
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

Нанополимеры улучшают доставку олигонуклеатидов для пропуска экзона и следовательно экспрессию дистрофина в скелетных мышцах mdx мышей
Jason H Williams1, Rebecca C Schray1, Shashank R Sirsi2 and Gordon J Lutz1*
Corresponding author: Gordon J Lutz *****@***edu
Author Affiliations
1 Drexel University College of Medicine, Department of Pharmacology and Physiology, Philadelphia, Pennsylvania 19102, USA
2 Drexel University, Department of Biomedical Engineering, Philadelphia, Pennsylvania 19104, USA
For all author emails, please log on.
BMC Biotechnology 2008, 8:35 doi:10.1186/1472-6750-8-35
The electronic version of this article is the complete one and can be found online at: http://www. /1472-6750/8/35
Received: 15 October 2007
Accepted: 2 April 2008
Published: 2 April 2008
© 2008 Williams et al; licensee BioMed Central Ltd.
Введение.
Стерический блок олигомеров, такой как 2'O-метил (2'OMe) и фосфородиамидат морфолино (PMO) обладают высоким сродством к комплементарной преМРНК и можгут создавать альтернативное соединение, вызвать пропуск или включение определенных экзонов. Такие модулирующие сплайсинг олигонуклеатиды(SMO), представляют сильный класс составов с возможностью широкого использования в качестве фармацевтических препаратов. Однако, использование в естественных условиях SMO показало необходимость разработки носителей для доставки к ядрам клеток. Действительно, неэффективная поставка SMO остается главным ограничением их полноценности как фармацевтических препаратов.
Мышечная дистрофия Дюшена (МДД) - смертельная x-сцепленная болезнь, вызванная мутациями в гене в 427 килодальтонов связанного с мембраной белка дистрофина, заканчивающаяся прогрессивным ослаблением мышц во всем теле и смертью обычно в середине третьего десятилетия жизни. Мутации при МДД чаще всего состоят из вставок и делеций, которые изменяют рамку считывания или вызывают преждевременную остановку кодона [3]. Эти типы мутаций приводят к производству обрезанного и нефункционального дистрофина.
Стерический блок олигонуклеатидов, пропускающих экзон(ESOs), исследовался в культурах клеток мыши, собак и клеток человека, чтобы вызвать пропуск экзонов, заканчивающийся производством мРНК дистрофина во всю длину (минус только пропущенные экзоны), и восстановление рамки считывания [4-10]. У животных - классической модели МДД - mdx мыши, есть точечная мутация в экзоне 23 гена дистрофина, которая производит к преждевременной остановке кодона. ESO, как показывали, вызвал пропуск экзона 23 и сопутствующее экспрессию дистрофина в скелетных мышцах mdx мышей и после местной и после системной поставки [8,11-17], и эта технология была недавно рассмотрена в контексте терапии для МДД [18,19]. Однако, эффективность 2'OMe поставки ESO и уровень сопутствующего экспрессии дистрофина у mdx мышей остается относительно низкой [11,13,16]. Несмотря на то, что ESO без носителя прошли до клинических испытаний фазы I, носители должны быть найдены прежде, чем этот подход можно рассмотреть как терапию для пациентов с МДД.
Богатый амином катионный полимер поли-(этиленовый имин) (PEI) является хорошо изученным составом, который эффективен при сжатии большой ДНК плазмиды и позволяет улучшить клеточное поглощение [20-25]. Хотя это относится не только к поставке плазмиды, недавние исследования также зарегистрировали PEI-расширенную поставку маленьких нуклеиновых кислот, таких как олигонуклеатиды и siРНК [26-33]. Положительный поверхностный заряд PEI-нуклеиновой-кислоты взаимодействует с отрицательно заряженными элементами на клеточной мембране, стимулируя поглощение путем эндоцитоза[20,21,24,25,34,35]. После того как PEI проникает в клетку происходит разрывание эндосомальной мембраны из-за осмотического лизиса [34,36-38]. В то время как внутриклеточное передвижение комплексов PEI-нуклеотида, однажды вышедших из эндосомы, неизвестны, олигонуклеатид должен отделиться от PEI, чтобы достигнуть ядра, поскольку ядерная оболочка, вероятно, непроницаема для PEI. Используя двойной флюоресцентный метод мы недавно показали, что кополимеры PEG-PEI увеличивали поставку SMO к ядру культивируемых mdx мышечных волокон, в то время как кополимеры, главным образом, не поступали в ядра [39].
Функциональность и биологическая совместимость PEI улучшены инкорпорацией неионогенного линейного полимера поли-(этиленовый гликоль) (PEG) в кополимеры PEG - PEI [23,24]. Макромолекулярные свойства PEG-PEI-oлигонуклеатида находятся под влиянием молекулярной массы PEI и природы PEGylation, который обуславливает способность трансфекции [22,23,40]. Мы недавно показали, что кополимеры PEG - PEI сделанные из MW PEI2K значительно улучшают поставку ESO к мышечным волокнам mdx мышей после внутримышечных инъекций по сравнению с MW PEI25K кополимером [13]. Мы приписывали превосходящую эффективность основанных на PEI2K кополимеров к низкому поверхностному заряду и высокой стабильности наночастиц, сформированных во время комплексообразования с ESO [41]. Хотя внутримышечная инъекция этих PEG - PEI-ESO комплексов произвела значительно больше дистрофин-положительных волокон, чем один ESO, уровень экспрессии дистрофина при Вестерн-блотт анализе достиг только 2-5% нормального уровня в ПББМ( передняя большеберцовая мышца). Поэтому, в этом отчете мы оценили, могли ли бы новые режимы инъекций кополимеров PEG - PEI улучшить экспрессию. дистрофина Наши результаты показывают, что повторные инъекции небольших количеств PEG-PEI-ESO очень эффективны при преобразовании экспрессии дистрофина, достигая до 20% нормальных уровней при самом благоприятном составе и режиме инъекции.
Результаты
Индукция экспрессии дистрофина внутримышечной инъекцией PEG-PEI-ESО
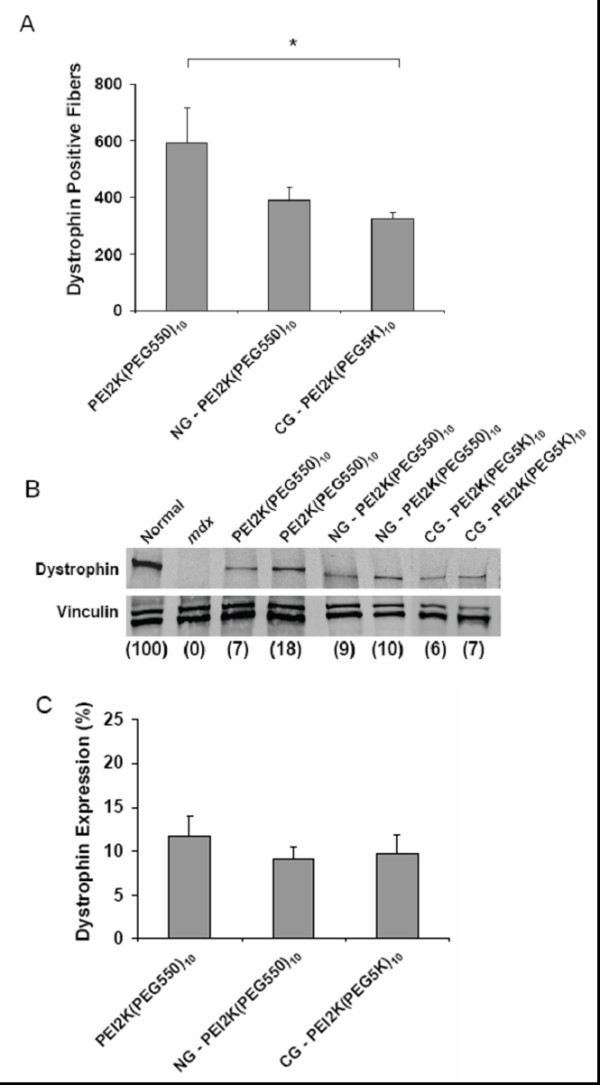
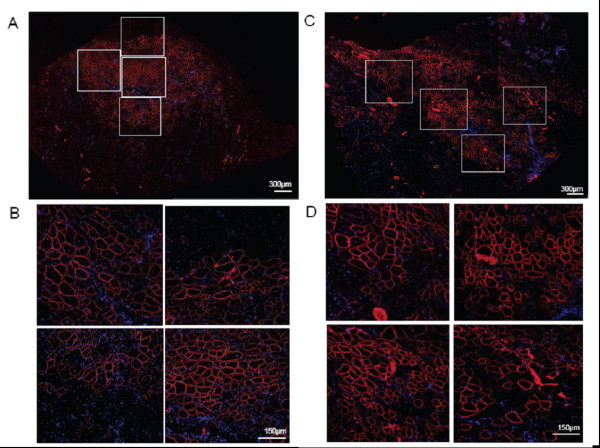
Цель этого исследования состояла в том, чтобы определить эффективный состав PEG-PEI-ESO и схемы инъекции для увеличения экспрессии дистрофина после внутримышечной инъекции в ПББМ 6–8 недельных mdx мышей мужского пола. ESO для всех экспериментов содержал 2'OMe олигонуклеатид, который, как ранее показывали, обладал способностью пропускать экзон 23, таким образом удаляя точечную мутацию, которая приводит к преждевременной остановке кодонов. Мыши получили три еженедельных внутримышечных инъекции 5 нг ESO комплекса или PEI2K (PEG550)10, NG-PEI2K (PEG550)10, или CG-PEI2K (PEG5K)10 кополимеров и были проанализированы на наличие экспрессии дистрофина через одну неделю после третьей инъекции. Иммуногистохимия поперечных секций показала, что все три кополимерных формы произвели существенно большее количество дистрофин-положительных волокон по сравнению с мышцами нелеченных mdx мышей (рисунок 1). Окрашивание гематоксилином-эозином показало морфологическую целостность без явных симптомов некроза мышцы или цитотоксического повреждения (рисунок 1). Количественная оценка целых поперечных сечений мышцы показала, что инъекции PEI2K (PEG550) 10-ESO форма произвела 594 ± 120 дистрофин-положительных волокон (рисунок 2), которые соответствует примерно 30% или приблизительно 2000 волокон в ПББМ [11,43].
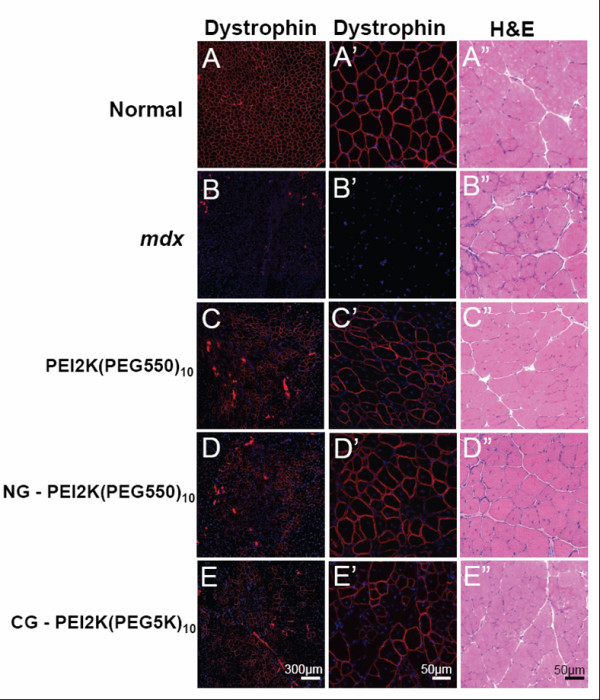
В соответствии с предыдущими исследованиями, показывая улучшил биологическую совместимость и увеличенное клеточное поглощение наночастицконъюгированных с золотом [44,45], мы выдвинули гипотезу, что конъюгация или адсорбция нанозолота (NG) или коллоидного золота (CG) улучшает свойства PEG - PEI кополимеров, как
![]()
носителей. Удивительно, что инъекции NG-PEI2K (PEG550) 10-ESO и CG-PEI2K 10-ESO (PEG5K) заканчивались обнаружением только 380 ± 36 и 322 ± 24 дистрофин-положительных волокон, соответственно, оба из которых были значительно меьше, чем с неконъюгированными PEI2K (PEG550) 10 кополимерами (рисунок 2A; P <0.05).
Чтобы определить количественно экспрессию дистрофина был выполнен Вестерн-блоттинг на поперечной секции целой мышцы, непосредственно на которых было проведено исследование количества волокон. Экспрессия дистрофина после трех еженедельных инъекций PEI2K (PEG550) 10-ESO (5 нг ESO на инъекцию) достигла 11.2 +/-2.4% уровня, найденного в нормальных мышцах (рисунок 2B-C). Как ожидалось, экспрессия дистрофина в контрольной группе mdx мышц не обнаружена, и была, вероятно, меньше чем 1% от нормы. Введение NG-PEI2K (PEG550) 10-ESO и CG-PEI2K (PEG5K), 10-ESO формы заканчивались экспрессией дистрофина 7.6 ± 1.0% и 9.8 ± 2.1% нормального уровня, ни один из которых статистически не отличался от неконъюгированого кополимера (рисунок 2B-C). Хотя мы не выполняли инъекции ESO без полимеров в этом исследовании, мы показали в других исследованиях, что инъекции 2'OMe ESO при подобном дозировании и режимах инъекции как использовалось здесь, произвели очень низкое число дистрофин-положительных волокон и не показал наличие дистрофина при Вестерн-блоттинге ([13] и Sirsi и др.). Точно так же предыдущее исследование, используя ESO, поставленный без носителя, не показало увеличения числа дистрофин-положительных волокон по сравнению с мышцами контроля mdx [43]. Неуместность поставки 2'OMe ESO без носителя у mdx мыши привела к исключению одного только ESO в других исследованиях по индукций синтеза дистрофина после местной и системной поставки [11,16].
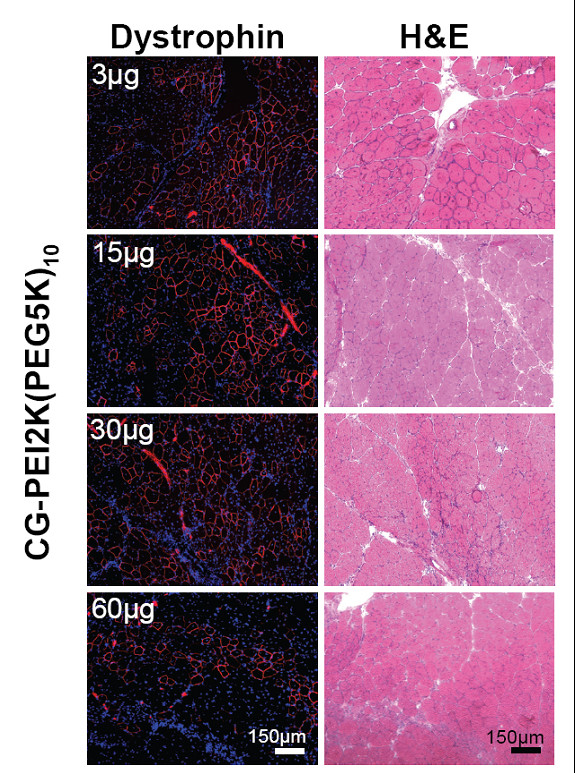
Профиль дозы PEG-PEI-ESO
![]()
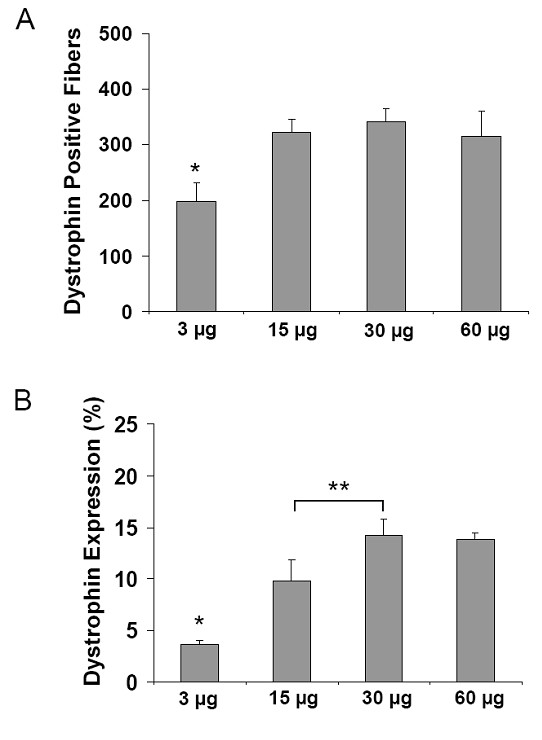
Используя тот же самый режим тройной инъекции как описано выше, свойства дозы CG-PEI2K (PEG5K) 10-ESO были оценены в диапазоне от 3 до 60 нг ESO. Дистрофин-положительные волокна были обнаружены при всех дозах (рисунок 3), но в мышцах с самой маленькой дозой ESO (1 нг Ч 3 инъекции) выражены значительно меньше, чем при введении дозы 15, 30, или 60 нг (рисунок 4A; P <0.05). Вестерн блот анализ показал, что высший уровень индукции синтеза дистрофина был достигнут, используя 5 нг ESO за инъекцию, что закончилось экспрессий 14.2 ± 1.6% на 1.6% дистрофина (рисунок 4B-C).
Долгосрочные повторные инъекции ESO увеличивают экспрессию дистрофина.
![]()
В попытке далее улучшить экспрессию дистрофина, мы выполнили долгосрочные повторные эксперименты по введению NG-PEI2K (PEG550) 10-ESO. Мыши получили 10 последовательных внутримышечных инъекций NG-PEI2K (PEG550) 10-ESO (1 или 5 нг ESO за инъекцию), с промежутком в 4 дня между инъекциями, образцы тканей были собраны через 6 недель после первой инъекции. Иммуногистохимия целых поперечных секций показала, что эта схема лечения приводила к увеличению поставки ESO к мышечным волокнам, производя обширные области дистрофин-положительных волокон (рисунок 5). В среднем мышцы, получившие дозу 1 и 5 нг ESO за инъекцию (10 и 50 нг ESO более чем 6 недель), содержали 832 ± 167 и 1225 ± 343 дистрофин-положительных волокон, соответственно (рисунок 6B). Число дистрофин-положительных волокон после поставки 10 нг ESO в течение 6 недель было 2.2-кратно больше, чем после 15 нг ESO в течение 3 недель, используя тот же самый NG-PEI2K (PEG550) 10. Окрашивание Г-Э подтвердило, что увеличенная экспрессия дистрофина после повторных инъекций, не была связана с явными признаками цитотоксичности (рисунок 6A).
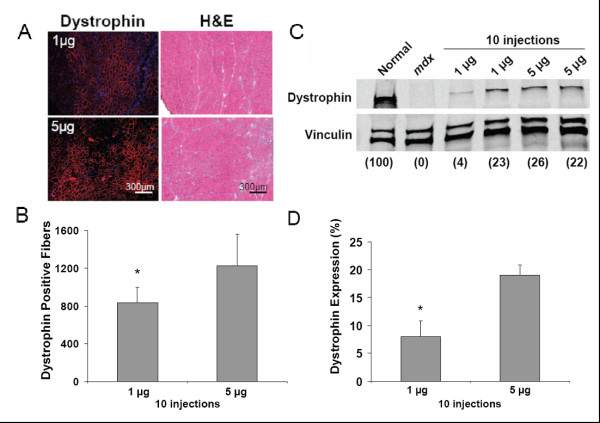
Вестерн-блоттинг установил возможность режима инъекций с 10 повторениями (NG-PEI2K (PEG550) 10-ESO (рисунок 6C-D). В среднем мышцы, получившие 50 нг ESO, имели 19.1 ± 1.8% нормального количества дистрофина, уровень, который, как ранее показывали, был достаточен для выздоровления механических свойств mdx мышцы [43,46,47]. Экспрессия дистрофина после 50 нг ESO была значительно больше, чем 8.1 ± 2.8%, произведенные при 10 нг ESO (P <0.05). Однако, была исключительно высокая вариабельность экспрессии дистрофина в пределах группы 10 нг, самые низкие и самые высокие показатели колебались от 4 до 23%. Пиковый уровень 23% экспрессии дистрофина наблюдался только после введения 10 нг ESO более чем 6 недель, демонстрировал эффективность нанополимер-ESO формы.
Экспрессия nNOS в mdx мышцах, леченных PEG-PEI-ESOs
Мышцы от леченных mdx мышей были проанализированы на экспрессию nNOS как признак экспрессии
![]()
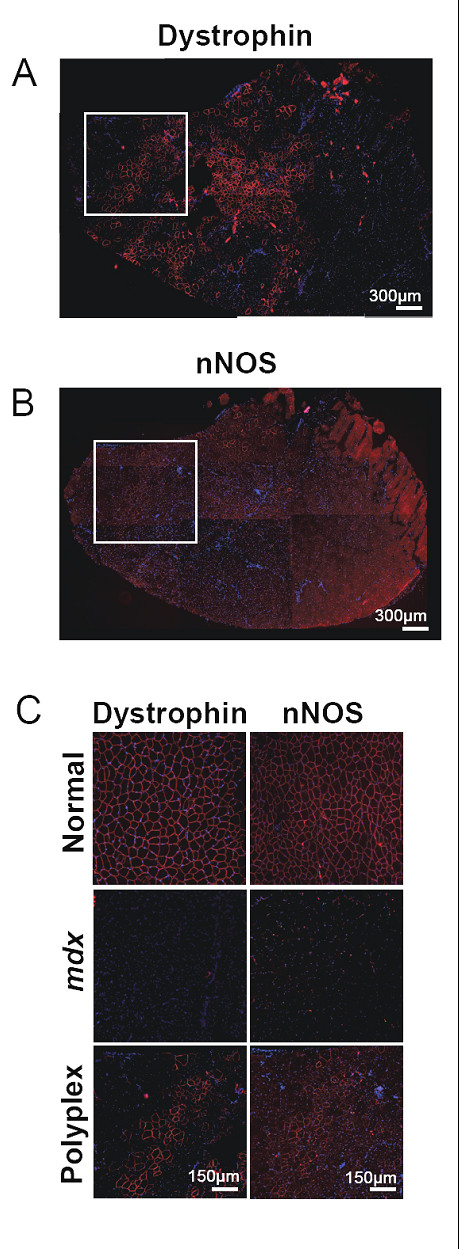
функционального дистрофина с интактной N-терминальной областью. Было ранее показано, что nNOS экспрессия отсутствует в мышцах mdx мышей [48] и может регулироваться положительно после пропуска экзона и восстановления экспрессии дистрофина[11]. Поэтому, мы задались вопросом: увеличила ли текущая стратегия восстановления экспрессии дистрофина уровень nNOS, Исследование последовательных секций на наличие дистрофина и nNOS показали очень высокую корреляцию между дистрофином и nNOS-положительными волокнами (рисунок 7). Специфично, связанный с мембраной nNOS, казалось, был выражен только в mdx мышечных клетках, которые также показали увеличенную экспрессию дистрофина, указывая, что ESO-выраженный дистрофин был функционально интактен. Как ожидалось, nNOS экспрессия отсутствовало в mdx мышцах группы контроля.
Обсуждение
Растущее мнение, что ESO находятся на пути к становлению жизнеспособной терапевтической возможности для лечения МДД, хорошо подтверждается данными исследований клеточных культур и животных, показывая возможность пропуска различных экзонов и проистекающую индукцию синтеза дистрофина почти во всю длину [4,6,7,9,11-16,49,50]. Однако, есть некоторые дебаты, относительно состава ESO, который может работать лучше
![]()
всего, и какой состав носителя обеспечит соответствующую поставку. ESO фосфоротиоат 2'OMe и PMO были лучшим, изученным у моделей животных, и оба в настоящее время участвуют в Фазе I клинического испытания [51]. И 2'OMe и PMO ESO функционируют, блокируя целевые последовательности преМРНК, и считается, что они могут использоваться попеременно, как только оптимальная последовательность (для единственного состава) была эмпирически определена [52]. Однако, принципиальные различия в 2'OMe и PMO устраняют использование универсального носителя для эффективной поставки. Специфично, PMO - синтетические составы, которые чрезвычайно устойчивы к химической деградации, но они - также имеют нейтральный заряд, который ограничивает взаимодействие с поверхностью клеток и клеточное поглощение. Недеградирующая природа PMO также ставит вопросы о их безопасности. Напротив, 2'OMe, ESO - анионная РНК, и несмотря на улучшенную стабильность из-за их фосфоротиоат структуры, остаются несколько восприимчивыми к деградации, в то время как отрицательный заряд препятствует биораспределению и клеточному поглощению.
В этом исследовании мы показали, что кополимеры PEG - PEI с низким MW PEI2K выполняют функцию эффективных носителей для поставки 2'OMe ESO к мышечным волокнам mdx мышей после внутримышечных инъекций, заканчивающихся улучшенными уровнями экспрессии дистрофина. Три еженедельных внутримышечных инъекции только 5 нг ESO с PEI2K (PEG550) 10 заканчивались образованием приблизительно 600 дистрофин-положительных волокон и экспрессией дистрофина на уровне приблизительно 11% нормального уровня через 3 недели после начальной инъекции. Еще более высокие уровни экспрессии дистрофина были достигнуты, используя режим инъекции два раза в неделю, в течение н6 недель. 10 последовательных инъекций NG-PEI2K (PEG550) 10 с 5 нг ESO произвели к образованию более чем 1200 дистрофин положительных волокон и уровню экспрессии дистрофина 20% нормального уровня. В областях с наибольшей концентрацией препарата мы наблюдали сопутствующее увеличение связанного с мембраной nNOS, специфично дистрофин-положительных волокон, в соответствии с предыдущими докладами [11].
Хотя уровни экспрессии дистрофина, показанные в этом отчете, демонстрируют полезность PEG - PEI нанополимеров для поставки ESO, эти составы, кажется, несколько менее эффективны, чем PMO. Alter и др. [14], недавно показал, что единственная внутримышечная инъекци 10 нг PMO заканчивались образованием максимум 60% нормального уровня дистрофина, хотя этот отчет, казалось, обеспечил только оценку эффективности и испытывал недостаток в статистическом анализе достоверности. Другие недавние исследования PMO сопряженным с специфичными пептидами для проникновения в клетки показали внушительные числа дистрофин-положительных волокон, но не обеспечивали полную оценку экспрессии дистрофина вестерн-блотт анализом [12].
PEI2K (PEG550) 10 и PEI2K (PEG5K) 10 кополимеры, используемые в этом исследовании, как ранее показывали, сформировали исключительно устойчивые комплексы с отрицательно заряженным ESO, и поверхностный заряд наночастиц был относительно низок [41]. Мы предлагаем, что высокая стабильность и низкий заряд этих частиц - две существенных особенности, которые делают их более применимыми в естественных условиях для поставки ESO, чем основанные на MW PEI25K кополимеры. Специфично, низкий заряд поверхности частиц улучшает биораспределение и уменьшает цитотоксичность, в то время как высокая стабильность позволяет оставаться связанным во время внеклеточного транспорта.
Наночастицы, содержащие золото, такие как NG и CG, как показывали, улучшили биологическую совместимость и увеличило клеточное поглощение различных типов лекарственных средств [44,45,53,54]. В частности NG, сопряженный с низким MW PEI2K, показал, по крайней мере, на порядок большую эффективность, чем PEI25K и был в 12 раз более мощным, чем немодифицированный PEI2K для поставки ДНК плазмиды в клеточной культуре [44]. Неожиданно, что наши результаты показали, что никакое ковалентное сопряжение NG или CG кополимеров PEG - PEI не улучшило поставку ESO. Пробы стабильности Polyplex в PBS показали, что CG и NG вызвал только умеренное ослабление стабильности (данные, не показаны), делая это маловероятным объяснением из-за отсутствия улучшенной поставки. Возможное объяснение состоит в том, что покрытие CG не было достаточно устойчиво, чтобы прилегать к кополимеру во время поставки. С другой стороны, NG был ковалентно спрягаемым PEG - PEI, и мы постулируем, что его неэффективность происходила более вероятно из-за 1:10 нг отношению к PEI2K, которое, возможно, было слишком низко, чтобы улучшить функциональность.
Экспрессия дистрофина, достигнутая в данном исследовании, была достигнута с носителями PEG - PEI, которые, казалось, не выявляли явных признаков цитотоксичности. Это в отличие от предыдущих исследований с использованием non-PEGylated PEI25K как носителя 2'OMe ESO, который был неэффективен и привел к значительным поврежденииям после очень ограниченного числа инъекций [55,56]. Основываясь на этих результатах, мы пришли к заключению, что катионные полимеры неподходят для естественных условий поставки АО в скелетные мышцы [11,57]. Однако, у частиц PEI-нуклеотида, используемых в этих предыдущих исследованиях несомненно, были очень высокие положительные поверхностные заряды, потому что они не содержали PEG, который, как известно, обеспечивает стерическое ограждение заряда на поверхности PEI. Кроме того, дисперсии чрезвычайно-заряженных-частиц после внутримышечной инъекции, вероятно, препятствуют взаимодействию заряда между PEI и отрицательно заряжеными элементами в пределах внеклеточной среды. Поэтому, не удивительно, что эти предыдущие исследования трансфекции мышцы non-PEGylated PEI приводили к неудовлетворительным результатам. Мы предполагаем, что комбинация MW PEI и PEGylation, используемого теперь, обеспечила благоприятную форму, которая была и эффективна и нетоксична. Однако, нехватка наблюдаемой цитотоксичности не устраняет возможность, что некоторое повреждение мышцы может встречается после инъекции, заканчивающейся некоторым уровнем регенерации дегенерации. Этот процесс может лежать в
![]()
основе до некоторой степени высокого числа дистрофин-положительных волокон, наблюдаемых в нашем 6-недельном (10 инъекций) испытания. Хотя не было систематически оценки, мы заметили, что кратковременное механическое повреждение встречается в mdx мышцах после внутримышечных инъекций различных составов (даже солевой раствор), который не встречается в нормальной мышце. Из-за нехватки дистрофина mdx мышцы более восприимчивы к механическому повреждению, чем нормальная мышца. Этот эффект может быть усилен до некоторой степени катионными частицами, или в этом отношении, любым типом состава носителя.
Мы предполагаем, что главное ограничение форм носитель-ESO, описанных в этом отчете, было в несоответствующей функциональностьи носителя, и свойственной силой ESO. ESO, используемый в данном исследовании (определяемый в литературе как M23D (+02–18)), как показывали, in vitro преобладающе производил пропуск экзона 23, хотя некоторый пропуск экзонов 22–23 действительно встречается [42]. Кроме того в этом исследовании мы показали, что при самом эффективном условии, приблизительно 50% волокон были дистрофин-положительны, что соответствовало 20% нормальной экспрессии дистрофина. Это указывает, что в средних дистрофин-положительных волокнах содержалось приблизительно 40% нормального уровня дистрофина. Подобное вычисление, основанное на данных, о которых сообщает Lu и др. [11], и наше недавнее исследование с, TAT - сопряженными кополимерами (Sirsi и др.,) предполагают, что уровень дистрофина может достигнуть 75% нормальных уровней.
Таким образом главное ограничение катионных носителей, как кажется, их плохое диффузионное распределение, как обнаружено, большими областями в мышцах без очевидной трансфекции. Таким образом дальнейшее совершенствование функциональности носителя, вероятно, потребуется, чтобы позволить использование в клинические условия для лечения МДД. Наша группа недавно показала, что сопряжение множественного HIV-TAT эпитопа с PEI2K (PEG5K)10 очень улучшили поставку ESO, используя подобное дозирование и подобный режим внутримышечных инъекций, заканчиваясь максимум 30% экспрессии дистрофина (Sirsi и др.,). Различные другие типы нацеленных на клетку лигандов, пептидов проникновения клетки могут также связываться с PEI, чтобы улучшить функциональность. Значительно, что этот тип сопряженного пептида-PEI может, вероятно, быть синтезирован для улучшения системной поставки, которая потребуется, чтобы достигать значительного терапевтического эффекта. PMO, как показывали исследования, ограничили эффективность после системной поставки. Например, интраперитонеальные инъекции у новорожденных mdx мышей 5 мг/кг/нед. PMO-пептида произвели широко распространенную экспрессию дистрофина в мышце диафрагмы с низкими уровнями экспрессии, наблюдаемыми в мышцах конечности [12,58].
Методы
Животные
Мdx мыши мужского пола (C57BL/10ScSn-Dmdmdx/J) и нормальные мыши в возрасте 6–9 недель(C57BL/10SnJ) приобретены в Лаборатории Джэксона (Bar Harbor, ME). Все животные использовались в соответствии с NIH и университетскими правилами (Drexel University College of Medicine, ULAR facility, Philadelphia, PA).
Синтез нанополимера
Синтез и физико-химическая характеристика PEI2K (PEG550) 10 и PEI2K (PEG5K) 10 кополимеров были ранее описаны [41,59]. Кополимеры определяются, используя спецификацию, где приписка указывает число цепей PEG, трансплантированных на молекулу PEI. Например, PEI2K (PEG550) 10 указывает 10 цепей PEG 550 дальтон, трансплантированных каждой молекуле PEI на 2 килодальтона. Частицы нанозолота(NG) спрягались с основными группами амина PEI на PEI2K (PEG550) 10 с использованием Сульфо-N-гидрокси-сукцинимидо нанозолота (Nanoprobes, Yaphank, Нью-Йорк). В этой реакции 75 нмолей PEI2K (PEG550) 10 были смешаны с 6 нмолями NHS нанозолота в 780 мкл стерилизованной воды (pH фактор = 8.0). Результат заморожен в течение 24 часов на льду и впоследствии высушен сублимацией и сохранялся при -20°C. Адсорбция коллоидного золота (CG) PEI2K (PEG5K) 10 была выполнена, смешивая 300 мкл частиц CG 5 нм (Sigma-Aldrich) с 30 мг PEI2K (PEG5K) 10 (в 1 мл DI H20) при 4°C быстро. Раствор полимера впоследствии сушился сублимацией и хранился при -20°C. NG и маркированные кополимеры CG определяются как NG-PEI2K (PEG550) 10 и CG-PEI2K (PEG5K) 10, соответственно.
Подготовка PEG-PEI-ESO
ESO, используемый во всех экспериментах, содержал 20-mer олигонуклеатидов (5 '-GGCCAAACCUCGGCUUACCU-3'), пропускал экзон 23 из гена дистрофина у mdx мышей [11,16,42]. Во время синтеза (Trilink, Сан-Диего, Калифорния) каждое основание было соединено с фосфоротиоатом и содержало метокси группу у 2' углерода. PEG-PEI-ESO были подготовлены в азоте для соединения с фосфатом 5 (N:P = 5); где N представляет заносы амина на PEI, и P представляет заносы фосфата на ESO. Комплексы были сформированы добавлением кополимерного PEG - PEI к ESO (в стерильном солевом растворе).
Внутримышечные инъекции PEG-PEI-ESO
Мdx мыши мужского пола (6–9 недель возраста) были анестезированы и сбривалась шерсть для визуализации мышц задней конечности. 15 мкл объемов PEG-PEI-ESO раствора в различных концентрациях были введены с двух сторон в середину части брюшка ПББМ, используя 31 инсулиновый шприц. После восстановления после анестезии мыши были возвращены к нормальной активности.
Мы использовали и 3 и 6 недельный график инъекций следующим образом. Для групп с 3 недельным графиком мыши получили инъекции в 0, 7, и 14 дни; и образцы были собраны на 21. В одной серии экспериментов в мышцы были введены 5 нг ESO (15 нг общих количеств) или PEI2K (PEG550) 10, NG-PEI2K (PEG550) 10, или CG-PEI2K (PEG5K) 10 кополимеры. Для анализа реакции на единственную дозу в мышцы были введены 1, 5, 10, или 20 нг
![]()
ESO (3, 15, 30, и 60 нг общих количеств) CG-PEI2K (PEG5K) 10 кополимеров. Для групп с 6 недельным графиком в мышцы был введен препарат в 0, 4, 8, 12, 16, 20, 24, 28, 32, и 36 день; и образцы были собраны на 42. Для этого эксперимента в мышцы были введены или 1 или 5 нг ESO за инъекцию (10 и 50 нг общих количеств) с NG-PEI2K (PEG550) 10 кополимеров. Для всех групп были проанализированы 2 мыши (N = 4 мышцы).
Иммуногистохимия, гистология и количество волокон.
В определяемых моментах времени были убиты мыши, и удалены ПББМ, заморожены жидким азотом в изопентане, так сохранились при -80°C. Мышцы контроля были собраны от нелеченных подобранных по возрасту mdx и нормальных мышей. Поперечные замороженные секции (10 мкм) были вырезаны из брюшка каждой мышцы используя криостат (Leica CM 3050 S, Бэннокберн, Иллинойс) и оттаяны для иммуногистохимии и гистохимии. Толстые (60 мm) поперечные секции, немедленно смежные с тонкими срезами, были разрезаны в криостате и помещены в трубки для центрифуги на 1.5 мл на сухом льду и сохранились при -80°C для последующего западного анализа.
Для иммунного анализа секции мышцы были обработанны 10%-ой нормальной сывороткой козы в 1%-ом BSA/PBS в течение 1 ч и затем в течение 1 ч поликлональными антидистрофиновыми антителами кролика (1:200; Abcam, Кембридж, Массачусетс) или анти-nNOS (1:125; Invitrogen, Карлсбад, Калифорния). Вторичное антитело было IgG Cy3-Anti-Rabbit (1:500; Jackson Immunoresearch, Западная Роща, Пенсильвания). Слайды были помещены в заливочную среду Vectashield с DAPI (Векторные Лаборатории, Берлингейм, Калифорния) и получены изображения (4/10/20Ч; Олимп, AX70, Мелвилл, Нью-Йорк).
Вестерн блот анализ экспрессии дистрофина.
Из секции замороженных ПББМ (60 мm) проведена экстракция путем добавления 1.5 мл к 50 мк белкового буфера, содержащего 125 мм Tris (pH фактор 6. 8), 4%-ого SDS, Экстракт был заморожен на льду (15 минут), кипел (5 минут) и центрифугирован (4000 Ч g в течение 5 минут). Белковая концентрация была измерена в экстрактах, используя пробу Coomassie (Проникните, Рокфорд, Иллинойс) и равный объем буфера SDS-PAGE (125 мм Tris (pH фактор 6. 8), 4%-ый SDS, 5%-ый 2-mercaptoethanol, 10%-ый глицерин и 0.05% бромфенола синего), был добавлен к экстракту. Образцы, содержащие 25 нг общего количества белка, были помещены в гели SDS (3% stacking: 7.5% resolving; Bio-Rad, Hercules, CA) и после 16 ч, окрашивались Ponceau S (Сигма, Сент-Луис, Миссури), чтобы визуализировать результат. Мембраны были в течение 1 ч обработанны моноклональными антидистрофиновыми антителами (MANDYS8; Сигма, Сент-Луис, Миссури) в разведениях 1:400 и 1:2000. Вторичные антитела осла против мыши IRDye, 800 (Биологические науки ЛИТИЯ БОЖЕ МОЙ, Lincoln, Небраска) были применены в течение 1 ч и мембрана была просмотрена с помощью Системы Отображения после многократного мытья TBS-T/TBS.
Статистический анализ
Все данные представлены в виде значение± SEM. Статистическия анализ выполнен методом ANOVA (Statview; SAS Institute, Cary, NC).
Оригинал статьи: http://www. /1472-6750/8/35
Переведено проектом МОЙМИО http://www. mymio. org/
![]()
