
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

Терапия белком Ламинин-111 предотвращает поражение мышц у мышиной модели мышечной дистрофии Дюшенна
Мышечная дистрофия Дюшенна (МДД) является наиболее распространенной формой мышечной дистрофии, диагностируемой у 1 из 3500 новорожденных мальчиков. Пациенты с МДД страдают от сильной, прогрессивной атрофии мышц, с клиническими симптомами впервые обнаруженными в 2-5 лет. По мере прогрессирования болезни пациенты теряют способность ходить в подростковом возрасте и умирает в возрасте 20 лет от сердечно-легочной недостаточности. Там в настоящее время нет эффективного лечения МДД. Пациенты с МДД и MDX мыши имеют мутации в гене дистрофина. Эти мутации приводят к отсутствию дистрофина, 427 кДа белка цитоскелета, который наряду с другими белками обеспечивает механическую связь между цитоскелетом клетки и ламинином во внеклеточном матриксе. У пациентов с МДД в скомпрометированной системе связи дистрофина мышечные волокна отделяются от внеклеточного матрикса во время сокращения мышц, что приводит к прогрессирующей потере целостности мышц и функции. В отсутствие дистрофина ?7?1 интегрин повышается в скелетных мышцах пациентов с МДД и MDX мышей. ?7?1 интегрин является преобладающим ламинин-связывающим интегрином в сердечной и скелетных мышцах. Мутации в гене ?7 интегрина вызывают врожденные миопатии у людей и мышей (8-10). Трансгенная сверхэкспрессия ?7-интегрина в скелетных мышцах мышей с дистрофией улучшает мышечную патологию и увеличивает продолжительность жизни. С другой стороны, потери ?7-интегрина у MDX мышей приводит к более тяжелому фенотипу и снижению жизнеспособности, с преждевременной смертью в 4-недельном возрасте. Вместе взятые, эти результаты подтверждают гипотезу о том, что ?7?1 интегрин является одним из основных модификаторов прогрессии заболевания и лекарственная терапия на его основе может облегчить МДД. Для идентификации молекул, которые способствуют экспрессии ?7 интегрину мы разработали клеточный анализ на промотерную активность ?7 интегрина. С помощью этого анализа, мы определили, что ламинин-111 увеличивает ?7-интегрин у мышей и МДД клеток. Внутримышечное или системное введение ламинина-111 мышам MDX увеличил экспрессию ?7 интегрина, что предотвращало наступление мышечной дистрофии и защищало мышцы от травмы при физической нагрузке. Вместе наши результаты определили, что ламинин-111 является эффективным терапевтическим белком у MDX мышей.
Результаты.
Ламинин-111 увеличивает ?7-Интегрин промоторную активность.
Чтобы проверить молекулы, которые увеличивают ?7-интегрин, мы разработали клеточный анализ мышц. Мы уже сообщали ранее о выведении ?7-интегрина нулевой мыши, у которой экзона 1 ?7-интегрина гена был заменен на ген LacZ репортера. У этих мышей все регулирующие транскрипцию элементы промотора ?7-интегрина сохраняются, позволяя ?-галактозидазе осуществлять выделение ?7-интегрина промотора. Первичные миобласты (обозначенного ?7?gal + / -), выделенные из 10-дневных ?7 + / - мышей анализируют на способность ?-галактозидазы активировать ?7-интегрин. ?7?gal + / - миобласты дифференцировались, затем их подвергали X-Gal окрашиванию и вестерн-блотту (фиг. 1 и В). ?-галактозидаза в ?7?gal + / - клетках увеличилось при миогенной дифференцировке, в соответствии с паттерном экспрессии ?7-интегрина в миобластах и мышечных трубках. Эти результаты подтверждают, что ген LacZ репортера в ?7?gal + / - мышечных клетках точно передает транскрипционную активность промотера ?7-интегрина.
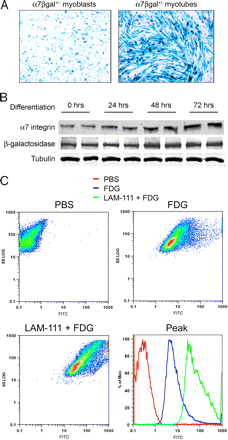
Несколько линий доказательств указывают на положительную обратную связь в регуляции экспрессии ламинина и ?7 интегрина. Чтобы проверить гипотезу, что ламинин регулирует экспрессию ?7 интегрина, ?7?gal + / - миобласты подвергали обработке 0-200 нМ ламинином-111 в течение 24 часов. Активность промотера ?7-интегрина измерялась расщеплением ?-галактозидазой нефлуоресцентного соединения флуоресцеина ди-?-D-галактопиранозида (ФДГ) с флуоресцеином. FACS показали, что лечение ?7?gal + / - миобластов в течение 24 ч 100 нМ ламинином-111 произвело к максимальному увеличению активности промотора ?7 интегрина (рис. 1в). Эти результаты показывают, что ламинин-111 способствует экспрессии ?7-интегрина в изолированных клетках мыши. Ламинин-111 увеличивает уровни ?7-Интегрина в мышиных и человеческих клетках.
Мы исследовали количественно уровни ?7 интегрина у C2C12 мыши и МДД первичных миобластов обработанных ламинином-111. Миобласты С2С12 обрабатывали 100 нМ ламинина-111 или PBS в течение 24 ч и анализировали с помощью Вестерн-блоттинга на ?7B-интегрина (фиг. 2А). Ламинин-111 привел к 2,2-кратному увеличению ?7B-интегрина в миобластах С2С12, подтверждая, что ламинин-111 способствует экспрессии ?7?1 (Рис. 2В).
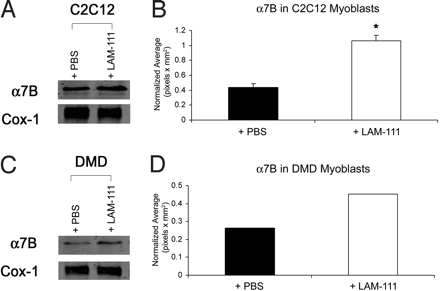
Мы тогда определили, увеличивает ли ламинин-111 ?7 интегрин в мышечных клетках. Первичные МДД миобласты обрабатывали 100 нМ ламинина-111 или PBS в течение 24 ч, а белковые экстракты подвергали Вестерн-блоттингу для ?7B-интегрина (фиг. 2С). Ламинин-111 произвел 1,7-кратное увеличение ?7B интегрина по сравнению с PBS-обработанными клетками (рис. 2г). Эти данные показывают, что механизм, посредством которого ламинин-111 увеличивает ?7-интегрин сохраняется в мышиных и человеческих клетках и предполагает, что ламинин-111, очень вероятно, увеличит ?7-интегрин в скелетных мышцах пациентов с МДД.
Внутримышечные инъекции Ламинина-111 предотвращают развитие заболевания у MDX мышей.
![]()
Мы затем определили, может ли ламинин-111 увеличивать ?7-интегрин в скелетных мышцах в естественных условиях. В левую переднюю большеберцовую (ТА) мышцу 10-дневных мышей MDX вводили 100 мкл 100 нМ ламинина-111, в то время как в правыую мышцу TA вводили 100 мкл PBS. В возрасте 5 недель, мыши были убиты, и TA мышцы были собраны. Ламинин-111 обычно не экспрессируется у взрослых или дистрофических мышц и иммунофлюоресценция выявила, что введенный ламинин-111 был депонирован в базальной мембране мышц ТА 5-недельных мышей MDX (Рис. 3А).
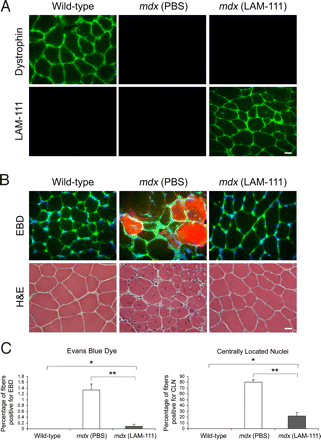
Для определения уменьшения ламинином-111 мышечной патологии у мышей MDX, провели окраску синим красителем Эванса (EBD) и гематоксилином / эозином (H & E). Окрашивание проводили на криосрезах у всех групп мышей (рис. 3б). MDX мышцы обработанные ламинином-111 имели в 12 раз меньше мышечных волокон положительных при окраске EBD по сравнению с контралатеральным контролем (рис. 3в). Кроме того, MDX мышцы обработанные ламинином-111 показали 4-кратное снижение доли мышечных волокон с центрально расположенными ядрами (рис. 3C). Эти результаты показывают, что внутримышечные инъекции ламинина-111 резко увеличивают целостность сарколеммы и уменьшают дегенерацию мышечных волокон.
Внутримышечные инъекции Ламинина-111 у мышей MDX повышают ?7-интегрин.
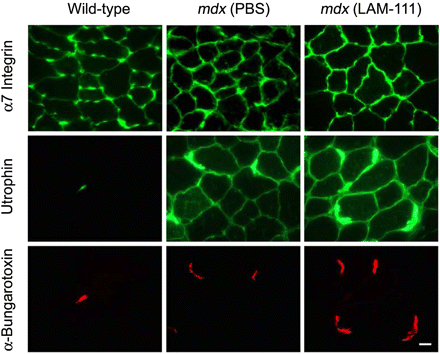
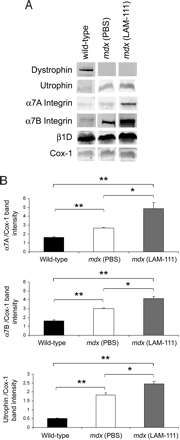
Чтобы определить механизм, посредством которого ламинин-111 защищает дистрофина-дефицитные мышцы от повреждений, иммунофлуоресцентный анализ атрофина и ?7-интегринов был выполнен. Наши результаты подтверждают повышенную экспрессию обоих белков в MDX скелетных мышцах, как сообщалось ранее. Лечение Ламинином-111 в дальнейшем повышало экспрессию ?7 интегрина в TA мышце мышей MDX (рис. 4).
Для подтверждения и количественной оценки этих наблюдений MDX мышцы подвергались вестерн-блоттингу (рис. 5а). 1,6 - и 2,6-кратное увеличение изоформ ?7A и ?7B-интегрина соответственно наблюдалось в ламинин-111-обработанных мышцах MDX сравнению с контрольной группой (фиг. 5В). Кроме того, 1,3-кратное увеличение атрофина наблюдалось в ламинин-111-обработанных мышцах (фиг.5В). Никаких существенных
![]()
изменений в уровнях ?1D интегрина не было замечено, что согласуется с результатами у трансгенных мышей. Эти результаты показывают, что ламинин-111 увеличил более чем в 4 раза экспрессию ?7 интегрина, белка, способного облегчить патологию дистрофических мышц.
Ламинин-111 может быть системно доставлен в MDX мышцы.
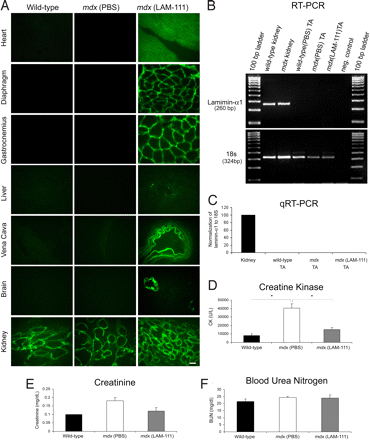
Пациенты с МДД страдают от атрофии мышц, поэтому эффективная терапия должна быть направлена ??на все мышцы, в том числе сердце и диафрагму. Поэтому необходимо определить, может ли ламинин-111 быть доставлен системно для этих мышц. Десяти-дневные MDX мыши получали внутрибрюшинно 1 дозу ламинин-111 в дозе 1 мг / кг, и ткани были проанализированы в возрасте 5 недель. Иммунофлуоресцентный анализ показал присутствие ламинина-?1 по всей базальной мембране икроножной мышцы, диафрагмы и кардиомиоцитов, в то время контроль был отрицательным (рис. 6A и рис. S1A).
![]()
Далее мы исследовали присутствие ламинина-111 в других тканях.
Ламинин-111, как правило, экспрессируется во взрослых почках и иммунофлюоресценцией обнаружен ламинин-?1 сигнал в почках мышей дикого типа и у PBS обработанных MDX мышей. Почки ламинина-111-обработанных мышей MDX показали повышение ламинина-?1 (фиг.6А). Мозг, печень и полая вена дикого типа и PBS обработанных MDX мышей не показали ламинин-?1 сигнала. В противоположность этому, ламинин-111-обработанные мыши показали MDX точечные области ламинина-?1 в печени и сильную иммунофлюоресценцию ламинин-?1 в полой вене и сосудах головного мозга (фиг. 6А). Сигнал не был обнаружен в паренхиме головного мозга, что указывает, что ламинин-111 не пересекает гематоэнцефалический барьер. Наконец, для подтверждения системной доставки, Alexa 488 меченный ламинин-111 внутрибрюшинно вводили MDX мышам, он был выделен спустя 48 ч для анализа. Alexa 488 меченный ламинин-111 был обнаружен в мембране (рис. S1B). Эти результаты показывают, что ламинин-111 может быть системно доставлен в скелетные и сердечную мышцы MDX мышей. Чтобы определить, является ли введенный ламинин-111 индуцированным эндогнной экспрессией, ламинин-?1-ПЦР была выполнена (фиг. 6В). Несмотря на то, ламинин-?1 транскрипт был обнаружен в почке мыши, как сообщалось ранее, ламинин-?1 транскрипт не был обнаружен у дикого типа или в MDX мышцах TA, получавшие буферный раствор или ламинин-111 (рис. 6В). Количественные TaqMan ОТ-ПЦР подтвердили эти наблюдения (рис. 6в). Эти данные свидетельствуют о том, что введенный ламинин-111 не вызывает экспрессию эндогенного ламинин-?1. Mdx мыши, получавшие ламинин-111, имеют нормальные уровни сывороточной креатинкиназы. Сывороточная креатинкиназа значительно выше у пациентов с МДД из-за повреждения мышц. Для определения терапевтического действия системной доставки ламинина-111 собирали сыворотку через 3 недели после однократной инъекции ламинина-111 для определения уровня креатинкиназы. Терапия ламинином-111 привела к 2,6-кратному снижению в сыворотке крови уровня креатинкиназы у MDX мышей, который статистически отличался от уровня наблюдаемого у животных дикого типа (рис. 6, г). Эти результаты показывают, что одна системная доза ламинина-111 предотвращает дистрофическую патологию у MDX мышей. Поскольку ламинин-111 большой белок и потенциально может отрицательно повлиять на функцию почек, мы измерили сывороточный креатинин и азот мочевины крови (АМК). Креатинин и мочевина статистически не отличались между ламинин-111-обработанными MDX и контрольными мышами (рис. 6 Е и F). Эти данные указывают на то, что терапия ламинином-111 не имела негативного воздействия на функцию почек.
MDX мыши, получавшие Ламинин-111, защищены от повреждений вызванных сокращением.
Чтобы изучить вопрос о том, как терапия ламинином-111 может предотвратить повреждения после физической нагрузки MDX мышей, которым вводили PBS или ламинин-111, мыши подвергались занатиям на беговой дорожке, и целостность сарколеммы анализировали поглощением EBD. Хотя TA мышцы PBS-обработанных мышей MDX показали большое количество EBD-положительных мышечных волокон, мыши, получавшие ламинин-111 показали несколько положительных мышечных волокон (фиг. 7А).
![]()
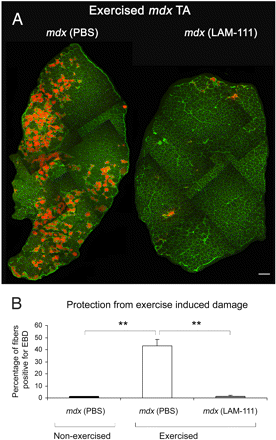
Количественный анализ этих наблюдений показал, что испытание на беговой дорожке произвело к 32-кратному увеличение EBD-положительных мышечных волокон по сравнению с мышами в покое (рис. 7б). Таким образом, беговая дорожка индуцирует значительное повреждение мышц у MDX мышей. В противоположность этому, ламинин-111-обработанные MDX мыши показали в 28 раз меньше EBD-положительных мышечных волокон по сравнению с контролем.
Обсуждение.
Несмотря на годы интенсивных исследований, до сих пор нет эффективного лечения МДД. Некоторые гены, как было показано, могут компенсировать потерю дистрофина и уменьшать дистрофические процессы, в том числе ?7?1 интегрин. Исследования показывают, что ?7?1 интегрин вносит свой вклад в структурную и функциональную целостность скелетных мышц. Поскольку ?7?1 интегрин экспрессируется повсеместно в скелетной и сердечной мышцах, небольшие молекулы, предназначенные для экспрессии этого гена, имеют значительные перспективы для лечения МДД. В этом исследовании мы определили ламинин-111 в качестве терапевтического белка для MDX мышиной модели МДД. Несмотря на то, ламинин-111 не экспрессируется в нормальных или дистрофических взрослых скелетных мышцах, исследования показывают, что это предпочтительный лиганд для ?7?1 интегрина. Лечение ламинином-111 стабилизировало сарколемму MDX скелетных мышц, снизило дегенерацию мышечных волокон, уменьшило содержание сывороточной креатинкиназы, защитило мышцы от повреждения при физической нагрузкой, что дает перспективу для дальнейшего развития терапии МДД. Пациенты с МДД обычно страдают от сердечно-легочной недостаточности, а системная доставка должны доставить препарат как к скелетным, так и сердечной мышце. К нашему удивлению, введенный ламинин-111 распределен по всей базальной мембране конечностей, диафрагмы и сердечной мышцы. После введения ламинина-111 у MDX мышей отмечаются уровни креатинкиназы как у мышей. Относительно большая молекулярная масса ламинина-111 (900 кДа), кажется, не стала препятствием для распространения по мышцам. Мышцы пациентов с МДД и MDX мышей очень восприимчивы к повреждению после физической нагрузки, и физические упражнения вызывает значительные повреждения в сарколеме MDX мышей. В этом исследовании мы показали, что системная доставка ламинина-111 не только предохраняет дистрофина-дефицитные мышцы от дегенерации, но и защищает мышцы от травмы, индуцированной сокращением. Эти результаты убедительно показывают, что терапия ламинином может предотвратить повторяющиеся циклы травмы, фиброз и потерю мышечной функции при МДД. Механизм, лежащий в основе защиты ламинина-111 в мышце MDX может включать повышенные уровни компенсационных белки и / или улучшенную адгезию. Наши исследования показывают ? 4-кратное увеличение ?7-интегрина. Небольшое увеличение атрофина наблюдалось после лечения ламинином-111, вряд ли ведет к улучшению мышечной патологии, поскольку исследования показывают, что значительно большее количество атрофина необходимо для терапевтической функции. В дополнение к повышенным уровням ?7 интегрина, ламинин-111 может также
![]()
механически укрепить сарколемму против сил сдвига во время мышечного сокращения. Наше исследование показывает, что ламинин-111 может быть очень мощным терапевтическим белком при МДД. Кроме того, терапия ламинином-111 может быть эффективным при лечении других заболеваний мышц, в том числе врожденной мышечной дистрофии типа 1А, конечности-поясной мышечной дистрофии, и ?7-интегрин врожденной миопатии. Эффективность ламинина-111 при МДД у мышиной модели показывает, что системное введение внеклеточных матричных молекул представляет собой новую парадигму для лечения многих генетических заболеваний.
Ссылки
? Bulfield G, Siller WG, Wight PA, Moore KJ (1984) X chromosome-linked muscular dystrophy (mdx) in the mouse. Proc Natl Acad Sci USA 81:1189–1192. Abstract/FREE Full Text
? Senter L, et al. (1993) Interaction of dystrophin with cytoskeletal proteins: Binding to talin and actin. Biochem Biophys Res Commun 192:899–904. CrossRef Medline Web of Science
? Campbell KP (1995) Three muscular dystrophies: Loss of cytoskeleton-extracellular matrix linkage. Cell 80:675–679. CrossRef Medline Web of Science
? Matsumura K, et al. (1994) Expression of dystrophin-associated proteins in dystrophin-positive muscle fibers (revertants) in Duchenne muscular dystrophy. Neuromuscul Disord 4:115–120. CrossRef Medline Web of Science
? Monaco AP, et al. (1986) Isolation of candidate cDNAs for portions of the Duchenne muscular dystrophy gene. Nature 323:646–650. CrossRef Medline
? Hodges BL, et al. (1997) Altered expression of the alpha7beta1 integrin in human and murine muscular dystrophies. J Cell Sci 110:2873–2881. Abstract
? Burkin DJ, Kaufman SJ (1999) The alpha7beta1 integrin in muscle development and disease. Cell Tissue Res 296:183–190. CrossRef Medline Web of Science
? Flintoff-Dye NL, et al. (2005) Role for the alpha7beta1 integrin in vascular development and integrity. Dev Dyn 234:11–21. CrossRef Medline Web of Science
? Hayashi YK, et al. (1998) Mutations in the integrin alpha7 gene cause congenital myopathy. Nat Genet 19:94–97. CrossRef Medline Web of Science
? Mayer U, et al. (1997) Absence of integrin alpha 7 causes a novel form of muscular dystrophy. Nat Genet 17:318–323. Medline Web of Science
? Burkin DJ, Wallace GQ, Nicol KJ, Kaufman DJ, Kaufman SJ (2001) Enhanced expression of the alpha 7 beta 1 integrin reduces muscular dystrophy and restores viability in dystrophic mice. J Cell Biol 152:1207–1218. Abstract/FREE Full Text
? Guo C, et al. (2006) Absence of alpha 7 integrin in dystrophin-deficient mice causes a myopathy similar to Duchenne muscular dystrophy. Hum Mol Genet 15:989–998. Abstract/FREE Full Text
? Rooney JE, et al. (2006) Severe muscular dystrophy in mice that lack dystrophin and alpha7 integrin. J Cell Sci 119:2185–2195. Abstract/FREE Full Text
? Song WK, Wang W, Foster RF, Bielser DA, Kaufman SJ (1992) H36-alpha 7 is a novel integrin alpha chain that is developmentally regulated during skeletal myogenesis. J Cell Biol 117:643–657. Abstract/FREE Full Text
? Peat RA, et al. (2008) The diagnosis and etiology of congenital muscular dystrophy. Neurology 71:312–321. Abstract/FREE Full Text
? Pons F, Nicholson LV, Robert A, Voit T, Leger JJ (1993) Dystrophin and dystrophin-related protein (utrophin) distribution in normal and dystrophin-deficient skeletal muscles. Neuromuscul Disord 3:507–514. CrossRef Medline
![]()
? Sasaki T, Giltay R, Talts U, Timpl R, Talts JF (2002) Expression and distribution of laminin alpha1 and alpha2 chains in embryonic and adult mouse tissues: An immunochemical approach. Exp Cell Res 275:185–199. CrossRef Medline Web of Science
? Burkin DJ, et al. (2005) Transgenic expression of {alpha}7{beta}1 integrin maintains muscle integrity, increases regenerative capacity, promotes hypertrophy, and reduces cardiomyopathy in dystrophic mice. Am J Pathol 166:253–263. Abstract/FREE Full Text
? Boppart MD, Volker SE, Alexander NM, Burkin DJ, Kaufman SJ (2008) Exercise promotes alpha7 integrin gene transcription and protection of skeletal muscle. Am J Physiol 295:R1623–R1630. Web of Science
? Boppart MD, Burkin DJ, Kaufman SJ (2006) Alpha7beta1-integrin regulates mechanotransduction and prevents skeletal muscle injury. Am J Physiol 290:C1660–C1665. CrossRef Web of Science
? von der Mark H, et al. (2002) Alternative splice variants of alpha 7 beta 1 integrin selectively recognize different laminin isoforms. J Biol Chem 277:6012–6016. Abstract/FREE Full Text
? Nishiuchi R, et al. (2006) Ligand-binding specificities of laminin-binding integrins: A comprehensive survey of laminin-integrin interactions using recombinant alpha3beta1, alpha6beta1, alpha7beta1 and alpha6beta4 integrins. Matrix Biol 25:189–197. CrossRef Medline Web of Science
? Straub V, Rafael JA, Chamberlain JS, Campbell KP (1997) Animal models for muscular dystrophy show different patterns of sarcolemmal disruption. J Cell Biol 139:375–385. Abstract/FREE Full Text
? Petrof BJ (1998) The molecular basis of activity-induced muscle injury in Duchenne muscular dystrophy. Mol Cell Biochem 179:111–123. CrossRef Medline Web of Science
? DelloRusso C, Crawford RW, Chamberlain JS, Brooks SV (2001) Tibialis anterior muscles in mdx mice are highly susceptible to contraction-induced injury. J Muscle Res Cell Motil 22:467–475. CrossRef Medline Web of Science
? Tinsley J, et al. (1998) Expression of full-length utrophin prevents muscular dystrophy in mdx mice. Nat Med 4:1441–1444. CrossRef Medline Web of Science
Переведено проектом МОЙМИО:
![]()
www.mymio.org
Оригинал статьи: http://www. pnas. org/content/10
