
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Личинки насекомых, зимующие под корой хвойных пород в окрестностях деревни Карманово
Руководитель —
Обоснование темы. Насекомые играют большую роль в жизни человека, причем большинство из них наносит прямой или косвенный вред (Аргиропуло др., 1948). Многие вредители-ксилофаги массово уничтожают запасы древесины и леса (Ижевский и др., 2005). Так, в Сибири, к 2017 г. площадь леса, пораженная пихтовым полиграфом (Polygraphus proximus Blanford), составила 490 000 км2. Настоящим бедствием для подмосковных лесов в последние годы стал короед-типограф (Ips typographus L.) на борьбу с которым тратятся миллиарды рублей (Савин, 2013). При этом есть виды, способные наносить вред только на имагинальной стадии развития, а есть насекомые, вредящие на личиночной стадии (Медведев, 1952). Определение и выявление экологических особенностей личинок таких видов — один из начальных этапов борьбы с насекомыми-вредителями (Мамаев, 1972; Ижевский и др., 2005).
Цель работы — изучить виды насекомых, зимующих в личиночной стадии под корой хвойных пород на севере Московской области.
Задачи: 1 — определить видовой состав личинок насекомых, зимующих под корой сосны и ели; 2 — установить фоновые виды для каждой из пород; 3 — сравнить породы и лесные формации между собой по набору видов; 4 — оценить обилие насекомых под корой обеих исследованных пород; 5 — выявить особенности зимовки насекомых под корой исследованных пород.
Методика. Работа проводилась 30.10–5.11.2016 г. окрестностях д. Карманово Талдомского р-на Московской обл., на территории на Кармановского участка Темповского охотхозяйства (в Танинском участковом лесничестве) (56.70319° с. ш., 37.19929° в. д. — 56.70319° с. ш., 37.21229° в. д. — 56.37031° с. ш., 37.20015° в. д. – 56.70317° с. ш., 37.198608° в. д. — 56.70319° с. ш., 37.19637° в. д. — 56.70313° с. ш., 37.20082° в. д.) на площади 0.22 км2. Была выбрана маршрутная методика. Биотопы были схематично описаны по ярусам леса (Дунаев, 1999). В них были обследованы сухостойные деревья ели европейской (Picea abies (L.) H. Karst.) и сосны европейской (Pinus sylvestris L.). Собранные личинки насекомых фиксировались в 75%-ном растворе этилового спирта. Определение насекомых проводили в камеральных условиях с помощью определителей (Арнольди и др., 1964; Мамаев, 1972; Мамаев и др., 1976; Ижевский и др., 2005). Травяные растения диагностировали по (1990), деревянистые растения — по (1999). Для сравнения пород и биотопов между собой по набору видов насекомых была использована формула Чекановского-Съёренсена, а обилие насекомых оценивали по шкале (Дунаев, 1997).
Результаты. За время проведения исследований было обследовано пять биотопов, обнаружено 133 экземпляра личинок 16 видов из 5 отрядов на 75 стволах деревьев (27 сосен и 48 елей).
Биотоп 1 представлял собой сосняк с ФСД (формулой состава древостоя) 10Со + ед. Бб. Биотоп был темным — сомкнутость крон составляла 75–80%. Захламленность и фаутность были по 2.5 баллов. Преобладала сосна обыкновенная (Pinus sylvestris L.), берёза бородавчатая (Betula pendula Roth.) росла по краю биотопа. В подросте в световых окнах скоплениями преобладала рябина обыкновенная (Sorbus aucuparia L.), рассеянно по биотопу встречались дуб черешчатого (Quercus robur L.) и ели европейской (Picea abies (L.) H. Karst). По краю биотопа в подросте рос клён ясенелистный (Acer negundo L.). По всему биотопу одиночно встречались липа мелколистная (Tilia cordata Mill.) и клён остролистный (Acer platanoides L.). В подлеске доминировали жимолость лесная (Lonicera xylosteum L.) и крушина ломкая (Frangula alnus Mill.). Были встречены скопления бересклета бородавчатого (Euonymus verrucosa Scop.), черёмухи птичьей (Padus avium Mill.) и дёрена белого (Cornus alba L.). Единично отмечена лещина обыкновенная (Corylus avellana L.). В травяно-кустарничковом ярусе доминировали кислица обыкновенная (Oxalis acetocella L.), звездчатка ланцетолистная (Stellaria holostea L.) и средняя (S. media L.) и недотрога мелкоцветковая (Impatiens parviflora DC.), росшие по всему биотопу. Одиночно были найдены хвощ лесной (Equisetum sylvaticum L.) и черника (Vaccinium myrtillus L.).
Биотоп 2 — березово-сосновый лес с ФСД 3Бб7Со + ед. Ее. Сомкнутость крон — 65%, захламленность и фаутность — по 1.5 баллов. Сосна обыкновенная и берёза бородавчатая росли обособленно друг от друга, ель европейская — на границе биотопа. В подросте было много сосны обыкновенной и ели европейской. Скопления образовывала рябина обыкновенная. Осина (Populus tremula L.) и ольха черная (Alnus glutinosa (L.) Gaertn.) встречались одиночно. В подлеске было несколько кустов жимолости лесной. На краю биотопа скоплением произрастала ива козья (Salix caprea L.). Было найдено несколько кустов черемухи птичьей. В травяно-кустарничковом ярусе доминировали злаки (Poaceae), земляника лесная (Fragaria vesca L.) и живучка ползучая (Ajuga reptans L.). Много было звездчатки ланцетолистной. Одиночно росли яснотка крапчатая (Lamium maculatum L.) и кислица обыкновенная.
Биотоп 3 представлял собой ельник с ФСД 8Ее1Бб1Лм + ед. Ос. Основные породы (ель, береза и липа) были равномерно распределены по биотопу. Осина единично встречалась на краю биотопа. Лес был очень темным — сомкнутость крон достигала 80–85%. Фаутность была невысокой — 2 балла, а захламленность — 3.5–4 балла. В подросте доминировали ель европейская, ольха черная и рябина обыкновенная. Меньше было подроста осины и липы мелколистной. Одиночно был отмечен дуб черешчатый. В подлеске доминировали черемуха птичья и жимолость лесная. Были встречены скопления дёрена белого и волчеягодника обыкновенного. На краю биотопа росла ива ушастая (Salix aurita L.). В травяно-кустарничковом ярусе доминировали черника и брусника. Очень много было кислицы обыкновенной и звездчатки ланцетолистной. Были одиночные находки земляники лесной и щитовника игольчатого.
Биотоп 4 — березняк с ФСД 7Бб2Ее1Ос + ед. Лм, Бп. Основные лесообразующие породы (берёза, ель и осина) были равномерно распределены по биотопу. Единично были встречены ольха черная, липа мелколистная и берёза пушистая (Betula pubescens Ehrh.). Сомкнутость крон — 55–60%, фаутность — 1.5 балла, захламленность — 3.5 балла. В подросте преобладали ольха черная, ольха серая (Alnus incana (L.) Moench), ель европейская и осина. Реже встречался подрост сосны обыкновенной, липы мелколистной и рябины обыкновенной. В подлеске доминировала крушина ломкая, жимолость лесная и черёмуха птичья. Большими скоплениями росла смородина чёрная (Ribes nigrum L.), одиночно росли волчеягодник обыкновенный и калина красная (Viburnum opulus L.). В травяно-кустарничковом ярусе преобладали злаки, звездчатка ланцетолистная, лютик ползучий (Ranuculus repens L.) и черника. Были найдены обширные скопления дудника лесного (Angelica sylvestris L.), зеленчука желтого, щитовника игольчатого и растения рода Бодяк (Cirsium). Меньше было земляники лесной, копытня европейского и ожики волосистой (Luzula pilosa (L.) Wild.). В состоянии ветоши был найден хвощ лесной.
Биотоп 5 — смешанный лес с ФСД 5Ее4Бб1О + ед. Олч. Ель европейская была выше березы бородавчатой. Осина и ольха черная встречались редко. Лес был тёмным, сомкнутость крон — 70–75%, захламленность и фаутность были невысоки — 1–1.5 балла. В подросте преобладала рябина обыкновенная скоплениями в световых окнах. По всему биотопу в подросте встречались осина, липа мелколистная и ольха черная. Очень много было ели европейской. Одиночно встречался дуб черешчатый. В подлеске доминировали крушина ломкая, лещина обыкновенная и жимолость лесная, росшие скоплениями в световых окнах. Местами встречались бересклет бородавчатый и волчеягодник обыкновенный (Daphne mezereum L.). В травяно-кустарничковом ярусе доминировали черника, копытень европейский (Asarum europaeum L.) и брусника (Vaccinium vitis-idaea L.), росшие скоплениями. Много было злаков. Меньше было щитовника игольчатого, звездчатки ланцетолистной и мокрицы. Единично были встречены хмель обыкновенный (Humulus lupulus L.) и паслен сладко-горький (Solanum dulcamara L.).
Размерные параметры основных лесообразующих пород биотопов представлены в табл. 1.
Таблица 1. Предельные высоты (h, м) и средний диаметр (ш, см) лесообразующих пород в биотопах (1–2).
Биотоп | Порода | h | ш |
1 | сосна | 22–24 | 27.8 |
2 | сосна | 20–22 | 25.2 |
береза | 18–20 | 23.5 | |
3 | ель | 23–24 | 25.2 |
береза | 19–21 | 23.2 | |
липа | 19–21 | 24.0 | |
4 | ель | 23–25 | 26.0 |
береза | 21–23 | 23.1 | |
осина | 20–21 | 23.9 | |
5 | ель | 22–24 | 25.7 |
береза | 20–21 | 23.0 | |
осина | 23–25 | 25.8 |
Большинство видов встречается на стволах одиночно, лишь четыре вида Coleoptera (25 %) встречались в среднем по 3 особи и один вид Diptera: Ceratopogonidae (6.2 %) встречались в большем количестве особей (в среднем по 23 особи) на один ствол (табл. 2). Подобное распределение соответствует кривой Раункиера (McIntosh, 1967; Макинтош, 2013) и правилу А. Тинемана, демонстрирующих доминирование суммы редких видов над числом фоновых (рис. 1), что вполне согласуется с принципом конкурентного исключения Вольтерры — Гаузе (Силин, Дунаев, 2011).
Таблица 2. Встречаемость (в %) и обилие (Р, в баллах) личинок видов насекомых на обследованных стволах (n) хвойных пород в разных биотопах (1–5).
Отряд | Порода | Сосна | Ель | Р | |
Биотоп (n) Таксон | 1 (23) | 2 (4) | 3 (25) | 4 (8) | 5 (15) |
Neuroptera | Hemerobiidae | 25 | 1 | ||
Lepidoptera | Polia nebulosa (Hufnagel) | 4.3 | 4 | 1 | |
Pyralidae | 4 | 13.3 | 1 | ||
Acronita sp. | 13.3 | 1 | |||
Euthrix (Phiulodora) potatoria L. | 12.5 | 1 | |||
Hymenoptera | Pristiphora sp. | 4.3 | 4 | 6.6 | 1 |
Coleoptera | Cucujus haematodes Erichson | 8.6 | 25 | 3 | |
Curculionidae | 25 | 6.6 | 1 | ||
Peltis (=Zimioma) grossum (L.) | 8 | 6.6 | 2 | ||
Pytho depressus (L.) | 4.3 | 3 | |||
Anthicidae | 4 | 1 | |||
Rhagium inquisitor L. | 4.3 | 3 | |||
Diptera | Therevidae | 25 | 12.5 | 6.6 | 1 |
Tipula irrorata Macquart | 4 | 1 | |||
Platypeza fasciata Meigen | 4 | 6.6 | 1 | ||
Ceratopogonidae | 17.3 | 5 |
Распределение фоновых видов видоспецифично и по породам, и по биотопам, в то время как более редкие виды могут встречаться в нескольких биотопах и на обоих породах (табл. 2). Наибольшее сходство (табл. 3) имеют ельник и березово-еловый лес (биотопы 3 и 5) что можно объяснить их сходством по видовому составу основных лесообразующих пород и степени освещенности.
Видовой состав насекомых, обитающих на лиственных и хвойных породах в зимний период в данном регионе заметно отличаются (Силин, Дунаев, 2009), но в целом преобладают Coleoptera и Diptera (Ющенко, Дунаев, 2012). Сходство ели и сосны по набору обитающих под корой насекомых составляет 40 %. Сосна в целом больше заражена вредителями древесины, чем ель, вероятно, из-за того, что её фитонцидные свойства выше (Цывин, 1973).
Фитофаги и ксилофаги на равных преобладают среди разных трофических групп личинок (рис. 2), что отличается от исследований, проведенных в этом регионе и ранее (Гомыранов, Дунаев, 2010). Число ксилофагов выросло, что можно связать с ослаблением деревьев из-за хозяйственной деятельности человека.
Визуально установлено, что на ели насекомые обитали по всему стволу, а на сосне в основном выше прикорневой зоны.
Личинки семейства Therevidae встречаются в сильно трухлявой древесине ели (табл. 2), что характерно для них в лесной зоне, т. к. обычные их местообитания — подстилка и песчаный грунт (Кривошеина, Кривошеина, 2015: 173).
N
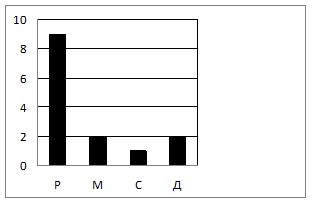
Рис. 1. Распределение числа видов (N, в %) по классам встречаемости (кривая Раункиера): Р — редкие, М — малочисленные, С — субдоминанты, Д — доминанты.
Таблица 3. Сходство между биотопами (1 — сосняк, 2 — березово-сосновый лес, 3 — ельник, 4 — березняк, 5 — березово-еловый лес) по набору насекомых под корой хвойных пород (в %).
1 | 2 | 3 | 4 | 5 |
1 | 20 | 31 | 0 | 13 |
2 | 0 | 33 | 18 | |
3 | 22 | 57 | ||
4 | 22 |
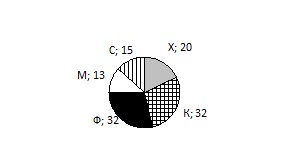
Рис. 2. Соотношение личинок насекомых разных трофических групп (в %): С — сапрофаги, М — микофаги, Ф — фитофаги, Х — хищники, К — ксилофаги.
Фитофаги и ксилофаги преобладают среди разных трофических групп личинок (рис. 2), что отмечали в этом регионе и ранее (Гомыранов, Дунаев, 2010).
Выводы
1. В окрестностях д. Карманово в обследованных биотопах под корой сосны и ели обитает не менее 16 видов насекомых в личиночной стадии.
2. Фоновые виды для обеих пород — личинки Pristiphora sp. Фоновый вид для сосны — Cucujus haematodes, для ели его выделить не удалось в связи со сходной степенью встречаемости большинства видов.
3. Наиболее близкие по видовому составу биотопы — ельник и березово-еловый лес, возможно благодаря сходству состава основных лесообразующих пород и освещенности.
4. Самый обильный вид — представитель семейства Ceratopogonidae, обитающий на сосне. Для него характерна зимовка очень большими скоплениями.
5. Из-за лучших фитоницидных свойств ель заражена вредителями древесины значительно меньше, чем сосна.
6. Распределение зимующих под корой сосен и елей личинок насекомых соответствует правилу Раункиера.
7. Личинки семейства Therevidae встречаются только в сильно трухлявой древесине ели, что характерно для них в лесной зоне, т. к. обычные их местообитания — подстилка и песчаный грунт.
Литература
, , Бей-, , , , , , Римский-, , 1948. Определитель насекомых европейской части СССР. — М.–Л.: ОГИЗ: 1128 с.
Арнольди, Л. В., , , , Мамаев, Б. М. , , 1964. Определитель обитающих в почве личинок насекомых. — М.: Наука, 919 с.
, , 2010. Экологические особенности распределения жуков (Hexapoda: Coleoptera) и их личинок под корой мертвых деревьев зимой. — Материалы I, II и III ежегодной Московской городской конференции учебно-исследовательских биологических работ школьников («Кобра») «Природа и исследовательская деятельность школьников». М.: ДНТТМ, с. 47–50.
, 1997. Методы эколого-энтомологических исследований. — М.: МГСЮН, 44 с.
, 1999. Деревянистые растения Подмосковья в осенне-зимний период. Методы экологических исследований. — М.: МГСЮН, 96 с.
, , 2005. Жуки-ксилотрофаги — вредители древесных растений России. — М.: Лесн. промышленность, 120 с.
, , 2015. Определитель двукрылых насекомых подотряда Brachycera — Orthorrhapha по личинкам. — М.: КМК, 384 с.
, 2013 Индекс разнообразия и соотношение некоторых концепций разнообразия. Ї Самарская Лука: проблемы региональной и глобальной экологии, т. 22, № 1, с. 104–127.
, , 1976. Определитель насекомых европейской части СССР. Ї М.: Просвещение, 304 с.
, 1972. Определитель насекомых по личинкам. — М.: Просвещение, 339 с.
, 1952. Личинки пластинчатоусых жуков. Определитель по фауне СССР, т. 47. — М.–Л.: АН СССР, 224 с.
, 1982. Принципы и методы количественного анализа в фаунистических исследованиях. — М.: Наука, с. 9–12.
2013. Короед-типограф в Подмосковье: казнить нельзя помиловать [Электронный документ]. — РИАМО. URL: https://riamo. ru/article/15582/koroed-tipograf-v-podmoskove-kaznit-nelzya-pomilovat. xl (дата публикации и последнего обновления: 23.09.2013, дата обращения: 20.06.2017).
, , 2009. Сравнение фауны насекомых под корой валежин березы и ольхи. — Материалы XXIII Московской городской конференции экспедиционных экологических отрядов учащихся. М.: МГСЮН, с. 43–48.
, , 2011. Сравнение лихенофлоры основных лесообразующих пород Владимирской Мещеры. — Мат-лы XXV Московской городской научной конф. экол. экспедиционных отрядов (в рамках II Московского экол. форума учащихся). М.: Экопресс, с. 57–61.
, 1990. Лесные травы. Определитель по вегетативным признакам. — Воронеж: ВГУ, 176 с.
, 1973. Использование древесной коры. — М.: Лесн. промышленность, 96 с.
, , 2012. Эколого-таксономический анализ зимующих под корой деревьев личинок насекомых в лесах Кармановского участка Темповского лесничества (Московская область). — Материалы V ежегодной Московской городской конференции учебно-исследовательских биологических работ школьников («КоБРа») «Природа и исследовательская деятельность школьников». М.: ДНТТМ, с. 74–77.
McIntosh R. P., 1967. An index of diversity and the relation of certain concepts to diversity. — Ecology, v. 48, № 3, р. 392–402.
