 и фракций после разделения на смоле ДЭАЭ 32 на количество выросших ЭККЧ А431 в культуре с использованием МТТ-теста. Было обнаружено, что СФ1 и все фракции ингибируют рост клеток (рис. 1). Однако, наибольший эффект оказала фракция №4, смываемая с колонки 0,2M NaCl. В концентрации 100 мкг/мл она ингибировала пролиферацию на 49%. Различие с контролем достоверно: Р<0,0001.
Выход фракции №2 (0,05М NaCl) составлял всего 3%. Полученного материала было недостаточно для проведения физико-химического и иммунобиологического анализа. Поэтому было решено при следующем разделении собирать не пять, а четыре фракции: 0М NaCl (фрация №1); 0,15M NaCl (фракция №2); 0,2M NaCl (фракция №3) и 2M NaCl (фракция №4). После разделения СФ1 на колонке 2,5х30 см со смолой ДЭАЭ32 фракции также были лиофилизированы и обессолены на колонке с сефадексом G-15. Взвешивали каждую фракцию и определяли, какой процент по весу она составляет от суммарного веса всех фракций. Выход фракции №1 составил 5%, фракции №2 – 40%, фракции №3 – 24% и фракции №4 – 31%.
Затем было изучено влияние СФ1 и фракций после ДЭАЭ 32 на пролиферацию ЭККЧ А431 в культуре с использованием флуоресцентного метода. Надо отметить, что как при использовании МТТ-теста, так и флуоресцентного метода, после инкубации клеток с препаратами количество мертвых клеток не увеличивалось. Жизнеспособность клеток оставалась примерно на одном уровне. Определение жизнеспособности клеток флуоресцентным методом при инкубации с препаратами не обнаружило нарушений метаболизма клеток и структурной целостности мембран. Можно полагать, что эффект препаратов не является цитотоксическим. Клетки пролиферировали митозом.
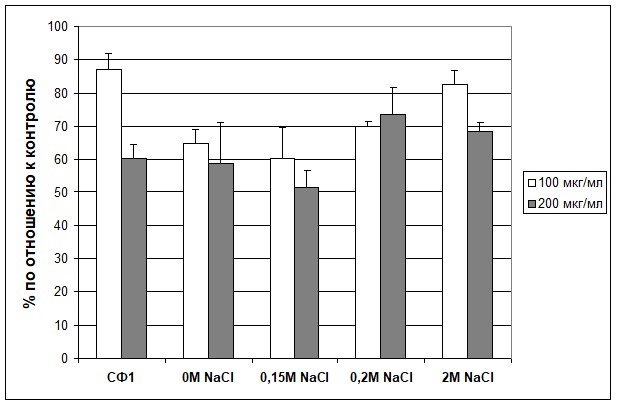
При анализе общего количества клеток в образце было обнаружено, что и исходная СФ1 и фракции после анионообменной хроматографии вызывали достоверное ингибирование пролиферации клеток А431 (рис.2). Наибольший эффект оказывала фракция №2, которая смывается с колонки при концентрации NaCl 0,15М. Она вызывала ингибирование пролиферации на 48% при концентрации белка 200 мкг/мл. Различие с контролем достоверно: Р<0,01. Анализ относительного количества колоний, в которых шла активная пролиферация (процент колоний, в которых клетки прошли 3 и более делений), также выявил максимальное и дозо-зависимое ингибирование именно этой фракцией. На этот тип анализа случайность распределения клеток по лункам влияет в меньшей степени. У остальных фракций ингибирующий эффект был менее выражен.
Относительно механизма гибели клеток надо отметить, что морфологических признаков апоптоза не наблюдалось, выраженной гибели клеток также не наблюдалось. Скорее всего, основной механизм действия препаратов связан с арестом клеточного роста, хотя механизм ингибирования пролиферации требует дальнейшего изучения.
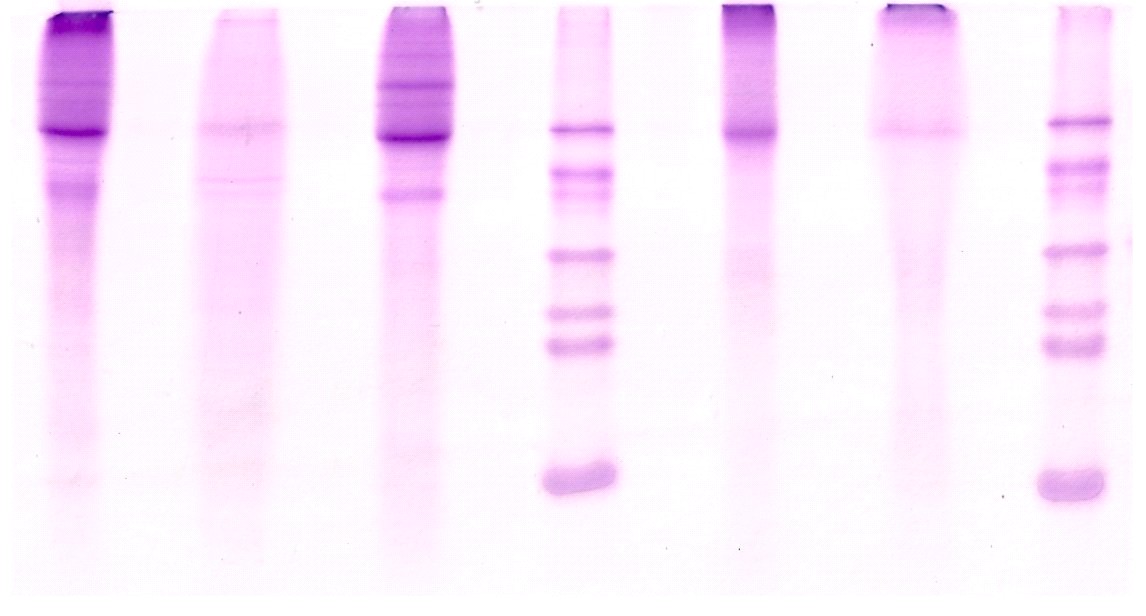
СФ1 и фракции после разделения на смоле ДЭАЭ 32 были проанализированы методом вертикального ДСН-электрофореза в 12,5% ПААГ с использованием реактивов Fisher's EZ-RunTM Protein Gel Solution. Использование EZ-Run Gel дает возможность разделять белки в большом диапазоне молекулярных масс. Гель обладает градиентно-подобными свойствами и позволяет разделять как низкомолекулярные белки с м. м. 10 кДа, так и высокомолекулярные белки с м. м. 200 кДа в одном и том же денатурирующем геле. Помимо этого препараты были проанализированы методом вертикального двуступенчатого ДСН-электрофореза по Лэммли [12] в 6-12% и 6-20% ПААГ. Окраска на белок – красителем Кумасси голубым R-350. Данные ДСН-электрофореза в 6-20% ПААГ представлены на рис. 3.
В СФ1 и во всех фракциях было выявлено по одной мажорной полосе с м. м. 63-67 кДа и ряд минорных полос. Наименее гетерогенной оказалась фракция №4 (2M NaCl). Помимо мажорной полосы, в ней выявлена лишь одна минорная полоса с м. м. 48 кДа. В остальных препаратах, помимо мажорной полосы, выявлено 4-8 минорных полос с м. м. от 25 до 50 кДа и от 92 до 237 кДа. Возможно, что мажорный компонентт с м. м. 63-67 кДа является одним и тем же белком с близкими молекулярными массами. Возможно также, что это разные белки. Во фракции №2 (0,15M NaCl), обладающей максимальной ингибирующей активностью, выявляется мажорный белковый компонент с м. м. 63 кДа, два минорных белковых компонента с м. м. 36 и 96 кДа и ряд более слабых белковых миноров с м. м. 28, 30, 47, 52, 81 и более 96 кДа.
Использование двух разновидностей метода оценки пролиферации: МТТ-теста и флуоресцентного теста дало возможность заключить, что наибольшей активностью обладали фракции, которые смываются с колонки при концентрации NaCl 0,15М и 0,2М. При сравнении результатов двух разделений можно заключить, что при первом разделении на анионообменной колонке максимальной активностью обладала фракция, которая смывалась с колонки при концентрации NaCl 0,2М; в то время как при втором разделении максимальной активностью обладала фракция, которая смывалась с колонки при концентрации NaCl 0,15М. Это может быть обусловлено несколькими причинами. Во-первых, были использованы разные методы оценки пролиферации клеток. После первого разделения влияние фракций на пролиферацию ЭККЧ А431 оценивали с использованием МТТ-теста, в то время как после второго разделения использовали флуоресцентный метод.
С помощью МТТ-теста мы определяем жизнеспособность клеток или количество жизнеспособных клеток. Он прост и дёшев и широко используется для оценки пролиферации клеток. Однако при его использовании интегрально оценивается как количество клеток, так и их метаболическая активность. Так что увеличение метаболической активности клеток может компенсироваться уменьшением их количества. Обычно этот тест используют для грубого скрининга, а затем применяют более специфические подходы в зависимости от задачи.
Флуоресцентный тест более специфичен, его обычно используют для изучения клеточного цикла. Ошибка используемого флуоресцентного метода невелика при условии, что анализируется несколько тысяч клеток. В данной работе это условие было соблюдено, причем подсчет производился автоматизировано, т. е. было устранено влияние оператора. Однако, прямого сопоставления двух методов в одних и тех же условиях мы не проводили, т. е. сравнивать их напрямую сложно. В основе визуализации этих двух тестов лежат разные субстраты, с чем и может быть связаны различия в полученных результатах.
Во-вторых, различия могут быть связаны с особенностями получения фракций. Метод получения СФ1 достаточно сложный. СФ1, полученные от выделения к выделению мало отличаются по ДСН-электрофорезу в ПААГ, но, как можно увидеть из вышеприведенных результатов, отличаются по проценту выхода ДЭАЭ-фракций. Можно предположить, что эти фракции могут также отличаться по распределению в них белков, влияющих на пролиферацию клеток.
Поскольку все фракции после разделения на ДЭАЭ 32 вызывали ингибирование пролиферации клеток, можно говорить об удалении из СФ1 менее активных компонентов в плане ингибирующего влияния на пролиферацию ЭККЧ А431. Не исключена возможность, что в СФ1 присутствует несколько активных белков, которые после разделения на ДЭАЭ-целлюлозе попадают в разные фракции. При оценке результатов исследования однозначно следует отдать предпочтение более специфичному флуоресцентному тесту.
Таким образом, была проведена очистка субфракции с молекулярной массой более 250 кДа, полученной из кожи свиньи и ингибирующей пролиферацию эпидермоидных клеток карциномы человека А431, методом анионообменной хроматографии на смоле ДЭАЭ 32 с использованием ступенчатого градиента концентрации NaCl. С помощью более специфичного флуоресцентного теста было выявлено, что наибольшей ингибирующей активностью обладала фракция №2, которая смывается с колонки при концентрации NaCl 0,15М. Методом вертикального электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия в этой фракции был выявлен один мажорный белковый компонент с м. м. 63 кДа и ряд минорных белковых компонентов.
Цифры к рисунку 1.
Значения средней арифметической:
10 мкг/мл | 100 мкг/мл | 200 мкг/мл | 300 мкг/мл |
СФ1 | 90,3 | 89,4 | 87,5 |
0M NaCl | 90,1 | ||
0,05M NaCl | 68,7 | ||
0,15M NaCl | 94,8 | 94,3 | |
0,2M NaCl | 77,6 | 50,7 | |
2M NaCl | 83,3 | 87 |
Значения ошибки средней:
10 мкг/мл | 100 мкг/мл | 200 мкг/мл | 300 мкг/мл |
СФ1 | 2 | 1,5 | 2,5 |
0М | 3,4 | ||
0,05М | 1,6 | ||
0,15М | 2,8 | 2,6 | |
0,2М | 2,9 | 3,9 | |
2М | 3,3 | 3,1 |
Цифры к рисунку 2.
Значения средней арифметической:
СФ1 | 0M NaCl | 0,15M NaCl | 0,2M NaCl | 2M NaCl | |
100 мкг/мл | 87 | 64,9 | 60,5 | 70 | 82,6 |
200 мкг/мл | 60,5 | 59 | 51,6 | 73,7 | 68,6 |
Значения ошибки средней:
СФ1 | 0M NaCl | 0,15M NaCl | 0,2M NaCl | 2M NaCl | |
100 мкг/мл | 5,0 | 4,1 | 9,1 | 1,5 | 4,3 |
200 мкг/мл | 4,0 | 12,1 | 5,1 | 8,1 | 2,5 |

Подписи к рисункам:
Рис. 1. Влияние субфракции №1 и фракций, полученных после разделения СФ1 методом анионообменной хроматографии на ДЭАЭ 32, на пролиферацию ЭККЧ А431 в культуре (МТТ-тест).
СФ1 – субфракция №1; 0М NaCl; 0,05M NaCl; 0,15M NaCl; 0,2M NaCl и 2M NaCl – обозначение фракций, полученных после разделения СФ1 методом анионообменной хроматографии на ДЭАЭ 32, смывающихся при соответствующей концентрации NaCl.
Рис. 2. Влияние субфракции №1 и фракций, полученных после разделения СФ1 методом анионообменной хроматографии на ДЭАЭ 32, на пролиферацию ЭККЧ А431 в культуре (флуоресцентный тест).
СФ1 – субфракция №1; 0М NaCl; 0,15M NaCl; 0,2M NaCl и 2M NaCl – обозначение фракций, полученных после разделения СФ1 методом анионообменной хроматографии на ДЭАЭ 32, смывающихся при соответствующей концентрации NaCl.
Рис. 3. Вертикальный двуступенчатый ДСН-электрофорез в 6-20% ПААГ СФ1 и фракций, полученных после ее разделения методом анионообменной хроматографии на ДЭАЭ 32.
СФ1 – субфракция №1; 0М NaCl; 0,15M NaCl; 0,2M NaCl и 2M NaCl – обозначение фракций то же, что и на Рис. 2. Окраска на белок красителем Кумасси R-350. М – маркеры: 1 - bovine albumin (66 kDa); 2 - egg albumin (45 kDa); 3 – glyceraldehydes-3-phosphate dehydrogenase, rabbit muscle (36 kDa), 4 - bovine carbonic anhydrase (29 kDa); 5 – trypsinogen, bovine pancrease (24 kDa), 6 – trypsin inhibitor, soybean (20.1 kDa), 7 – б –lactalbumin, bovine milk (14.2).
Литература
, , и др. // Аллергология и иммунология. 2009. Т. 10, № 3. С. 363-364. , , и др. // Бюл. экспер. биол. мед. 2000. Т.129, № 2. С. 194-197. , , и др. // Патент № 000. Зарегистрирован 27.06.1995. Бюл. изобрет. 1995; 18: 24. , , и др. // Rus. J. Immunol. 1999. Vol. 4, N 2. P. 151-157. , , и др. // Патент № 000. Зарегистрирован в Государственном реестре изобретений РФ 27.04.2003. Москва. , , и др. // Иммунопатология аллергол. инфектол. 2005. № 3. С. 35-44. , , и др. // Патент. Регистрационный № 000/10(038939). Приоритет от 01.01.2001. Решение о выдаче патента 28.11.2012. , , // Цитология. 2003. Т. 45, № 2. С. 158-165. , , и др. // Цитология и генетика. 2001. Т. 35, № 6. С. 64-73. Denizot F., Lang R. // J. Immunol. Meth. 1986. V. 82. P. 271–277. Hirobe T. // Pigment. Cell Res. 2005. Vol. 18, N. 1. P. 2-12. Laemmli U. K. // Nature. 1970. Vol. 227. P. 680-685. Shirakata Y. J. // Dermatol. Sci. 2010. Vol. 59, N 2. P. therland J, Denyer M, Britland S. // J. Anat. 2005. Vol. 207, Is. 1. P. 67-78.Подписи авторов:
Автор, осуществляющий связь с редакцией:
119435, а.
ФГБУН НИИ ФХМ ФМБА России,
Тел.: (499) 246-43-42; (499) 246-42-96
E-mail: *****@***ru
Ф. И.О. авторов:
Ольга Владимировна Белова (O. V.Belova)
Валерий Иванович Сергиенко (V. I.Sergienko)
Виталий Яковлевич Арион (V. Ya. Arion)
Татьяна Абрамовна Луканидина (T. A.Lukanidina)
Светлана Николаевна Москвина (S. N.Moskvina)
Ирина Васильевна Зимина (I. V.Zimina)
Григорий Григорьевич Борисенко (G. G.Borisenko)
Геннадий Владимирович Луценко (G. V.Lutsenko)
Мария Владимировна Гречихина (M. V.Grechikhina)
Екатерина Валерьевна Ковалева (E. V.Kovaleva)
Жанетта Игоревна Ключникова (J. I.Klyuchnikova)
Автор, осуществляющий связь с редакцией:
119435, а.
ФГБУН НИИ ФХМ ФМБА России,
Тел.: (499) 246-43-42; (499) 246-42-96
E-mail: *****@***ru
