

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

Глюкозорегулируемый белок 94 участвует в дифференцировке мышц через контроль аутокринного производства инсулин-подобного фактора роста.
Введение
Ряд доказательств указывают на важную роль GRP94 ER шаперона в мышечной физиологии. Во-первых, GRP94-/ - ЭС клетки не могут дифференцироваться в любую из мышечных суб-линий, и снижение уровней GRP94 в скелетных миобластах приводит к потере миоцитов. Во-вторых, экспрессия GRP94 повышается в течение внутриутробного развития кролика и отображает различные паттерны экспрессии в скелетных и сердечной мышцах [3]. В-третьих, GRP94 функционирует как стрессовый белок в мышцах: увеличение его экспрессии наблюдалось временно в миоцитах предсердий и сверхэкспрессия GRP94 в напряженных кардиомиоцитах защищает их от гибели. Стресс-защитная роль специфична для GRP94 и не свойственна другим шаперонам эндоплазматического ретикулума таким как BiP или кальретикулин. GRP94 может оказывать свое воздействие на мышечные клетки благодаря своей важной роли в производстве IGF, от деятельности которых зависят мышечные волокна и, или потому, что он имеет значение для гомеостаза кальция в возбужденных клетках. Мы были особенно заинтригованы возможностью GRP94 значения для роста мышц через его шаперонную активность по отношению к IGF. Сверхэкспрессия инсулиноподобного фактора роста-II в мышиных эмбриональных стволовых клетках способствует их миогенной дифференцировке. Локализованная IGF-I экспрессия повышает мышечную гипертрофию, поддерживает гипертрофию и регенерацию стареющих скелетных мышц, что ускоряет регенерацию мышц, возможно и у MDX мышей. Наконец, мышца производит специфические изоформы IGF-I, функция которых пока неясна. Эта работа показывает, что активность GRP94 действительно важна для миогенной дифференцировки клеток и, что важен GRP94 на промежуточной фазе миогенеза, потому что он способствует производству локально действующих IGF.
Результаты
Отсутствие GRP94
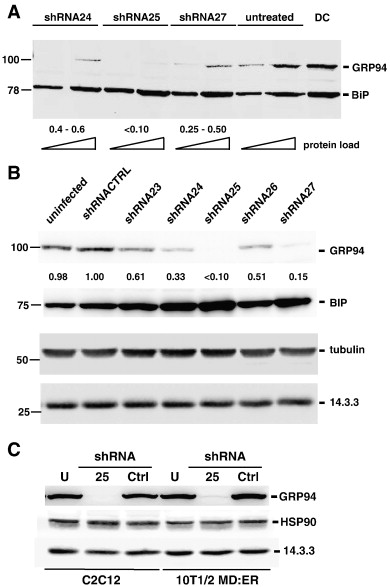
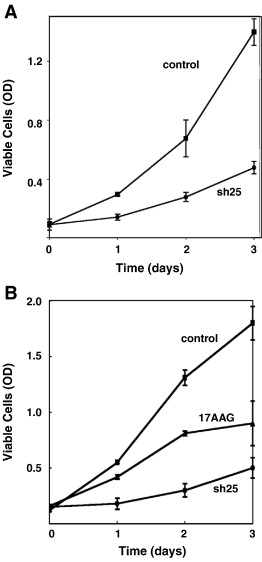
Чтобы проверить механизм GRP94 действия во время развития мышц, мы использовали два дополнительных инструмента: понижающую регуляцию экспрессии GRP94 с РНК-интерференцией и фармакологическое ингибирование деятельность 17AAG. Из семи потенциальных последовательностей, предсказанных алгоритмов иРНК, три ингибируют экспрессию GRP94 (shRNA24, 25 и 27). Как показано на рис. 1A, три ShRNA вектора в клетках С2С12, с SH25, наиболее эффективныв в блокаде GRP94 (обычно> 90% нокдаун), sh24 частично эффективны (примерно 60% нокдаун) и SH27 занимают промежуточный уровень между ними. Для эффективной блокады через трансфекцию требуется более 60 ч после трансфекции, в соответствии с длительным периодом полураспада от GRP94 (> 48 часов). Из-за плохой трансфектабельности C2C12, мы впоследствии доставили shRNAs с помощью лентивирусной инфекции, которая оказалась высокоэффективным методом блокады GRP94 (рис. 1б). Специфика ShRNA реагентов к GRP94 демонстрирует их неспособность блокировать BiP (78 кДа). Оба GRP94 и BiP совместно обнаружены при вестерн-блот анализе при использовании анти-KDEL моноклональных антител (фиг. 1А). Специфика блокады также демонстрирует неспособность анти-GRP94 РНК-интерференции для подавления экспрессии тубулина.
3.2. Ингибирование GRP94 в миобластах снижает мышечную дифференцировку.
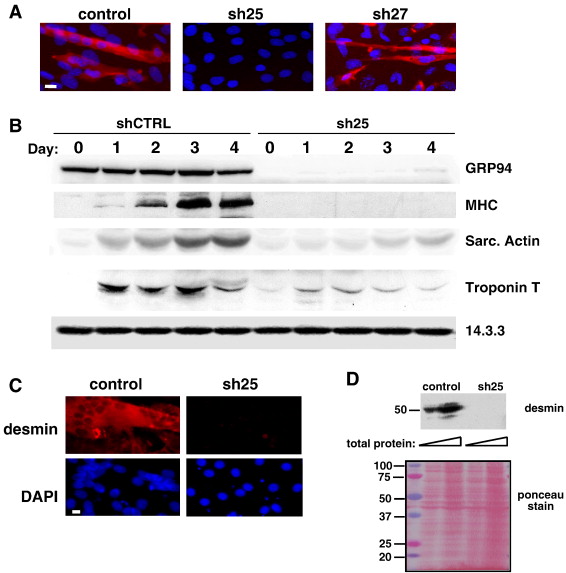
Когда GRP94 уровни были истощены, способность клеток С2С12 дифференцироватся ингибируется. Меньше клеток сливаются в мышечных трубочки - синцитий (рис. 2а), меньшее число клеток экспрессирует тяжелую цепь миозина (рис. 2а) или десмин (рис. 2в), а общее количество сократительных белков, продуцируемых клетками резко сократилось: индукция экспрессии тяжелой цепи миозина, саркомерного актина, тропонина Т и десмина ингибируется (фиг. 2B, D). Тяжесть дефекта дифференцировки коррелирует со степенью блокады в нескольких экспериментах, обеспечивая хорошее доказательство того, что С2С12 миогенная дифференцировка пропорциональна активности GRP94.
Ингибирование как миогенной дифференцировки, так и IGF секреции были подтверждены фармакологически, а не генетическим GRP94 ингибированием активности. При обработке 17AAG, C2C12 клетки не экспрессировали тяжелой цепи миозина (рис. 4D), и не дали значительных уровней IGF-II (рис. 3а). В то время как 17AAG дает преимущество быстрого анализа, это менее конкретный агент, потому что он ингибирует цитозольный HSP90 так же эффективно, как GRP94. Сочетание эффектов препарата и фенотипа иРНК, однако, подтверждает, что GRP94 имеет важное значение для миогенеза из-за его шаперонной активности по отношению к IGF.
![]()
3.3. Комплементация с экзогенным IGF
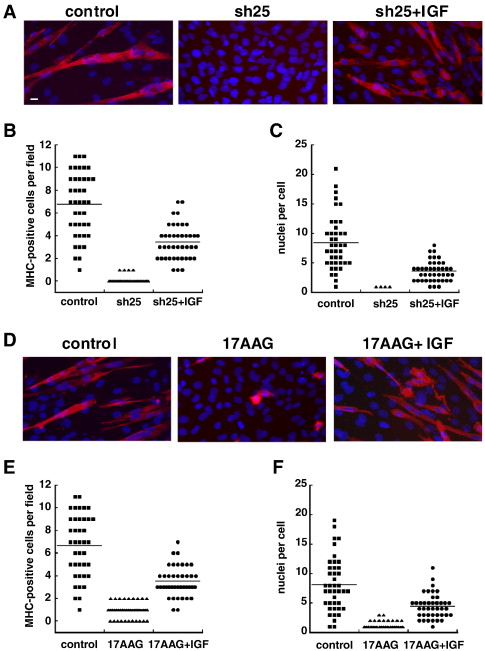
Важно отметить, что добавление экзогенного IGF к миобластами с блокадой GRP94 частично восстановило дифференцировку. Экспрессия сократительных белков и слияние клеток были восстановлены в присутствии IGF. Либо рекомбинантный IGF-I или рекомбинантный IGF-II были эффективны в восстановлении миогенеза, в соответствии с общим количеством рецептора IGF-IR и долей двух гормонов. C2C12 и 10T1/2MD: ER производили IGF, но гораздо больше IGF-II (рис. 3). Следует отметить, что восстановление миогенеза экзогенным IGF является неполным: среднее количество MHC-позитивных клеток при наблюдении за ростом поля от 0 до приблизительно 4 в присутствии IGF-II, по сравнению с приблизительно 7 в необработанных клетках С2С12 ( Рис. 4В). Аналогичным образом изменялись параметры: число клеток, слитых с получением миотубы, среднее число 4 ядер в мышечных трубочках в присутствии IGF, по сравнению с 8 в контрольных миотубах (рис. 4С). Восстановление может быть неполным, поскольку мы не достигли оптимальной концентрации активных IGF, или, скорее всего, потому, что другие GRP94, участвуют в миогенезе. Кроме того, количественное соотношение между уровнем IGF-II в среде, и качеством мышечной дифференциации последовательно наблюдается в нескольких экспериментах (например, рис. 3 и рис. 4). Таким образом, мы приходим к выводу, что основной причиной торможения мышечной дифференциации при блокаде GRP94 является неспособность миобластов производить IGF. Как мы показали ранее, GRP94 является обязательным шапероном для производства компетентных IGF-I и-II, и что он связывается с ER формами про-гормонов, прежде чем они будут обработаны протеолитически в зрелые факторы роста [6].
3.4. GRP94 деятельность необходима для промежуточных шагов при миогенной дифференцировке.
Затем мы спросили, является ли влияние GRP94 распространенным на всю программу миогенной дифференцировки, или же оно необходимо на определенной стадии дифференцировки. Важно отметить, что блокада экспрессии GRP94 не помешала индукции экспрессии MyoD, фактора транскрипции, который инициирует миогенную программу. Это было показано при иммуноблоте (рис. 5а) и иммунофлюоресценции (рис. 5б).
![]()
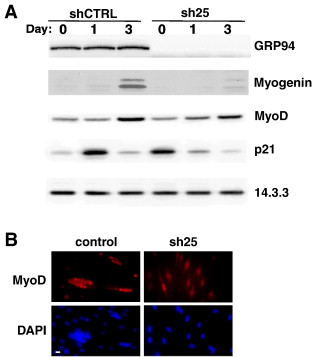
В отличие от MyoD индукция экспрессии другого специфического для мышц фактора транскрипции, миогенина, ингибируется в GRP94-дефицитных клетках С2С12 (фиг. 5А). Неспособность экспрессировать миогенин не является исключительной собственностью C2C12, потому что миогенин также обнаруживается в мышечных дифференцирующихся культурах, полученных из GRP94-/ - ЭС клеток. Даже в 10T1/2MD: ER клетки, которые сверхэкспрессировали MyoD, индукция экспрессии миогенина ослабляется блокадой GRP94 (данные не показаны). Это согласуется с известным синергизмом, который приводит к активации промоутера миогенина, который отражает комплексные действия MyoD и от других факторов роста. В самом деле, родительская клетка 10T1 / 2 линии, которая не выражает MyoD, не может дифференцироваться и выделяет лишь незначительное количество IGF-II (0,94 ± 0,013 нг / мл через 48 ч, по сравнению с 30 нг / мл выделяемого 10T1/2MD: ER( см. рис 3В).
3.5. Истощение GRP94 не вызывает ER стресса и не меняет апоптоза.
Сайленсинг GRP94 в C2C12 или 10T1/2MD: ER клеток приводит к повышенной регуляции экспрессии BiP (рис. 1, 6.). Это явление не похоже на типичную реакцию на стресс ER. Во-первых, величина индукции BiP является субмаксимальной, как можно видеть при сравнении с использованием индукции BiP ER тапсигаргином (рис. 6). Во-вторых, многие из целей ER стресса, такие как кальретикулин и дисульфидные изомеразы, не индуцируются когда GRP94 теряется (Eletto соавт., В процессе подготовки). Кроме того, влияние стресса на ER IGF-II секрецию противоположно влиянию блокаторов GRP94. Как показано на рис. 3D, IGF-II секреция усиливается, не ингибируется, путем обработки низкой концентрацией туникамицина, либо в условиях роста или дифференцировки.
![]()
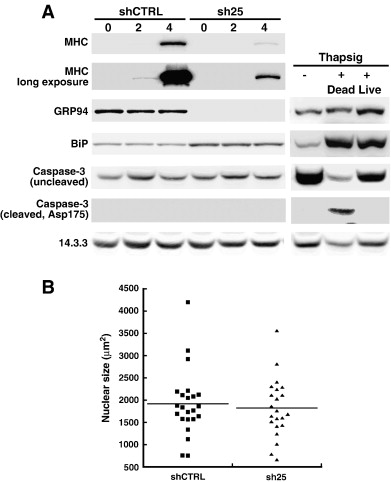
Известно, что во время миогенной дифференцировки в культуре фракции клеток умирают апоптозом, вызванным стрессом ER. Мы отделили дифференцирующиеся клетки от апоптоза с помощью 10T1/2MD: ER клеток, которые могут быть индуцированы к дифференцировке эстрадиолом даже в присутствии сыворотки. Когда вводили эстрадиол, эти клетки дифференцируются, как показано повышеной регуляцией экспрессии МНС (фиг. 6А). Без GRP94, дифференцировка в значительной степени подавляется. В этих условиях нет обнаруживается расщепление каспазы 3- отличительная черта апоптоза (рис. 6а). Если бы не было значительного апоптоза, это было бы легко обнаружить при активации каспазы 3, когда те же клетки обрабатываются индуктором ER стресса тапсигаргином (фиг. 6А). Кроме того, мы количественно определили размер ядра из DAPI-окрашенных изображений, таких как показаны на рис. 2, где C2C12 клетки дифференцируются после стресса. Там не было увеличения фракции клеток с конденсированными ядрами, SH25-экспрессирующих клеток по сравнению с контрольными клетками (фиг. 6В). Это согласуется с заключением, что истощение GRP94 не направляет клеточный апоптоз.
GRP94 важен для контроля клеточного цикла.
Миогенез включает в себя не только приверженность к дифференцировке и экспрессии мышц-специфических генов, но и запрограммированный выход из клеточного цикла, через понижающие регуляцию активаторы клеточного цикла и регуляцию ингибиторов клеточного цикла. Более того, в дополнение к ингибированию экспрессии специфических мышечных генов, отсутствие GRP94 замедляет клеточный цикл миогенных клеток. Это не уникальное свойство одной клеточной линии, потому что мы наблюдали пониженную пролиферацию MCF-7 RD, 293T и клеток СНО, а также первичных хондроцитов (данные с MCF-7 показано на рис. 7Б, представитель другие клетки ). Сайленсинг GRP94 привел к повышению экспрессии ингибиторов клеточного цикла p21 даже в среде роста, до индукции дифференцировки (фиг.5А). В соответствии с уровнями р21, экспрессия циклина D1 была ниже в C2C12, в которых экспрессия GRP94 была подавлена (данные не показаны). Эти данные хорошо согласуются с наблюдаемым уменьшением скорости пролиферации.
![]()
Обсуждение
Эта работа демонстрирует, что миогенная дифференцировка в культуре требует активности белка ER стресса GRP94 и обеспечивает молекулярное объяснение этому требованию. Мы покажем, насколько нужен GRP94 в первую очередь из-за его влияния на производство IGF, факторов роста, которые очень важны для развития мышц, а также для поддержания мышечной массы. Как мы недавно показали с использованием другой экспериментальной системы, что GRP94 действует как шаперон к ИФР-II, и благодаря его активности про-ИФР-II обрабатывается до зрелого, активного гормона. Давно известно, что дифференцировка миобластов в сократительной миотубы включает аутокринную секрецию IGF, как IGF-I и IGF-II, и мы показали здесь, что GRP94 необходим для этого существенного шага миогенеза. Далее мы показали потребность в GRP94 не для инициирования миогенеза, а при реализации программы дифференцировки после индукции MyoD и миогенина до увеличения. Этапы миогенеза действительно требовали GRP94 для IGF, как было показано ранее. IGF сигнализация интегрирует сигналы окружающей среды с собственной генетической программой, которая контролируется MyoD MEF2 и семейным факторами транскрипции, в создании сети, где аутокринные MyoD усиливают позже шаги по индукции местного действия фактора роста. С другой стороны, фактор роста усиливает влияние на экспрессию MyoD транскрипционных регуляторов совместно необходимых сопутствующих факторов для MyoD. Создание усиления каскада для поддержания ранних событий дифференцировки в мышцах. Хотя априори недостаточность миогенеза в отсутствие GRP94 может быть отнесена к нескольким белкам, производство которых может быть необходимо для терминальной дифференцировки, активность GRP94 к IGF играет основную роль в этом процессе, потому что экзогенная поставка гормонов дополняет слияние миобластов в терминальной стадии дифференцировки даже в клетках, лишенных GRP94. Поэтому, даже если другие белки зависят от GRP94 деятельности, их роль в дифференцировке не является существенной. Например, GRP94 необходим для правильной экспрессии некоторых поверхностных белков. Тем не менее, клетки с отсутствием GRP94 могут сливаться, когда получают только IGF. Таким образом, существует, по-видимому, достаточно компенсационная деятельность, предусмотренная GRP94 независимыми интегринами. Другая линия доказательств, указывающих на важнейшую роль GRP94 в IGF-оси является этапом миогенеза, которая требует GRP94 деятельности. В GRP94 обедненных миогенных клетках был нормальный уровень MyoD, но выражение миогенина, десмина, тяжелой цепи миозина, тропонина Т и саркомерного актина были незначительными. Действие IGF, как показано, является критическим для активации миогенных промоторов, синтеза белка, апоптоза, регенерации и гипертрофии мышц. Мышечные клетки без GRP94 не способны производить IGF, MyoD, и дифференцировка в клетках не в состоянии приступить к терминальной стадии. Завершение миогенной дифференцировки требует GRP94 IGF-оси. Наши данные являются дополнением к предыдущим наблюдениям Gorza соавт.. Повышенная экспрессия GRP94 служит в качестве защитного механизма против мышечного стресса, такие как кальциевые перегрузки в кардиомиоцитах. Мы предложили, что увеличение GRP94 выражения, необходимы для поощрения местного производства IGF напряженных миоцитов, что позволяет им лучше справляться со стрессом. Интересно, что Фрассон соавт. недавно сообщали, что в начале дифференциации миоцитов фракция GRP94 фосфорилируется, а затем экспортированы из эндоплазматического ретикулума в комплекс Гольджи. Таким образом, фосфорилирование GRP94 было зарегистрировано в кардиомиоцитах. Функциональная связь этого фосфорилированного пула GRP94 молекулы и IGF - зависимой дифференцировки мышцы в настоящее время неизвестна, и эксперименты в настоящее время продолжают изучать это интригующее отношение. Наши данные в добавление к опубликованной работе утверждают, что белок оказывает положительное влияние на миогенез. Например, Наканиши и соавт. показали, что предварительная обработка клеток С2С12 туникамицином или тапсигаргином на самом деле способствует дифференцировке клеток вступивших на путь апоптоза. Мы обнаружили, что низкие дозы туникамицина не только побуждают некоторые развернутые ответы белка, но так же способствуют IGF-II секреции, даже в условиях роста, не вызывая полной программы дифференцировки. Тем не менее, эффект от белка отличен от последствий истощения GRP94, даже если эти два процесса разделяют индукцию экспрессии BiP. Хотя мы ориентируемся на IGF в этой работе, наши
![]()
данные не исключают других видов деятельности или целей GRP94, которые действительно могут влиять на мышечную дифференцировку. Например, GRP94, как известно, связан с высокой производительностью кальций-связывающего белка, и эта деятельность необходима для кальциевого гомеостаза. Вполне вероятно, что отсутствие кальций-связывающей способности GRP94, ставит под угрозу дифференцировку саркоплазматического ретикулума. Более того, наши данные действительно показывают, что GRP94 деятельность так же важна для контроля клеточного цикла еще до начала дифференцировки, и выход из клеточного цикла, как известно, необходимы для миогенной дифференцировки. Хотя GRP94-дефицитные клетки имеют более высокий уровень p21 в условиях роста, они не могут выполнить программы дифференцировки. В настоящее время у нас нет молекулярного объяснения для пересечения с GRP94 регуляторами клеточного цикла. Дополнительные механизмы, следовательно, стоит исследовать; GRP94, как известно, важен для правильной экспрессии некоторых интегринов, и это может быть важным для слияния-компетенции миобластов. Доказательства приведенные в этой статье показывают, что GRP94 имеет важное значение для формирования мышц в первую очередь из-за его шаперонной активности необходима для производства IGF. Мы предлагаем не только IGF-II, но также, IGF-IA и другие связанные изоформы зависят от GRP94.
Ссылки
[1]S. Wanderling, B. B. Simen, O. Ostrovsky, N. T. Ahmed, S. Vogen, T. Gidalevitz, Y. Argon
GRP94 is essential for mesoderm induction and muscle development because it regulates IGF secretion
Mol. Biol. Cell, 18 (2007), pp. 3764–3775
View Record in Scopus | Full Text via CrossRef | Cited By in Scopus (47)
[2]L. Gorza, M. Vitadello
Reduced amount of the glucose-regulated protein GRP94 in skeletal myoblasts results in loss of fusion competence
FASEB J., 14 (2000), pp. 461–475
View Record in Scopus | Cited By in Scopus (30)
[3]M. Vitadello, P. Colpo, L. Gorza
Rabbit cardiac and skeletal myocytes differ in constitutive and inducible expression of the glucose-regulated protein GRP94
Biochem. J., 332 (1998), pp. 351–359
View Record in Scopus | Cited By in Scopus (14)
[4]M. Vitadello, J. Ausma, M. Borgers, A. Gambino, D. C. Casarotto, L. Gorza
Increased myocardial GRP94 amounts during sustained atrial fibrillation: a protective response?
Circulation, 103 (2001), pp. 2201–2206
View Record in Scopus | Full Text via CrossRef | Cited By in Scopus (18)
[5]M. Vitadello, D. Penzo, V. Petronilli, G. Michieli, S. Gomirato, R. Menabo, F. Di Lisa, L. Gorza
Overexpression of the stress protein Grp94 reduces cardiomyocyte necrosis due to calcium overload and simulated ischemia
FASEB J., 17 (2003), pp. 923–925
[6]O. Ostrovsky, N. T. Ahmed, Y. Argon
GRP94 protects cells grown without serum from apoptotis by regulating the secretion of insulin-like growth factor II
Mol. Biol. Cell, 20 (2009), pp. 1855–1864
[7]A. Musaro, K. McCullagh, A. Paul, L. Houghton, G. Dobrowolny, M. Molinaro, E. R. Barton, H. L. Sweeney, N. Rosenthal
Localized Igf-1 transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle
Nat. Genet., 27 (2001), pp. 195–200
[8]A. Philippou, M. Maridaki, A. Halapas, M. Koutsilieris
The role of the insulin-like growth factor 1 (IGF-1) in skeletal muscle physiology
In Vivo, 21 (2007), pp. 45–54
[9]G. Goldspink
Mechanical signals, IGF-I gene splicing, and muscle adaptation
Physiology (Bethesda), 20 (2005), pp. 232–238
[10]A. Musaro
Growth factor enhancement of muscle regeneration: a central role of IGF-1
Arch. Ital. Biol., 143 (2005), pp. 243–248
[11]C. Biswas, O. Ostrovsky, C. A. Makarewich, S. Wanderling, T. Gidalevitz, Y. Argon
The peptide binding activity of GRP94 is regulated by calcium
Biochem. J., 405 (2007), pp. 233–241
[12]K. Prelle, A. M. Wobus, O. Krebs, W. F. Blum, E. Wolf
Overexpression of insulin-like growth factor-II in mouse embryonic stem cells promotes myogenic differentiation
Biochem. Biophys. mun., 277 (2000), pp. 631–638
[13]S. Lee, E. R. Barton, H. L. Sweeney, R. P. Farrar
Viral expression of insulin-like growth factor-I enhances muscle hypertrophy in resistance-trained rats
J. Appl. Physiol., 96 (2004), pp. 1097–1104
[14]E. R. Barton, L. Morris, A. Musaro, N. Rosenthal, H. L. Sweeney
![]()
Muscle-specific expression of insulin-like growth factor I counters muscle decline in mdx mice
J. Cell. Biol., 157 (2002), pp. 137–148
[15]L. Pelosi, C. Giacinti, C. Nardis, G. Borsellino, E. Rizzuto, C. Nicoletti, F. Wannenes, L. Battistini, N. Rosenthal, M. Molinaro, A. Musaro
Local expression of IGF-1 accelerates muscle regeneration by rapidly modulating inflammatory cytokines and chemokines
Faseb J., 21 (2007), pp. 1393–1402
[16]E. R. Barton
The ABCs of IGF-I isoforms: impact on muscle hypertrophy and implications for repair
Appl. Physiol. Nutr. Metab., 31 (2006), pp. 791–797
[17]S. M. Vogen, T. Gidalevitz, C. Biswas, B. S. Simen, E. Stein, F. Gulmen, Y. Argon
Radicicol-sensitive peptide binding to the N-terminal portion of GRP94
J. Biol. Chem., 277 (2002), pp. 40742–40750
[18]S. M. Hollenberg, P. F. Cheng, H. Weintraub
Use of a conditional MyoD transcription factor in studies of MyoD trans-activation and muscle determination
Proc. Natl. Acad. Sci. U. S. A., 90 (1993), pp. 8028–8032
View Record in Scopus | Full Text via CrossRef | Cited By in Scopus (125)
[19]J. Baker, J. P. Liu, E. J. Robertson, A. Efstratiadis
Role of insulin-like growth factors in embryonic and postnatal growth
Cell, 75 (1993), pp. 73–82
Article | PDF (3817 K) | View Record in Scopus | Cited By in Scopus (1354)
[20]J. Tureckova, E. M. Wilson, J. L. Cappalonga, P. Rotwein
Insulin-like growth factor-mediated muscle differentiation: collaboration between phosphatidylinositol 3-kinase-Akt-signaling pathways and myogenin
J. Biol. Chem., 276 (2001), pp. 39264–39270
View Record in Scopus | Full Text via CrossRef | Cited By in Scopus (100)
[21]E. M. Wilson, M. M. Hsieh, P. Rotwein
Autocrine growth factor signaling by insulin-like growth factor-II mediates MyoD-stimulated myocyte maturation
J. Biol. Chem., 278 (2003), pp. 41109–41113
View Record in Scopus | Full Text via CrossRef | Cited By in Scopus (56)
[22]P. Klover, L. Hennighausen
Postnatal body growth is dependent on the transcription factors signal transducers and activators of transcription 5a/b in muscle: a role for autocrine/paracrine insulin-like growth factor I
Endocrinology, 148 (2007), pp. 1489–1497
View Record in Scopus | Cited By in Scopus (47)
[23]N. Morishima, K. Nakanishi, H. Takenouchi, T. Shibata, Y. Yasuhiko
An endoplasmic reticulum stress-specific caspase cascade in apoptosis. Cytochrome c-independent activation of caspase-9 by caspase-12
J. Biol. Chem., 277 (2002), pp. 34287–34294
View Record in Scopus | Full Text via CrossRef | Cited By in Scopus (440)
[24]K. Nakanishi, do, N. Morishima
Endoplasmic reticulum stress signaling transmitted by ATF6 mediates apoptosis during muscle development
J. Cell. Biol., 169 (2005), pp. 555–560
View Record in Scopus | Full Text via CrossRef | Cited By in Scopus (62)
[25]M. Kitzmann, A. Fernandez
Crosstalk between cell cycle regulators and the myogenic factor MyoD in skeletal myoblasts
Cell. Mol. Life Sci., 58 (2001), pp. 571–579
[26]A. C. Paul, N. Rosenthal
Different modes of hypertrophy in skeletal muscle fibers
J. Cell. Biol., 156 (2002), pp. 751–760
[27]A. Musaro, K. J. McCullagh, F. J. Naya, E. N. Olson, N. Rosenthal
IGF-1 induces skeletal myocyte hypertrophy through calcineurin in association with GATA-2 and NF-ATc1
Nature, 400 (1999), pp. 581–585
[28]D. D. Sarbassov, R. Stefanova, V. G. Grigoriev, C. A. Peterson
Role of insulin-like growth factors and myogenin in the altered program of proliferation and differentiation in the NFB4 mutant muscle cell line
Proc. Natl. Acad. Sci. U. S. A., 92 (1995), pp. 10874–10878
[29]E. M. Wilson, P. Rotwein
Control of MyoD function during initiation of muscle differentiation by an autocrine signaling pathway activated by
![]()
insulin-like growth factor-II
J. Biol. Chem., 281 (2006), pp. 29962–29971
[30]F. Randow, B. Seed
Endoplasmic reticulum chaperone gp96 is required for innate immunity but not cell viability
Nat. Cell Biol., 3 (2001), pp. 891–896
[31]U. Mayer
Integrins: redundant or important players in skeletal muscle?
J. Biol. Chem., 278 (2003), pp. 14587–14590
[32]J. Palsgaard, A. E. Brown, M. Jensen, R. Borup, M. Walker, P. De Meyts
Insulin-like growth factor I (IGF-I) is a more potent regulator of gene expression than insulin in primary human myoblasts and myotubes
Growth Horm. IGF Res., 19 (2009), pp. 168–178
[33]S. Noguchi
The biological function of insulin-like growth factor-I in myogenesis and its therapeutic effect on muscular dystrophy
Acta Myol., 24 (2005), pp. 115–118
[34]E. Tarricone, A. Ghirardello, S. Zampieri, R. M. Elisa, A. Doria, L. Gorza
Cell stress response in skeletal muscle myofibers
Ann. N. Y. Acad. Sci., 1069 (2006), pp. 472–476
[35]M. Frasson, M. Vitadello, A. M. Brunati, N. La Rocca, E. Tibaldi, L. A. Pinna, L. Gorza, A. Donella-Deana
Grp94 is Tyr-phosphorylated by Fyn in the lumen of the endoplasmic reticulum and translocates to Golgi in differentiating myoblasts
Biochim. Biophys. Acta, 1793 (2008), pp. 239–252
[36]S. E. Cala, L. R. Jones
GRP94 resides within cardiac sarcoplasmic reticulum vesicles and is phosphorylated by casein kinase II
J. Biol. Chem., 269 (1994), pp. 5926–5931
[37]K. Nakanishi, N. Dohmae, N. Morishima
Endoplasmic reticulum stress increases myofiber formation in vitro
FASEB J., 21 (2007), pp. 2994–3003
View Record in Scopus | Full Text via CrossRef | Cited By in Scopus (34)
[38]D. P.J. Macer, G. L.E. Koch
Identification of a set of calcium-binding protein in reticuloplasm, the luminal content of the endoplasmic reticulum
J. Cell Sci., 91 (1988), pp. 61–70
View Record in Scopus | Cited By in Scopus (74)
[39]Y. Fujio, K. Guo, T. Mano, Y. Mitsuuchi, J. R. Testa, K. Walsh
Cell cycle withdrawal promotes myogenic induction of Akt, a positive modulator of myocyte survival
Mol. Cell. Biol., 19 (1999), pp. 5073–5082
View Record in Scopus | Cited By in Scopus (144)
[40]Q. Wei, B. M. Paterson
Regulation of MyoD function in the dividing myoblast
FEBS Lett., 490 (2001), pp. 171–178
Переведено проектом МОЙМИО: http://mymio. org/
![]()
Оригинал статьи: http://www. /science?_ob=Artic
