
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Лицей 1568
Исследовательская работа
Применение полимеразной цепной реакции для внесения точечных изменений в последовательность из генома мыши
Васильева Елена
Научный руководитель: к. б.н.
Москва, 2016-2017 гг.
Оглавление
1. Введение 3
2. Материалы и методы 9
2.1 Материалы, реактивы и приборы, использованные в работе 9
2.2 Методы 9
2.2.1 Полимеразная цепная реакция (ПЦР) 9
2.2.2 Горизонтальный электрофорез ДНК в агарозном геле 12
3. Результаты 12
3.1 Внесение точечных изменений в нуклеотидную последовательность ДНК 12
3.2 Детекция наличия в реакционной смеси продукта амплификации. 14
Выводы 16
Заключение 17
Литература 18
1. Введение
Согласно центральной догме молекулярной биологии (Рис. 1) перенос генетической информации осуществляется от нуклеиновой кислоты (ДНК или РНК) на другую нуклеиновую кислоту или белок. По этому правилу происходит экспрессия гена [1].
Регуляция экспрессии гена - сложный и многоуровневый процесс, протекающий на каждом уровне реализации генетической информации: транскрипции, процессинга и трансляции. На уровне транскрипции клетка реализует множество механизмов регуляции. В клетках эукариот существует несколько больших ферментативных комплексов, осуществляющих синтез РНК и работа каждого из таких комплексов регулируется по-своему [2]. Одним из наиболее важных регуляторных элементов является промотор - нуклеотидная последовательность, прилежащая к началу кодирующей области гена, которая ответственна за инициацию его транскрипции; с ней специфически связывается фермент ДНК-зависимая РНК-полимераза [3].
Эукариоты обладают различными типами РНК-полимераз, классифицируемыми по типам РНК, которые они производят [4]:
- РНК-полимераза I синтезирует 45S-предшественника рРНК, которая превращается в рРНК 28S, 18S и 5,8S, а они уже образуют главные РНК-секции рибосомы. РНК-полимераза II производит предшественников для мРНК, а также для большинства мяРНК и миРНК. Это наиболее хорошо изученный тип РНК-полимеразы. РНК-полимераза III, синтезирует тРНК, 5S рРНК и другие малые РНК, присутствующие в ядре и цитозоле.
Регуляция активности каждого типа РНК-полимераз осуществляется определенными типами промоторов.
Существуют и другие последовательности, являющиеся регуляторными элементами транскрипции [3;5]. Например,
модуляторы — последовательности ДНК, усиливающие уровень транскрипции. Усилители (энхансеры) — последовательности ДНК, усиливающие уровень транскрипции и действующие независимо от своего положения относительно кодирующей части гена. Терминаторы — специфические последовательности, прекращающие и транскрипцию, и трансляцию.Промоторы, узнаваемые РНК(полимеразой III, обычно делят на три класса, основываясь на их структурной организации и наборе факторов транскрипции, необходимых для эффективного считывания соответствующих генов [6].
К первому типу относятся промоторы генов 5S рРНК. Классическим примером является ген 5S рРНК шпорцевой лягушки Xenopus laevis (рис. 2), в состав которого входят три внутренних элемента, необходимых для эффективной транскрипции: А-блок, расположенный между нуклеотидами +50 и +64, промежуточный сегмент, локализованный в положении от +67 до +72, и С-блок – от +80 до + 97 .
Второй тип промоторов обнаружен в генах тРНК, VA-генах аденовирусов и многих семействах умеренно повторяющихся генов. Он состоит из двух высоко консервативных блоков А и В, расположенных внутри транскрибируемой области (рис. 2). Блоки А первого и второго типа промоторов гомологичны друг другу и иногда могут быть взаимозаменяемы, но в случае второго типа промоторов А блок располагается гораздо ближе к точке начала транскрипции. Расположение блока В сильно варьирует не только от вида к виду, но даже среди тРНК одного организма. Так, например, у пекарских дрожжей Saccharomyces cerevisiae расстояние между блоками А и В может составлять от 31 до 93 п. н.

Некоторые матрицы, транскрибируемые РНК полимеразой III, не содержат внутригенных промоторных элементов. Для таких генов характерно расположение перед геном и они называются промоторами третьего типа. Например, было установлено, что промоторы генов U6 мяРНК человека и мыши сохраняют полную активность при делеции последовательности, лежащей непосредственно за точкой начала транскрипции в направлении к 3'-концу.
Для генов с промотором РНК-полимеразы III второго типа транскрипция считалась не зависящей от прилегающих последовательностей. Этот факт сделал такие гены особенно интересными для использования в генной терапии. Однако в лаборатории Эволюции геномов эукариот (ИМБ РАН) было открыто, что иногда последовательности, лежащие перед соответствующим геном, могут влиять на эффективность его транскрипции [7].
Роль последовательностей, лежащих снаружи гена — очень актуальный вопрос. Лаборатория эволюции геномов эукариот, в которой я делала работу, занимается изучением этих последовательностей. И моя работа — часть большого исследовательского проекта по выявлению роли последовательностей, прилегающих к гену малой РНК, синтезируемой РНК - полимеразой III, в эффективности ее транскрипции. Эта РНК называется 4,5SH РНК.
В лаборатории изучается, что получится, если изменить нуклеотидную последовательность участка, прилегающего к началу гена этой малой РНК. Это многоэтапное исследование и на моем этапе нужно было внести точечные изменения в последовательность ДНК и увеличить количество ее копий. (Рис. 3)

Рис. 3 Блок-схема, показывающая этапы исследования
Для проверки как тот или иной нуклеотид влияет на синтез, нужно точечно внести изменения в последовательность ДНК. Существует множество методов, которые позволяют вносить такие изменения. В нашей работе был использован метод ПЦР - полимеразной цепной реакции.
Этот подход заключается в избирательном копировании определённого участка ДНК при помощи ферментов в искусственных условиях (in vitro). При этом происходит копирование только того участка, который удовлетворяет заданным условиям, и только в том случае, если он присутствует в исследуемом образце. При ПЦР проводится повторение цикла, за который ДНК денатурируется, далее отжигаются праймеры, и достраивается комплиментарная цепь ДНК [8]. Метод ПЦР был открыт в 1983 году американским биохимиком Кэри Муллисом и его сотрудниками, это изобретение произвело революцию в молекулярной биологии и молекулярной медицине [9]. Первоначально целью было создание метода, который позволит in vitro амплифицировать, то есть умножить, специфический участок ДНК, расположенный между двумя участками ДНК с известной последовательностью. Ранее можно было получить только минимальные количества ДНК специфического гена, теперь с использованием ПЦР даже единичная копия может быть амплифицирована до миллиона копий за не очень продолжительный отрезок времени. Через 8 лет после этого за изобретение метода ПЦР Кэри Муллис получил Нобелевскую премию.
С годами метод ПЦР совершенствовался трудами многих ученых, а сейчас эта методика стала необходимой в криминалистических исследованиях, в персонализированной медицине (в выявлении особенностей экспрессии генов и восприятия лекарственных препаратов), а также в анализе пищевых продуктов, в пробах на наличие генетически модифицированных ДНК или на микробиологическое заражение.
Многие модификации данного подхода используются с различными целями. В нашей работе мы применили редкую разновидность ПЦР - ПЦР с измененными праймерами.
Целью моей работы было внесение точечного изменения в последовательность из генома мыши при помощи метода ПЦР.
В связи с этим были поставлены задачи:
1. С помощью научной литературы понять и изучить принципы метода ПЦР и возможность его использования для изменения нуклеотидной последовательности.
2. Провести синтез фрагмента ДНК с измененной нуклеотидной последовательностью с помощью ПЦР.
3. Подтвердить, что синтез прошел успешно с помощью метода горизонтального электрофореза.
Объект исследования: фрагмент ДНК - нуклеотидная последовательность из генома мыши.
Гипотеза исследования: с помощью ПЦР можно получить последовательность, отличающуюся от изучаемой на один заданный нуклеотид.
2. Материалы и методы
2.1 Материалы, реактивы и приборы, использованные в работе
- Амплификатор для ПЦР: Eppendorf Прибор для электрофореза Bio-Rad Спектрофотометр NanoDrop1000 'Termo Scientific' (США) Набор для ПЦР «Силекс»
2.2 Методы
2.2.1 Полимеразная цепная реакция (ПЦР)
Как было сказано выше, полимеразная цепная реакция (ПЦР) — это экспериментальный метод молекулярной биологии, позволяющий добиться значительного увеличения малых концентраций определенных фрагментов нуклеиновой кислоты (ДНК/РНК) в биологическом материале (пробе) [8].
Метод ПЦР основан на механизме репликации ДНК in vivo (в пробирке): двуцепочечная ДНК раскручивается до одноцепочечной под действием температуры, дуплицируется и снова закручивается, далее цикл повторяется определенное количество раз.
- Денатурация ДНК путем плавления при повешенной температуре для превращения двуцепочечной молекулы в одноцепочечную. Отжиг (гибридизация) двух олигонуклеотидов, используемых в качестве праймеров для целевой ДНК. Удлиннение цепи ДНК, начиная от праймеров, путем добавления нуклеотидов с использованием фермента ДНК-полимеразы в присутствии ионов Mg2+
Но чтобы реакции прошли успешно необходимо наличие в реакционной смеси ряда основных компонентов.
- Анализируемый образец
Подготовленный для внесения в реакционную смесь препарат, содержащий молекулы ДНК, которые служат базой для последующего многоразового копирования. При отсутствии образца специфический продукт амплификации не образуется.
- Праймеры
Это искуственно синтезированный олигонуклеотиды, обычно имеющие размер от 15 до 30 нуклеотидов, идентичные соответствующим участкам мишени (ДНК). Праймеры играют ключевую роль в образовании специфичных продуктов реакции, и поэтому должны удовлетворять некоторым требованиям: во-первых, они должны быть специфичны. В противном случае велик вероятность синтеза цепи с ошибками, и последующей потери чувствительности реакции. Во-вторых, не должно образовываться устойчивых двойных цепей ( комплиментарного присоединения) прамеров самих на себя в результате отжига. В третьих, желательно, чтобы праймер был богат C-G связями, как известно они образованы тремя водородными связями, что повышает их прочность, в свою очередь это повышает специфичность и чувствительность при отжиге праймера. И последнее, область отжига праймеров не должна заходить на зону исследуемой последовательности, ради умножения копий которой проделывается ПЦР и подбираются праймеры.
- Taq - полимераза
В оригинальном методе ПЦР использовалась полимераза E. coli. Этот фермент денатурируется при температуре, более низкой, чем это требуется для большинства матричных дуплексов. В ранних экспериментах, свежий фермент требовалось добавлять в реакции после каждого цикла, что очень затормаживало процесс. Модификация процесса произошла после исследования термостабильной ДНК-полимеразы, в эксперименте я использовала фермент Taq-полимеразу, выделенную из бактерий Termus aquaticus (1998 год), живущих в горячих источниках при температуре, близкой к 85 градусам по Цельсию. Открытие термостабильных ДНК-полимераз облегчило процесс, так как добавление фермента требуется только в начале, при подготовке реакционной смеси.
- Смесь дезоксинуклеотидтрифосфатов (дНТФ)
Это «строительный материал», необходимый для синтеза ДНК. Концентрации всех четырех дНТФ должны быть эквивалентны для минимизации ошибок присоединения.
- Буфер
Буфером называют смесь катионов и анионов в определенной концентрации, обеспечивающей оптимальные условия для реакции. Важным компонентом являются ионы Mg2+, образующие растворимый комплекс с дНТФ, который необходим для присоединения нуклеотидов.
- Деионизированная вода
В рамках одного цикла ПЦР проходя 3 процесса: денатурация, отжиг праймеров и сам синтез.
- Денатурация ДНК — это соответственно перевод молекулы из двухнитевой формы в однонитевую, путем разрыва водородных связей между комплиментарными парами оснований. Отжиг — присоединение праймеров к денатурировавшим молекулам ДНК. Синтез — достраивание второй цепи ДНК, начиная с 3` конца молекулы с помощью Taq-полимеразы.
Для каждого процесса учеными подобраны различные температуры. Иногда, температура отжига праймеров и температура оптимальной работы фермента очень близки, и становится возможен двухэтапный ПЦР.
В работе реакционная смесь объемом 25 мкл содержала:
ДНК матрица 1 мкл
Праймер (прямой) 1.25 мкл (6 пмоль)
Праймер (обратный) 1.25 мкл (6 пмоль)
Буфер х10 2.5 мкл
дНТФ 1 мкл
Taq-полимераза 0.5 мкл
Деионизированная вода 17.5 мкл
2.2.2 Горизонтальный электрофорез ДНК в агарозном геле
Электрофорез в геле — аналитический метод в молекулярной биологии, позволяющий разделить фрагменты ДНК по размеру [10].
Электрофорез проводят в пластинке застывшего агарозного геля. Агароза образует пористую пространственную решетку, за счет этого более крупные фрагменты ДНК преодолевают путь медленнее, чем короткие. Двигаться молекулы ДНК начинают из-за постоянного электрического поля в геле, создаваемого двумя электродами. Это движение возможно за счет отрицательного заряда, который несут молекулы нуклеиновых кислот. Одним из компонентов геля является бромистый этидий, который интеркалирует ( то есть обратимо включается) между азотистыми основаниями и флюоресцирует в УФ-лучах, что позволяет определить местоположение молекул ДНК в геле.
3. Результаты
3.1 Внесение точечных изменений в нуклеотидную последовательность ДНК
В процессе выполнения работы были изучены основные принципы метода ПЦР. Были дизайнированы праймеры для внесения изменений в последовательность ДНК. (Рис. 4)
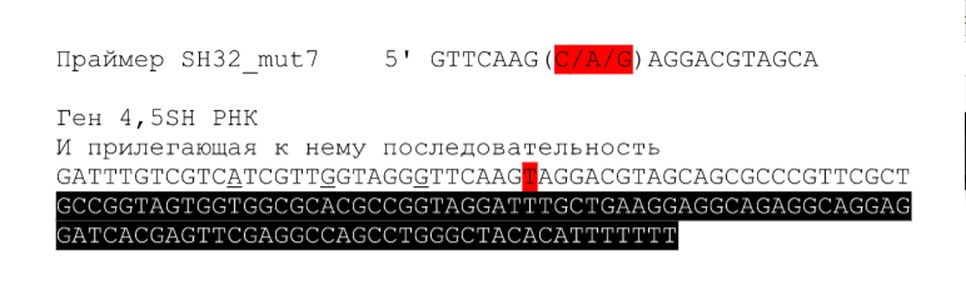
Красным цветом на рис. 4 обозначен нуклеотид (тимидин - Т) в цепочке ДНК, вместо которого мы хотели поставить другой нуклеотид (изменить последовательность). Соответствующая позиция обозначена в последовательности праймера тоже красным цветом. Мы использовали смесь трех праймеров, в каждом из которых на заданном месте был один из альтернативных нуклеотидов - А, Ц или Г (в последовательности на рисунке указаны через /, это было нужно для последующих этапов работы моих коллег).

Для постановки ПЦР в пробирке согласно методам и материалам были смешаны необходимые реагенты. Далее пробирку ставили в амплификатор, устанавливали подходящий температурный режим и время. (Таблица 1)
|
Длительность протекания каждой стадии - 1 минута, общее время реакции - 90 минут.
Таблица 1. температурный режим ПЦР
3.2 Детекция наличия в реакционной смеси продукта амплификации.
По окончанию реакции пробирки извлекли из прибора, 5 мкл реакционной смеси были смешаны со специальным буфером и использованы для детекции методом горизонтаольного электрофореза.
Для проведения электрофореза в специальную плашку заливали расплавленный агарозный гель, вставляли гребенку и ждали его полного застывания. Застывший гель поместили в прибор для электрофореза и залили специальным буфером для того, чтобы была возможность проведения электрического тока. Далее в лунки в геле наносили продукты ПЦР и маркер длин фрагментов ДНК и подключали ток. Через некоторое время наблюдали, насколько продвинулись полученные ДНК (см Материалы и методы). Визуально подтверждали наличие фрагментов ДНК соответствующей длины с помощью добавления вещества - бромистого этидия. В результате был детектирован фрагмент, по длине (150 н. п.) соответствующий требуемому. (Рис.5)
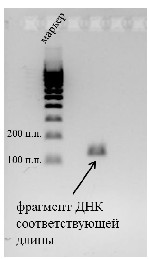
Поскольку с помощью горизонтального электрофореза нами было подтверждено наличие продукта реакции, пробирка с оставшейся реакционной смесью (20 мкл) была передана коллегам для дальнейшего исследования. На настоящий момент наличие изменений в нуклеотидной последовательности подтверждено методом секвенирования.
Выводы
Подтверждена возможность внесения изменений в последовательность ДНК с помощью ПЦР. Проведена модификация последовательности из генома мыши путем замены одного нуклеотида с помощью метода ПЦР. Визуально подтверждено наличие фрагмента определенной длины с помощью метода горизонтального электрофореза.
Заключение
В ходе работы был изучен принцип метода ПЦР. В настоящее время на основе этого метода, изначально созданного с целью специфичкской амплификации заданного участка ДНК, создано множество подходов для решения как научных, так и практических задач. ПЦР широко используется в медицине для диагностических целей при инфекционных заболеваниях, в экспертизе и установлении личности, для определения генетических особенностей организма. Есть надежда, что в будущем с помощью ПЦР будут создавать конструкции для лечения и коррекции генетических заболеваний [11].
В результате моей работы был получен фрагмент ДНК с измененной последовательностью, прилегающей к началу гена малой 4,5SH РНК. В настоящее время в Лаборатории эволюции геномов эукариот ИМБ РАН проверяют, как именно такое изменение последовательности влияет на эффективность синтеза это РНК с помощью РНК-полимеразы III. Полученные в результате этих экспериментов данные объяснят, за счет каких нуклеотидов происходит регуляция работы огромного ферментативного комплекса.
Эта информация очень важна, поскольку ранее считалось, что прилегающие, внешние для гена 4,5SH РНК последовательности, не оказывают влияния на эффективность его транскрипции.
