
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ГБОУ ЛГК на Юго-Востоке
Московский Химический Лицей 1303
Институт биоорганической химии им. и
Создание флуоресцентного сенсора на поли-АДФ-рибозу
(исследовательская работа)
Выполнила ученица 11 «В» класса Ипатова Дарья
Научные руководители: , , .
Введение
В молекулярной биологии использование флуоресцентных белков стало удобным подходом к визуализации внутриклеточных процессов. В частности, с их помощью можно отслеживать изменение активности ферментов и концентраций исследуемых веществ в клетках и тканях.
Целью нашей работы стало создание флуоресцентного сенсора для определения поли-(АДФ-рибозы). Поли-(АДФ-рибоза) участвует во многих внутриклеточных процессах, таких как например, укладка хроматина, регуляция транскрипции и репарация ДНК. Нашей задачей было изучение роли поли-(АДФ-рибозы) в цикле репарации ДНК. Сенсоры для визуализации ПАР в живых клетках ранее описаны не были.
Литературный обзор
Поли (АДФ)-рибоза (рис.1) (ПАР, PAR) обнаружена в клетках у всех эукариот (кроме дрожжей). Процесс синтеза поли(AДФ-рибозы) начинается в ядре ответ на повреждения ДНК и предшествует началу ее репарации. ПАР образуется из НАД+ под действием фермента поли(АДФ-рибоза)полимеразы. Таким образом, образование ПАР изменяет энергетический баланс клетки. Большие затраты энергии на биосинтез этого полимера указывают на его важную роль. Известно, что при увеличении концентрации ПАР до критического уровня, образующийся в ядре ПАР не успевает деградировать, остатки ПАР выходят в цитоплазму, связываются с AIF, индуктором апоптоза, и таким образом запускается процесс программируемой клеточной смерти.
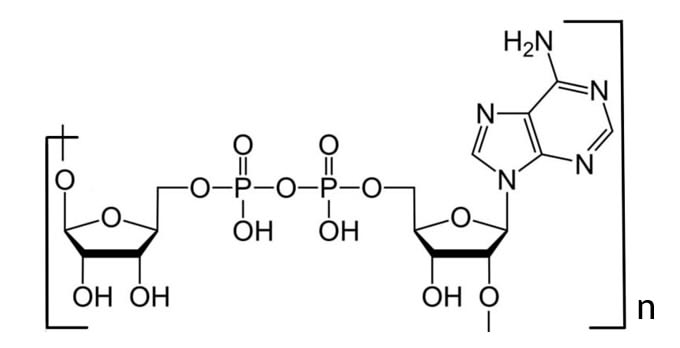
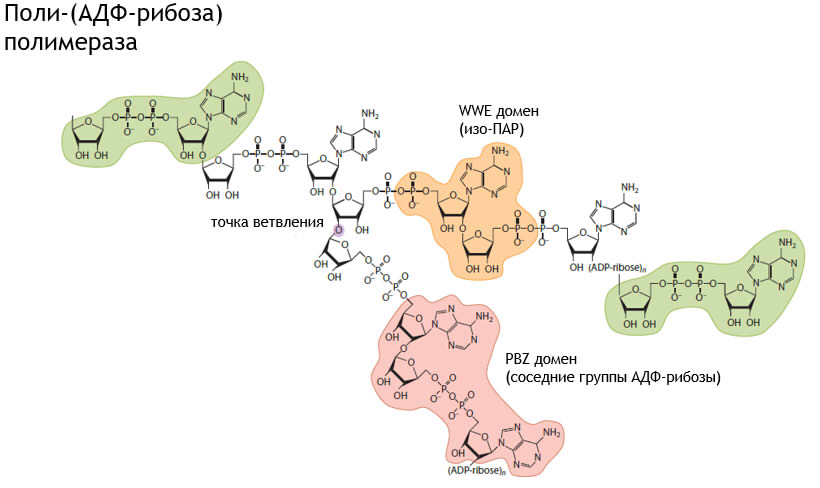
Несмотря на обширные исследования, роль и механизм действия ПАР в процессе репарации ДНК остается до конца неизученным из-за структурной сложности и разнообразия. Недавнее открытие и определение строения четырех разных ПАР-связывающих мотивов представляют огромные возможности для более подробного изучения молекулы. (рис.2-механизм распознавания и прикрепления доменов к ПАР)
(Luo, X., & Lee Kraus, W. (2012). On par with PARP: Cellular stress signaling through poly(ADP-ribose) and PARP-1. Genes and Development, 26(5), 417–432. https://doi. org/10.1101/gad.183509.111
Schreiber, V., Dantzer, F., Ame, J.-C., & de Murcia, G. (2006). Poly(ADP-ribose): novel functions for an old molecule. Nature Reviews. Molecular Cell Biology, 7(7), 517–28. https://doi. org/10.1038/nrm1963)
- PBM (PAR-bindingmotif).
Консенсусная последовательность PBM включает в себя 8 аминокислот: [HKR]1-X2-X3-[AIQVY]4-[KR]5-[KR]6-[AILV]7-[FILPV]8. Белки, содержащие данный ПАР-связывающий мотив участвуют во многих биологических процессах, например, в репарации ДНК, укладке хромосом, синтезе РНК и регуляции клеточного цикла. Несмотря на то, что PBM-первый описанный ПАР-связывающий мотив и были представлены 3D структуры некоторых PBM-содержащих полипептидов, механизм взаимодействия PBM с поли-(АДФ-рибозой) не известен. В PBM входят положительно заряженные аминокислоты, в следствие этого предположительно происходит связывание с отрицательно заряженными фосфатными группами ПАР. Макродомен
Впервые макродомен, проявляющий ПАР-связывающую активность, был выделен из белка Af1521 термофильного организма Archaeoglobusfuldigus. В организме человека 10 генов кодируют 11 белков, содержащих макродомен. Он способен связываться не только с ПАР, но и с ADPR (АДФ-рибоза), О-ацетил-АДФ-рибозой, синтезирующейся в процессе NAD+-зависимой деацетиляци иацетилированных белков. Цинковые пальцы (PBZ)
С2Н2 цинковый палец, способный эффективно связываться с ПАР (распознавая соседние группы АДФ-рибозы), был выделен из супрессора опухоли CHFR. Консенсусная последовательность мотива – [K/R]xxCx[F/Y]GxxCxbbxxxxHxxx[F/Y]xH. Мотив был найден только в двух белках млекопитающих: гистоновый шаперон и днк-репарирующий белок APLF (aprataxinandPNK-likefactor) и промежуточно сшивающий репарирующий белок SNM1. WWE
Последний открытый ПАР-связывающий мотив-WWE. WWE содержит 170 аминокислот, назван по самым консервативным – двум триптофанам и глутаминовой кислоте. Он был найден в белках, участвующих в поли-(АДФ-рибозилировании) или убиквитинировании. WWE, выделенный из белка RNF146 убиквитиновой Е3-лигазы, проявляет ПАР-связывающую активность, распознавая уникальную гликозидную связь рибоза-рибоза (т. н. изо-ПАР
Флуоресцентные белки-белки, способные при облучении их светом определенной длины волны светиться в видимом диапазоне. Первым был открыт белок GFP (green fluorescent protein) выделенный из медузы Aequoreavictoria. исследователем Осаму Симомурой в 1962 году (схема-рис.3). А в 2008 Осаму Симомура, Мартин Чапфи, Роджер Тсьен получили Нобелевскую премию по химии «за открытие и разработку зелёного 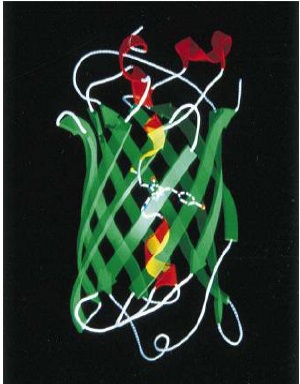 флуоресцентного белка GFP»...
флуоресцентного белка GFP»...
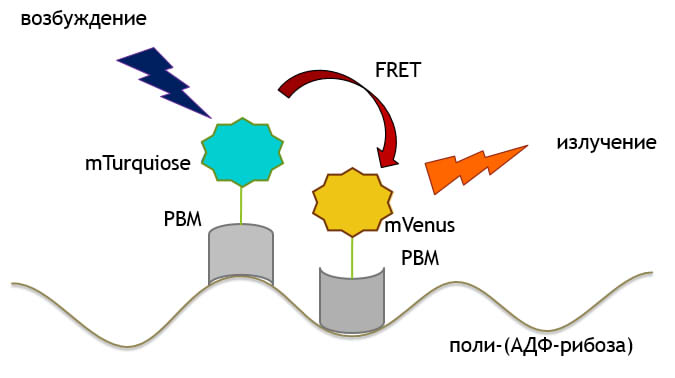
Пространственная структура GFP и подобных ему флуоресцентных белков очень устойчива и представлена в-бочкой, сформированной 11 в-листами и б-спиралью. На основе GFP были созданы другие флуоресцентные белки, в частности белки mTurquoise ( светящийся в синем диапазоне) и mVenus (светящийся в желтом диапазоне).
Между двумя флуоресцентными белками может возникнуть FRET-Fцrster resonance energy transfer (фёрстеровский резонансный перенос энергии). Это процесс, в котором принимает участие два флуоресцентных белка: донор и акцептор переноса (рис.4). Во время FRET происходит перенос энергии от одного белка к другому. То есть, возбуждая одну молекулу (донор), можно наблюдать флуоресценцию с другой (акцептора). Важно, чтобы спектр флуоресценции донора перекрывался со спектром поглощения акцептора. (рис.5) Известно, что димеризация донора и акцептора повышает эффективность FRET.
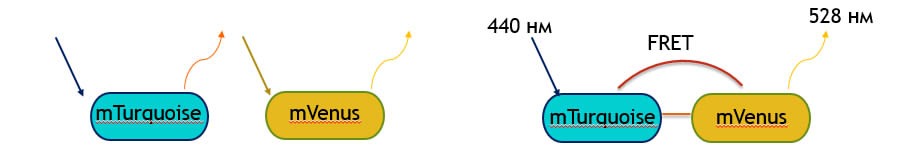
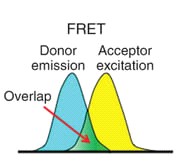
Белки mTurquoise и mVenus-известная FRET-пара.
Задачи:
- Амплификация последовательности PBM-mTurquoise(рис.7)с помощью step-outPCR (полимеразная цепная реакция) Клонирование вставки (пцр-продукт PBM-mTurquoise) в вектор (mTurquoise(206K) и mTurquoise (K206A) ) для экспрессии в клетках млекопитающих(рис.8) Отбор колоний, содержащих вставку Детекция экспрессии PBM-mTurquoise в клетках млекопитающих с помощью флуоресцентной микроскопии
Методы:
- Step-out PCR Обработка эндонуклеазами рестрикции Очистка смеси с помощью микроколонок. Переосаждение Лигирование Трансформация бактерий Скрининг (аналитическая ПЦР с колоний) Флуоресцентная микроскопия
Ход работы:
Мы амплифицировали последовательность, кодирующую белок mTurquoise(206A или 206K),содержащую на N-конце PBM.(рис 9)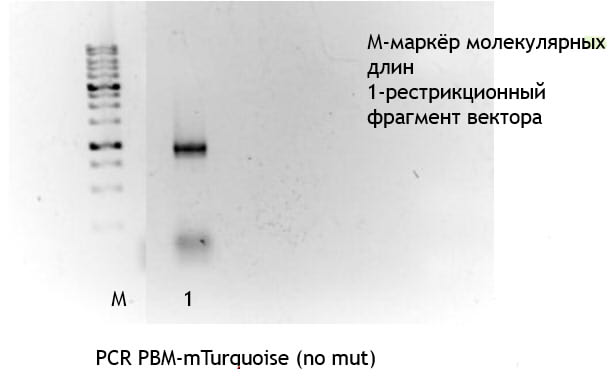

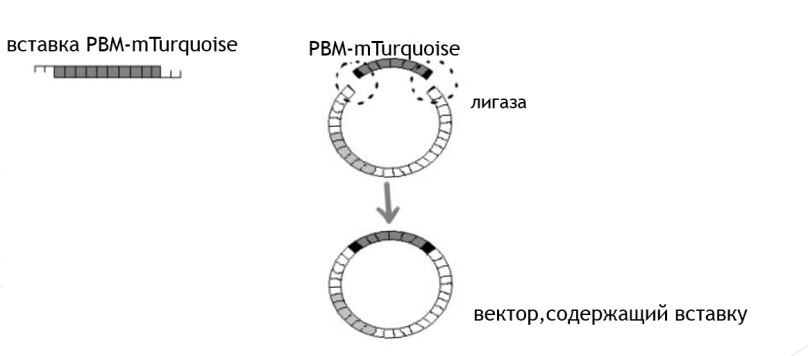
4)Мы провели трансформацию бактерий-внесение полученной плазмиды в бактериальные клетки. Клетки, поглотившие плазмиду, приобретают устойчивость к антибиотику, следовательно, могут вырасти на чашке Петри, содержащей питательную среду с антибиотиком.
5)Мы внесли полученную конструкцию в клеточную культуру млекопитающих-клетки аденокарцинома шейки матки HeLa
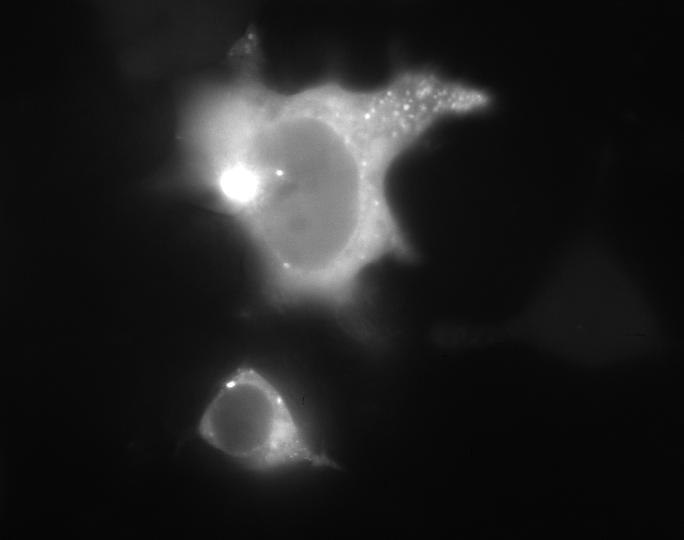
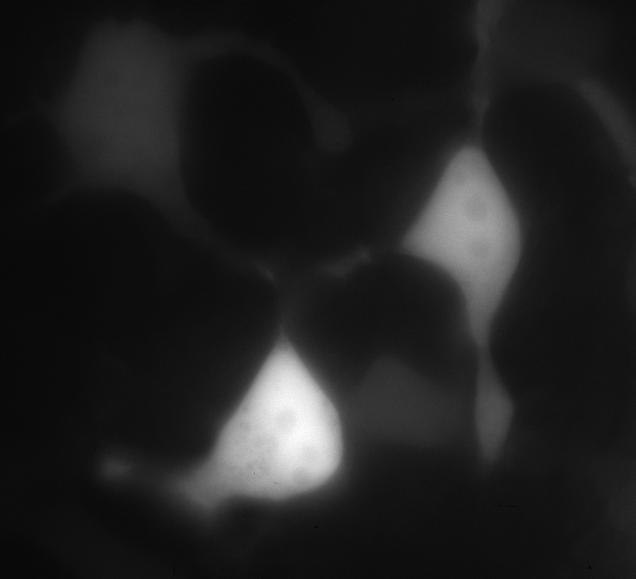
6)Мы детектировали экспрессию PBM-mTurquoise в клетках HeLa с помощью флуоресцентной микроскопии(рис.12-13)
7) Мы добавили к клеточной культуре пероксид водорода для индукции клеточного стресса.
Химерный белок PBM-mTurquoise экспрессировался в живых клетках, но не транслоцировался в ядро после добавления пероксида водорода, возможно из-за того, что одного PBM-домена недостаточно для связывания ПАР(рис.14-15). Параллельно в лаборатории была получена генетическая конструкция mVenus-WWE (WWE-мотив, способный селективно связываться с ПАР) Химерный белок mVenus-WWE также экспрессировался в живых клетках, и при добавлении оксида водорода наблюдалась транслокация флуоресцентного сигнала в ядро (рис.16-17).
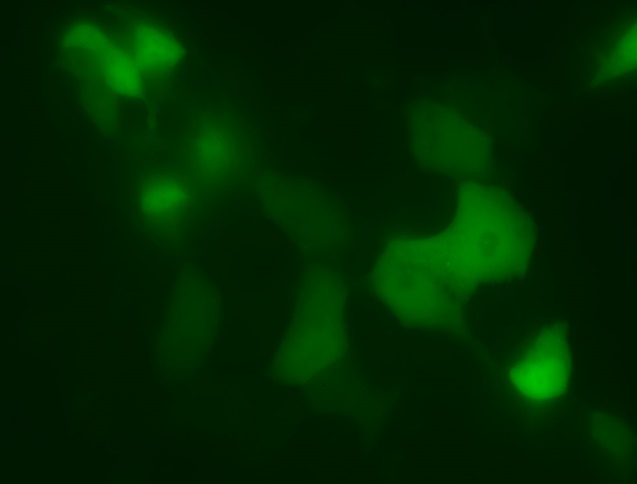
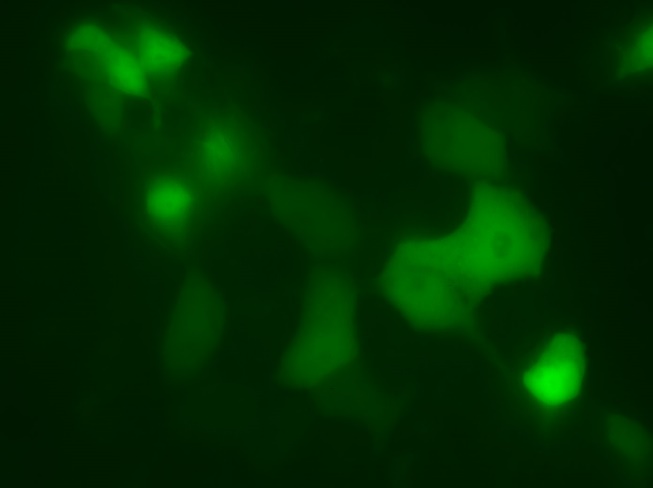
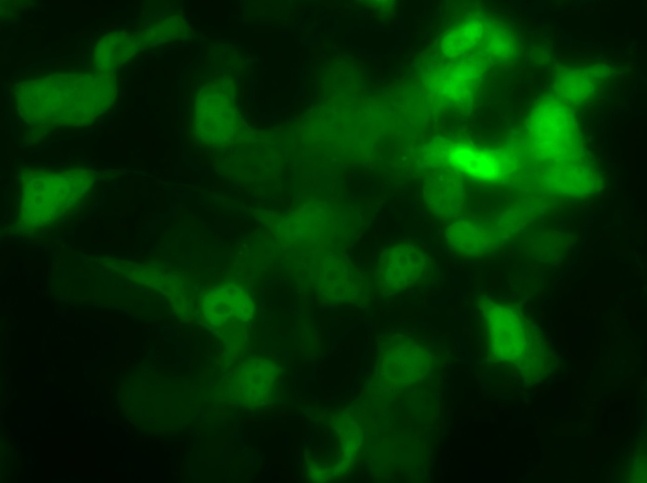
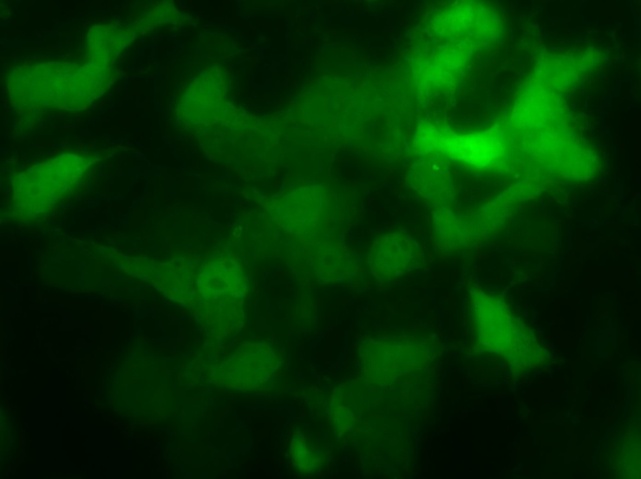
Выводы:Мы синтезировали генетические конструкции, кодирующие PBM-mTurquoise (206K) и(K206A), mVenus-WWEи mTurquoise-WWE.
Полученные конструкции способны экспрессироваться в клетках млекопитающих.
Возможно, одного PBM-домена недостаточно для связывания ПАР, из-за чего мы не наблюдаем транслокацию при индукции клеточного стресса.
Получены химерные белки, содержащие мотив WWE, которые могут быть использованы в качестве генетически кодируемых сенсоров ПАР.
Дальнейшие планы:Подбор оптимальных условий для регистрации сигнала FRET-сенсора. Изучение изменения концентрации ПАР в живых клетках млекопитающих под влиянием индукторов клеточного стресса.
