
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

Антагонисты миостатина усиливают восстановление мышц во время саркопении
У млекопитающих масса скелетных мышц зависит от количества мышечных волокон, их типа и размера. Все три аспекта, как полагают, изменяются под влиянием различных условий, в том числе старения, которое может привести к атрофии отдельных мышечных волокон, потери волокна, и переключение между типами волокна. Важным компонентом размера мышечных волокон является участие клеток-сателлитов, популяции миогенных клеток-предшественников, связанных с мышечными волокнами. Спутниковое клетки расположены между базальной пластинки и сарколеммой и существуют преимущественно в фазу митотического покоя. Эти клетки, как полагают, в значительной степени ответственны за регенерацию мышц, порождая миобласты, тем самым позволяя дополнительным ядрам сливаются с существующими волокнами или формируя новые мышечные волокна. Для прогрессирования этого процесса сателлитным клеткам необходимо выйти из состояния покоя и начать миогенез. Очевидно, что число клеток-сателлитов повлияет на степень восстановления мышц. Одним из факторов, которые, как полагают, может серьезно повлиять на поведение спутниковых клеток является старение. Спутниковые клетки, выделенные у старых животных показали значительное отставание при входе в клеточный цикл, снижение пролиферации и потенциал а дифференцировки, и повышенную склонность к аппаптозу. Кроме того, различные исследования показывают, что с возрастом мышцы ограничены в своих способностях содействовать активации спутниковых клеток. Использование трансплантации поперечных мышц продемонстрировало,, что бедная регенерация, связанна с возрастом животных, и является функцией возрастных нарушений. Кроме того, есть предположения, что недостаточная регуляция No - сигнального пути несет прямую ответственность за нарушение активации спутниковых клеток с возрастом. Совсем недавно, эксперименты показали, что системные факторы, от молодых мышей восстановливали активация сигнального пути, а также способствовали распространению и регенеративной способности сателлитных клеток. Кроме того, негативные регуляторы, такие как миостатин, могут подавлять активность спутниковых клеток. Миостатин является трансформирующим фактором роста, и действует как мощный ингибитор роста мышцы. Это выражается как в пре-, так натально развивающихся миотомах и после рождения во взрослых скелетных мышцах, тем самым предполагая, что миостатин играет постоянную роль в миогенезе. Действительно, у животных с отсутствием миостатина наблюдается значительно больше мышечной массы в результате гиперплазии мышц и их гипертрофии. Исследования показывают, что миостатин влияет на миогенез C2C12 культур миобластов через регуляции клеточного цикла и миогенных факторов. Кроме того, миостатин ингибирует активацию спутниковых клеток у мышей, и это будет иметь серьезные последствия в контексте возрастной атрофии мышц и регенерации. Интересно, что увеличение миостатина было связано с атрофией мышц во время разгрузки у мышей, атрофии мышц при инфекции вирусом иммунодефицита человека, и кахексии при индуцированном системном введении миостатина Хотя точная роль миостатина в возрастной регуляции мышечной массы остается неясной, его длительное отсутствие в дородовом этапе, как показано на мышах, продемонстрировало саркопению. Скорей всего, антагонист миостатина будет иметь значительный терапевтический потенциал при саркопении и нарушении регенерации мышц, которое наблюдается у старых животных. В самом деле, по нашим данным, краткосрочная блокада функции миостатина с помощью введения усеченного белока значительно повышает регенерацию мышц у пожилых мышей. Эта регенерация произошла в процессе восстановления миогенной и воспалительной реакции у старых мышей, что привело к увеличению спутниковой активности клеток и макрофагов и расширения миграции миобластов.
Результаты
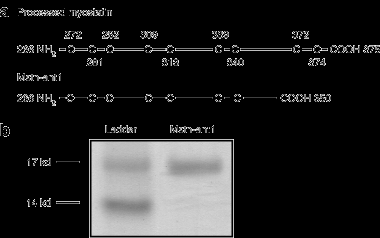
Производство антагониста MSTN-ANT1
Ранее мы показали, что длительное отсутствие миостатина уменьшает саркопения у миостатина-нулевых mice. Мы избрали для проверки конкретные молекулы, усеченную версию миостатина, которая может подавлять синтез миостатина и, возможно, имеет терапевтические возможности для лечения саркопении. Миостатин протеолитически перерабатывается, что приводит к биологически активному зрелому миостатину, который охватывает аминокислоты 266-375. Этот 110-аминокислотный белок связывается с рецептором активин типа IIB в виде димера. Для того, чтобы противодействовать миостатину, мы подготовили усеченный с С конца на аминокислоты с 266 до 350, в качестве доминирующего отрицательного
![]()
(миметического) белка. Это усечение назвали MSTN-ANT1,, показан на рисунке 1а. MSTN-ANT1 был экспрессирован кишечной палочкой и очищен до гомогенности (рис. 1б).
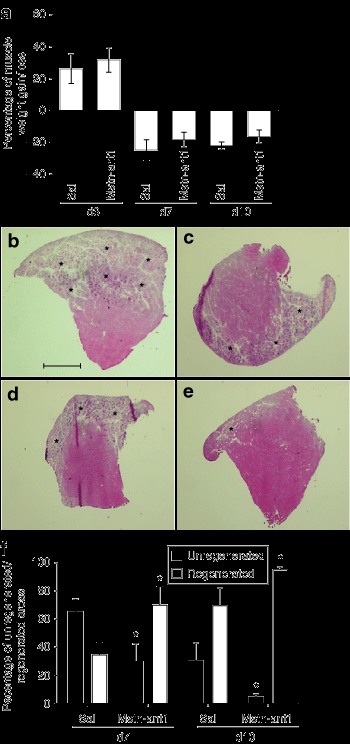
Антагонист миостатина усиливает регенерацию мышц после травмы.
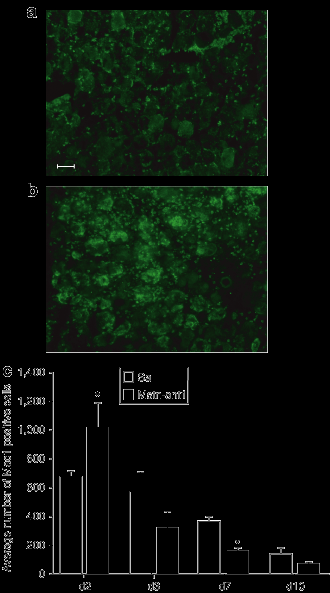
Далее мы проверили пользу MSTN-ANT1 в модели у мышей. Обычно после травмы, есть изначальное увеличение веса раненых мышц вследствие отека, а затем снижение вызвано некрозом поврежденных мышечных волокон. Мышечная масса затем начинает возрастать снова, как происходит регенерация волокон. Полученные результаты свидетельствуют о тенденции, в которой потери мышечной массы была менее выраженной у MSTN-ANT1 обработанных мышц по сравнению с обработанными физиологическим раствором мышцами в дни 7 и 10 (рис. 2а).Наблюдаемая тенденция может быть связана с сокращением потери мышечной массы во время некроза или, наоборот, в связи с формированием новых волокон в результате антагонизма MSTN-ANT1 к миостатину. В подтверждение этого, анализ передней большеберцовой мышцы (TA) показал, что зарождающееся формирование мышечных волокон (районы регенерации) проходит раньше, и не было связанно с сокращением некротической области (неперерожденной области) в мышцах обработанных MSTN - ANT1 по сравнению с обработанными физиологическим раствором мышцами в дни 7 и 10 (рис. 2, б-е) (P <0,05). Измерение волокна может указывать на прогрессирование регенерации, происходящее в восстановленных мышцах. Поэтому мы были намерены оценить регенерацию волокна через 28 дней после травмы, и результаты показали, что регенерированных волокон из мышц MSTN-ANT1-мышей было значительно больше (рис. 3) (P <0,05). Кроме того, поскольку уровни коллагенных депозитов могут быть измерены в регенерирующих после травмы мышцах, окрашивание по Ван Гизону было проведено (рис. 3а, б). Снижение уровня коллагена было найдено в MSTN-ANT1 обработанных мышцах в дни 10 и 28 (P <0,05) (рис. 3).
![]()
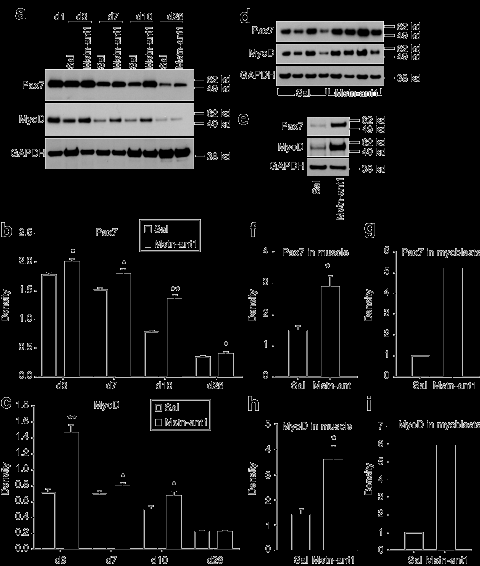
Экспрессия MyoD и Pax7 в регенерирующих мышцах изменяется при лечении антагонистом. Было установлено, что Pax7 и MyoD являются сильными маркерами миогенеза. В то время как уровень Pax7 может означать степень активации бассейна спутниковых клеток, а также их самообновление, MyoD может означать уровень миогенеза, происходящего в мышцах. Для исследования Pax7 и MyoD уровня белка в поврежденных мышцах, Вестерн-блот анализ был выполнен. Анализ контроля и MSTN-ANT1 обработанных мышц показал, что уровень Pax7 белка был выше у получавших MSTN-ANT1 (рис. 4, б) (P <0,05). Точно так же, MyoD уровни были выше у получавших MSTN-ANT1 лечение, чем получавшие физиологический раствор (рис. 4а и C) (P <0,05). Сопоставимые уровни MyoD были отмечены между группами на 28 день, однако, более высокий уровень Pax7 наблюдается в той же точке времени с группой, получавшей MSTN-ANT1. Как и в случае повреждения нотексином, хотя и в меньшей степени, с возрастом мышцы также отображают уровень текущего миогенеза за счет поддержания и восстановления мышц, связанного с процессом старения. Таким образом, Pax7 и MyoD также проанализированы в мышцах мышей, получавших в течение 6 недель инъекции с физиологическим раствором или MSTN-ANT1. Уровни белка, выделенного из икроножной мышцы при завершении исследования, значительно выше Pax7 и MyoD белков в MSTN-ANT1-обработанных мышцах мышей по сравнению с контрольной группой мышей (рис. 4d, Р и Н) (Р <0,05). Кроме того, белок, выделенный из активно растущих первичных миобластов, также отображается повышение уровня и Pax7 и MyoD (рис. 4e, г и я) после MSTN-ANT1 лечения.
![]()
Спутниковая активация и пролиферация клеток и увеличение мышечной силы в ответ на MSTN-ANT1 лечение.
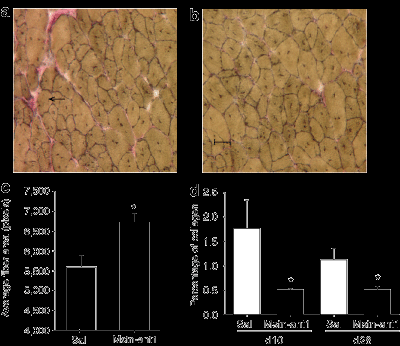
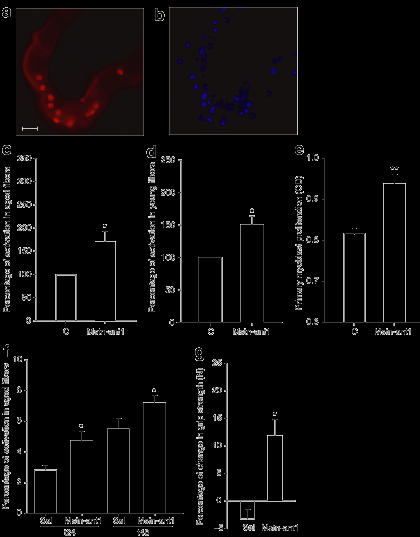
Чтобы исследовать, как MSTN-ANT1 повлияет на спутниковую активацию клеток, изолированные одиночные волокна 1 - и 24-месячных мышей культивировали с или без MSTN-ANT1. Спутниковая активация клеток определяется путем иммуноцитохимии с использованием антител для ядерного антигена пролиферирующих клеток, маркера репликации ДНК (рис. 5а). Волокна, культивированные с MSTN-ANT1, последовательно демонстрируют высокий процент активированных клеток-сателлитов на волокно, чем волокна, культивированных с физиологическим раствором (рис. 5в, г) (P <0,05).
Миостатин влияет на пролиферацию клеток.
Таким образом, для изучения эффективности антагонистов в увеличении распространения миобластов у 1-месячных мышей культивировали с или без MSTN-ANT1. Результаты показали, что культивирование с MSTN-ANT1 увеличило распространение миобластов на 15% (рис. 5е) (P <0,001).В дополнение к культивированию изолированных волокон с MSTN-ANT1, спутниковая активация клеток была также исследована в волокнах изолированных от мышей, которым вводили физиологический раствор или MSTN-
![]()
ANT1 в течение 6 недель. Волокна изолированые от MSTN-ANT1 мышей отображают значительно более высокий процент активированных спутниковых клеток в волокнах (рис. 5е) (P <0,05). Кроме того, чтобы проверить, может ли 6-недельное лечение увеличить мышечную силу, испытания силы мышц были проведены в начале и завершении периода лечения. Результаты показали, что MSTN-ANT1 значительно увеличило силу старых мышей на 12% (P <0,05) (рис. 5г).Антагонист изменяет миграцию макрофагов, и, как полагают, препятствует миграции макрофагов. Поэтому в поврежденные мышцы вводили физиологический раствор или MSTN-ANT1 и обследовали на наличие макрофагов, анти-Mac1 антитела были использованы для определения эффективности антагониста в усилении миграции макрофагов. Действительно, на 2-й день после травмы, мышцы, которые были обработаны MSTN-ANT1, показали больший процент Mac1-положительных ядер (рис. 6а-с) (P <0,05). На 3-й день, этот процент упал в MSTN-ANT1 обработанных мышцах ниже, чем в мышцах, в которые вводили физиологический раствор, и продолжал быть ниже в дни 7 и 10.
Миобласты и миграция макрофагов изменяются с возрастом и при MSTN-ANT1 лечении.
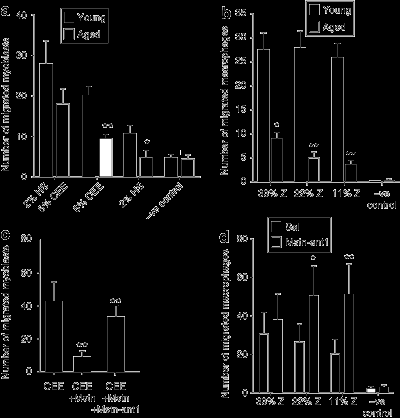
Миобласты и миграция макрофагов, как известно, изменяется под влиянием сигналов хемотаксиса. Для того чтобы выяснить влияние последствий воздействия миостатина и старения на миграцию миобластов и макрофагов, был выполнен ряд анализов хемотаксиса. Полученные результаты свидетельствуют, что миграция значительно отстает в изолированных от 24-месячных мышей по сравнению с 1-месячными мышами в присутствии 5% экстракта куриного эмбриона или 2% лошадиной сыворотки (рис. 7а) (P <0,05). Точно так же, миграция макрофагов изолированных от 24-месячныъ мышей была ниже по сравнению с 1-месячными мышами, независимо от концентрации хемоаттрактанта (рис. 7б) (P <0,001).Эффективность MSTN-ANT1 в восстановлении миграции клеток в ответ лечение исследовали с помощью первичных миобластов, изолированных от 24-месячных мышей. Когда MSTN-ANT1 был добавлен, можно было
![]()
восстановить миграцию клеток, препятствуя тормозящему действию миостатина (рис. 7) (P <0,05). Для того, чтобы проверить эффективность MSTN-ANT1 в усилении миграции клеток после длительного лечения в естественных условиях костного мозга, макрофаги были выделены у мышей, которым вводили в течение 6 недель физиологический раствор или MSTN-ANT1. На самом деле макрофагов MSTN-ANT1-обработанных мышей были способны мигрировать более эффективно (рис. 7б) (P <0,05).
Дискуссия
Многочисленные исследования показывают, что старение значительно влияет на миогенез из-за окружающей среды мышц или системных факторов. Недавно было показано, что длительное отсутствие миостатина снижает саркопению у миостатин-нулевых мышей. Однако, так как этим мышам не хватает миостатина в дородовой стадии, необходимо было провести дополнительные исследования, чтобы определить влияние краткосрочного ингибирования миостатина в пожилом возрасте. Предыдущее исследование показало, что увеличение мышечной массы и силы происходит после лечения ингибиторами миостатина. В этом исследовании мы показали, что краткосрочное введение антагониста миостатина восстанавливает регенеративную и миогенную мощность мышц. В соответствии с прошлыми исследованиями, которые показали, что отсутствие миостатина значительно улучшает регенерацию мышц и уменьшает фиброз, результаты, представленные здесь, предполагают, что блокада миостатина путем введения MSTN-ANT1 сразу после травмы может повторить этот же эффект. Вес мышц, гистология, большие регенерирующие области в дни 7 и 10, а также значительно больше отдельных участков волокна на 28-й день, все критерии регенерации наблюдались после MSTN-ANT1 лечения (рис. 2 и 3, в). Кроме того, MSTN-ANT1 обработанные мышцы отображают снижение уровня коллагена (рис. 3), тем самым предполагая, что миостатин участвует в процессах фиброза и образовании коллагена. Это подтверждается тем фактом, что во время миграции фибробластов миостатин может вести себя как хемоаттрактант фибробластов (данные не показаны). В совокупности, эти результаты показывают, что длительное отсутствие миостатина, как это наблюдалось у миостатин-нулевых мышей, не является необходимым для получения расширенной регенерации мышц и снижения фиброза. Скорее всего, краткосрочная блокада миостатина во время периода регенерации является достаточной для улучшения процесса регенерации. Во время восстановления мышц, MyoD выражается раньше и на более высоких уровнях в миостатина-нулевых мышцах по сравнению с диким типом мышц. Аналогичным образом, Вестерн-блот анализ регенерирующих мышц от мышей, получавших MSTN-ANT1, показал повышение уровней MyoD во время регенерации, тем самым предполагая, что увеличение миогенеза является прямым результатом блокады миостатина MSTN-ANT1 (рис. 4в). Кроме того, Pax7, который экспрессируется в спокойных и пролиферирующих клетках, был выше при MSTN-ANT1 лечении в течении испытательного периода, тем самым предлагая сравнительное увеличение числа спутниковых клетки, активации и / или самообновления (рис. 4б ). Эти более высокие Pax7 и MyoD уровни могут быть связаны с увеличением числа клеток-сателлитов и последующим миогенезом и увеличением спутникового обновления клеток. Следует отметить, что в то время как аналогичные уровни MyoD наблюдались в MSTN-
![]()
ANT1регенерирующих мышцах на 28-й день, более высокий уровень Pax7 были замечены в MSTN-ANT1 обработанных мышцах, чем в группе контроля. Pax7 является маркером для самообновления спутниковых клеток, высокий уровень Pax7 указывает, что MSTN-ANT1 должен повышать это самообновление. Это в соответствии с ранее сделанными выводами, означает, что миостатин тормозит спутниковые клетки самостоятельно. Кроме того, MSTN-ANT 1введение мышам в возрасте 6 недель также привело к увеличению MyoD и Pax7 белка в икроножных мышцах и первичных миобластов (рис. 4f-я). Возраст мышц может повлиять на этот процесс из-за потери волокон и атрофии существующих волокон, следовательно, продолжение миогенеза на таком же уровне необходим для поддержания и восстановления мышц. Соответственно, MyoD экспрессия, как было показано, высокая у молодых животных, значительно уменьшается у взрослого, а затем снова растет с возрастом. Однако, несмотря на уровни MyoD и других миогенных регуляторных факторов, увеличение возраста мышц не восстанавливает миогенную способность мышц к уровню молодых мышц. Потому что миостатин, как известно, уменьшает экспрессию MyoD, можно было бы ожидать, что миостатин негативно регулирует этот возрастной миогенез. В самом деле, путем противодействия миостатину, мы смогли повысить уровень миогенеза, о чем свидетельствует увеличение уровня как MyoD и Pax7 белка. В поддержку этих результатов, повышение спутниковой активации клеток наблюдалось в 1 - и 24-месячных одиночных волокнах при культивировании в присутствии MSTN-ANT1 (рис. 5в, г). Кроме того, MSTN-ANT1 вводили в естественных условиях мышам в возрасте, он был в состоянии увеличить процент активированных клеток-сателлитов на волокно (рис. 5е). Это предполагает, что антагонизм миостатина через подкожные инъекции MSTN-ANT1 может предоставить средства для увеличения миогенеза, возможно, за счет увеличения числа спутниковых клеток. Если это действительно так, то, возможно, антагонизм миостатина у старых животных может способствовать преодолению тенденции к сокращению сателлитных клеток и / или их потенциалу активации, который, как сообщается, уменьшается с возрастом. Увеличение активации спутниковых клеток, как можно было бы ожидать, приведет к увеличению прочности. Как показано на рисунке 5 , после 6-недельного лечения MSTN-ANT1, сила мышц увеличилась у обработанных животных (P <0,05).Как отмечалось ранее, одним из основных компонентов процесса регенерации является воспалительная реакция. После травмы, макрофаги, а также миобласты мигрируют в место повреждения в ответ на сигналы воспалительных цитокинов и различных факторов роста. Подтверждающие результаты исследования с использованием миостатин-нулевых мышей, ускоренная миграция макрофагов наблюдалось после MSTN-ANT1 лечения (день 2, рис 6в), тем самым предполагая, что MSTN-ANT1 быстро и эффективно противодействует тормозящему действию миостатина. Аналогичные выводы показали анализы хемоатрактантов (рис. 7). Старение значительно негативно влияло как на первичные миобласты и миграцию макрофагов (рис. 7а, б). Это может быть обусловлено снижением склонности к хемотаксису 24-месячных миобластов и макрофагов в ответ на их соответствующие хемоаттрактанты, или за счет снижением экспрессии рецепторов. Интересно, что после 6 недель MSTN-ANT1 лечения, миграционная способность макрофагов была восстановлена (в неизвестном степени) у старых мышей (рис. 7б). В совокупности представленные здесь результаты показывают, что краткосрочные блокады миостатина и его функции через лечение антагонистом может эффективно увеличить восстановление мышц у пожилых мышей после травм и во время возрастной атрофии мышц. Последствия лечения антагонистом для здоровья человека являются обширными. Эффективность лечения антагонистом была продемонстрирована здесь, только четыре дозы антагониста в критический период восстановления после травмы было достаточно, чтобы значительно улучшить восстановление мышц. Поэтому мы полагаем, что антагонист миостатина является жизнеспособным вариантом лечения недостаточной регенерации мышц и саркопении у людей, восстанавливая миогенную и воспалительную реакцию и уменьшая фиброз.
Материалы и методы
Генерация MSTN-ANT1.
Система кишечной палочки была использована для создания усеченного миостатина. Усиленную комплементарную ДНК из C-концевой области миостатина очищают, а затем вставляют в вектор рЕТ 16-B (Novagen, Madison, WI). В целях формирования MSTN-ANT1, комплементарная ДНК была усечена на 350 аминокислоты. Белок очищают с использованием никель-нитрилоуксусной кислоты (Ni-NTA) агарозы (Qiagen, Hilden, Германия) . Для того, чтобы проверить чистоту антагониста, он был отделен от NuPAGE в 4-12% Bis-Tris геле (Invitrogen, Carlsbad, CA), окрашивали кумасси синим, а затем обесцвечивали.
Животные.
Дикий тип мышей C57BL/10 был выведен на небольшой колонии животных. Все животные были обработаны в соответствии с руководящими принципами Ruakura животного комитета по этике (AgResearch, Гамильтон, Новая Зеландия).
Введение MSTN-ANT1 и нотексина мышам.
В 0 день 1-летних мышей анестезировали с 10% кетамином гидрохлоридом (100 мг / мл) / 5% Rompun (20 мг /
![]()
мл) в 0,1 мл / 7 г веса тела. Небольшой разрез был сделан на левой TA, и нотексин (10 г / мл; Venom, Австралия) вводили в мышцы. На 1, 3, 5 и 7 группы получили лечение MSTN-ANT1 на 6 г / г веса тела подкожно, в то время как контрольная группа получала эквивалентный объем физиологического раствора. Мыши были подвергнуты эвтаназии в дни 1, 2, 3, 7, 10 и 28 (4-6 мышей в группе в день). Кроме того, мышам в возрасте 13-16 месяцев вводили подкожно три раза в неделю MSTN-ANT1 на 6 г / г веса тела, или эквивалентный объем физиологического раствора, в течение 6 недель (10 в группе). ТА и икроножные мышцы были собраны для одного выделения волокон и мышечных белков, соответственно, в то время как остальные мышцы задних конечностей были собраны для изоляции миобластов. Костный мозг был также собран для культур макрофагов, используемых для анализа хемотаксиса.
Оценка регенерации мышечной массы и силы.
TA мышцы были вскрыты, взвешанны и заморожены для выделения белка или срезов ткани. Срезы мышц окрашивали гематоксилином и эозином для визуализации и измерения неперерожденной и регенерирующей области. Для визуализации областей коллагена на 10 и 28 день после травмы, мышечные срезы окрашивали по Ван Гизону.
Количество коллагена.
Затем измеряют в пределах каждого среза в процентах от общей площади сечения. Срезы мышц в дни 2, 3, 7, и 10 были исследованы на Mac1. Двойное измерение мышечной силы проводится у мышей, которым вводили физиологический раствор или MSTN-ANT1, в течение 6 недель в начале исследования и при завершении (MK-380S, Муромати, Токио, Япония).
Изоляция волокна и анализ активации спутниковых клеток.
TA мышцы собраны у мышей были использованы для выделения отдельных волокон. Изолированные волокна затем культивировали в течение 24 или 48 часов при 37 ° C в 5% CO2. Кроме того, отдельные волокна были выделены из необработанных 1-месячных и 24-месячных мышей. Эти волокна культивировали с или без MSTN-ANT1 с 5 г / мл в течение 24, 48 или 72 часов при 37 ° С в 5% СО2. После того, как требуемое время культивирования прошло, волокна подверглись иммуноокрашиванию на ядерный антиген пролиферирующих клеток и с 4 ',6-диамидино-2-фенилиндол (см. дополнительные данные S1).Положительные ядра на ядерный антиген пролиферирующих клеток были подсчитаны в процентах от общего числа ядер для нормализации данных и исключить любые расхождения в длине волокна. Выделение первичных миобластов и перитонеальных макрофагов. Спутниковое клетки были изолированы из мышц задних конечностей мышцы в соответствии с протоколом. После 48 часов культивирования клетки были собраны для изоляции белка. Кроме того, первичные миобласты, выделенные у необработанных 1-месячных и 24-месячных мышей для анализа хемотаксиса. Для анализа пролиферации миобласты культивировали с или без MSTN-ANT1 10 г / мл в течение 96 часов. Пролиферация клеток была позже оценена с помощью фотометрического анализа. Макрофаги костного мозга были получены путем посева клеток костного мозга на 5 10 6 клеток/пластина в среде Игла, модифицированной (DMEM) + 10% эмбриональной телячьей сывороткой + 10% L929 кондиционированной среде (содержащей колониестимулирующий фактор-1) в течение 5 дней, чтобы побудить дифференциацию макрофагов. Перитонеальные макрофаги были получены путем промывания брюшной полости 1-месячных и 24-месячных необработанных мышей холодным фосфатно-солевом буфером.
Миобласты и анализ макрофагального хемотаксиса.
Анализы хемотаксиса проводили в двух экземплярах с использованием культуры клеток, содержащие полиэтилентерефталат 0,8 м (BD Biosciences, San Jose, CA) Для хемотаксиса макрофагов, DMEM + зимозан (Sigma, Сент-Луис, Миссури), активированная мышинная сыворотка используется в качестве хемоаттрактанта на 33, 22 и 11% (об / об). Для хемотаксиса миобластов, DMEM + 2% лошадиной сыворотки + 5% экстракт куриных эмбрионов (оптимальной), DMEM + 2% лошадиной сыворотки (субоптимальных) или DMEM + 5% экстракт куриных эмбрионов (субоптимальных) были использованы в качестве хемоаттрактантов. Эксперименты проводились с 2,5 или 5 г / мл рекомбинантного миостатина в присутствии или в отсутствии MSTN-ANT1 . Клетки инкубировали в течение 4 и 7 часов для макрофагов и миобластов, соответственно. Перенесенные клетки подсчитывали по четыре поля на мембране.
Анализ белков.
TA мышцы мышей гомогенизировали в 1 мл буфера для лизиса белков [0,05 М Трис, рН 7,5, 0,25 М NaCl, 5 мМ ЭДТА, ингибитора протеазы (Roche, Annheim, Германия) и 0,1% NP40]. Реагента Брэдфорда (Bio-Rad Laboratories, Hercules, CA) был использован для оценки общего белка. Общий белок (10 г) был разделен на NuPAGE 4-12% Bis-Tris геле (Invitrogen) и перен на нитроцеллюлозные мембраны (Bio-Rad), прежде чем определены MyoD или Pax7 (см. дополнительные данные S1). Общий белок (15 г) был использован для вестерн блот анализа на MyoD и Pax7.
Изображение и статистический анализ.
Неперерожденные / регенерирующие области и отложение коллагена были проанализированы с помощью Olympus SZ-PT стереомикроскопа (Olympus, Токио, Япония), три прибора со связью с камерой (Dage MTI, Мичиган-Сити, IN) и Scion Image программного обеспечения (Scion, MD). Полученные изображения были затем оценены с помощью программного обеспечения ImageJ (Национальный институт здоровья). Положительное окрашивание на Mac1 оценено под флуоресцентным освещением с использованием микроскопа Olympus BX50, SPOT-RT 4,01 камеры и программное обеспечение (Diagnostic Instruments Inc, Sterling Heights, MI) Все данные представлены в виде значений и стандартных ошибок. Дисперсионный
![]()
анализ и тест Тьюки был использован для определения значимости (P <0,05), данные различия между группами.
Ссылки
REFERENCES
Alnaqeeb, MA and Goldspink, G (1987). Changes in fibre type, number and diameter in developing and ageing skeletal muscle. J Anat 153: 31–45. | PubMed | ISI | ChemPort |
Gutmann, E and Hanzlikova, V (1966). Motor unit in old age. Nature 209: 921–922. | Article | PubMed | ISI | ChemPort |
Holloszy, JO, Chen, M, Cartee, GD and Young, JC (1991). Skeletal muscle atrophy in old rats: differential changes in the three fiber types. Mech Ageing Dev 60: 199–213. | Article | PubMed | ISI | ChemPort |
Lexell, J, Taylor, CC and Sjostrom, M (1988). What is the cause of the ageing atrophy? Total number, size and proportion of different fiber types studied in whole vastus lateralis muscle from 15- to 83-year-old men. J Neurol Sci 84: 275–294. | Article | PubMed | ISI | ChemPort |
Rowe, RW (1969). The effect of senility on skeletal muscles in the mouse. Exp Gerontol 4: 119–126. | Article | PubMed | ISI | ChemPort |
Mauro, A (1961). Satellite cell of skeletal fibers. J Biophys Biochem Cytol 9: 493–498. | Article | PubMed | ISI | ChemPort |
Schultz, E, Gibson, MC and Champion, T (1978). Satellite cells are mitotically quiescent in mature mouse muscle: an EM and radioautographic study. J Exp Zool 206: 451–456. | Article | PubMed | ISI | ChemPort |
Partridge, TA (2002). Cells that participate in regeneration of skeletal muscle. Gene Ther 9: 752–753. | Article | PubMed | ISI | ChemPort |
Bischoff, R (1994). The satellite cell and muscle regeneration. In: Engel, AG and Franzini-Armstrong, C(eds). Myology: Basic and Clinical, McGraw-Hill, New York: pp. 97–112.
Schultz, E and McCormick, KM (1994). Skeletal muscle satellite cells. Rev Physiol Biochem Pharmacol 123: 213–257. | PubMed | ISI | ChemPort |
Johnson, SE and Allen, RE (1995). Activation of skeletal muscle satellite cells and the role of fibroblast growth factor receptors. Exp Cell Res 219: 449–453. | Article | PubMed | ISI | ChemPort |
Lees, SJ, Rathbone, CR and Booth, FW (2006). Age-associated decrease in muscle precursor cell differentiation. Am J Physiol Cell Physiol 290: C609–C615. | PubMed | ISI | ChemPort |
Schultz, E and Lipton, BH (1982). Skeletal muscle satellite cells: changes in proliferation potential as a function of age. Mech Ageing Dev 20: 377–383. | Article | PubMed | ISI | ChemPort |
Jejurikar, SS, Henkelman, EA, Cederna, PS, Marcelo, CL, Urbanchek, MG and Kuzon, WM Jr. (2006). Aging increases the susceptibility of skeletal muscle derived satellite cells to apoptosis. Exp Gerontol 41: 828–836. | Article | PubMed | ISI | ChemPort |
Krajnak, K, Waugh, S, Miller, R, Baker, B, Geronilla, K, Alway, SE et al. (2006). Proapoptotic factor Bax is increased in satellite cells in the tibialis anterior muscles of old rats. Muscle Nerve 34: 720–730. | Article | PubMed | ISI | ChemPort |
![]()
Bockhold, KJ, Rosenblatt, JD and Partridge, TA (1998). Aging normal and dystrophic mouse muscle: analysis of myogenicity in cultures of living single fibers. Muscle Nerve 21: 173–183. | Article | PubMed | ISI | ChemPort |
Conboy, IM, Conboy, MJ, Smythe, GM and Rando, TA (2003). Notch-mediated restoration of regenerative potential to aged muscle. Science 302: 1575–1577. | Article | PubMed | ISI | ChemPort |
Decary, S, Mouly, V, Hamida, CB, Sautet, A, Barbet, JP and Butler-Browne, GS (1997). Replicative potential and telomere length in human skeletal muscle: implications for satellite cell-mediated gene therapy. Hum Gene Ther 8: 1429–1438. | PubMed | ISI | ChemPort |
Carlson, BM and Faulkner, JA (1989). Muscle transplantation between young and old rats: age of host determines recovery. Am J Physiol 256: C1262–C1266. | PubMed | ISI | ChemPort |
Conboy, IM, Conboy, MJ, Wagers, AJ, Girma, ER, Weissman, IL and Rando, TA (2005). Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature 433: 760–764. | Article | PubMed | ISI | ChemPort |
McPherron, AC, Lawler, AM and Lee, SJ (1997). Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 387: 83–90. | Article | PubMed | ISI | ChemPort |
Langley, B, Thomas, M, Bishop, A, Sharma, M, Gilmour, S and Kambadur, R (2002). Myostatin inhibits myoblast differentiation by down-regulating MyoD expression. J Biol Chem 277: 49831–49840. | Article | PubMed | ISI | ChemPort |
Thomas, M, Langley, B, Berry, C, Sharma, M, Kirk, S, Bass, J, et al (2000). Myostatin, a negative regulator of muscle growth, functions by inhibiting myoblast proliferation. J Biol Chem 275: 40235–40243. | Article | PubMed | ISI | ChemPort |
McCroskery, S, Thomas, M, Maxwell, L, Sharma, M and Kambadur, R (2003). Myostatin negatively regulates satellite cell activation and self-renewal. J Cell Biol 162: 1135–1147. | Article | PubMed | ISI | ChemPort |
Carlson, CJ, Booth, FW and Gordon, SE (1999). Skeletal muscle myostatin mRNA expression is fiber-type specific and increases during hindlimb unloading. Am J Physiol 277: R601–R606. | PubMed | ISI | ChemPort |
Gonzalez-Cadavid, NF, Taylor, WE, Yarasheski, K, Sinha-Hikim, I, Ma, K, Ezzat, S et al. (1998). Organization of the human myostatin gene and expression in healthy men and HIV-infected men with muscle wasting. Proc Natl Acad Sci USA 95: 14938–14943. | Article | PubMed | ChemPort |
Zimmers, TA, Davies, MV, Koniaris, LG, Haynes, P, Esquela, AF, Tomkinson, KN et al. (2002). Induction of cachexia in mice by systemically administered myostatin. Science 296: 1486–1488. | Article | PubMed | ISI | ChemPort |
Siriett, V, Platt, L, Salerno, MS, Ling, N, Kambadur, R and Sharma, M (2006). Prolonged absence of myostatin reduces sarcopenia. J Cell Physiol 209: 866–873. | Article | PubMed | ISI | ChemPort |
Wagner, KR, Liu, X, Chang, X and Allen, RE (2005). Muscle regeneration in the prolonged absence of myostatin. Proc Natl Acad Sci USA 102: 2519–2524. | Article | PubMed | ChemPort |
Lee, SJ and McPherron, AC (2001). Regulation of myostatin activity and muscle growth. Proc Natl Acad Sci USA 98: 9306–9311. | Article | PubMed | ChemPort |
McCroskery, S, Thomas, M, Platt, L, Hennebry, A, Nishimura, T, McLeay, L et al. (2005). Improved muscle healing through enhanced regeneration and reduced fibrosis in myostatin-null mice. J Cell Sci 118: 3531–3541. | Article | PubMed | ISI | ChemPort |
![]()
Oustanina, S, Hause, G and Braun, T (2004). Pax7 directs postnatal renewal and propagation of myogenic satellite cells but not their specification. EMBO J 23: 3430–3439. | Article | PubMed | ISI | ChemPort |
Seale, P, Sabourin, LA, Girgis-Gabardo, A, Mansouri, A, Gruss, P and Rudnicki, MA (2000). Pax7 is required for the specification of myogenic satellite cells. Cell 102: 777–786. | Article | PubMed | ISI | ChemPort |
Grounds, MD, Garrett, KL, Lai, MC, Wright, WE and Beilharz, MW (1992). Identification of skeletal muscle precursor cells in vivo by use of MyoD1 and myogenin probes. Cell Tissue Res 267: 99–104. | Article | PubMed | ISI | ChemPort |
Bischoff, R (1997). Chemotaxis of skeletal muscle satellite cells. Dev Dyn 208: 505–515. | Article | PubMed | ISI | ChemPort |
Jones, GE (2000). Cellular signaling in macrophage migration and chemotaxis. J Leukoc Biol 68: 593–602. | PubMed | ISI | ChemPort |
Welle, S, Bhatt, K and Thornton, CA (2000). High-abundance mRNAs in human muscle: comparison between young and old. J Appl Physiol 89: 297–304. | PubMed | ISI | ChemPort |
Whittemore, LA, Song, K, Li, X, Aghajanian, J, Davies, M, Girgenrath, S et al. (2003). Inhibition of myostatin in adult mice increases skeletal muscle mass and strength. Biochem Biophys Res Commun 300: 965–971. | Article | PubMed | ISI | ChemPort |
Zammit, PS, Golding, JP, Nagata, Y, Hudon, V, Partridge, TA and Beauchamp, JR (2004). Muscle satellite cells adopt divergent fates: a mechanism for self-renewal? J Cell Biol 166: 347–357. | Article | PubMed | ISI | ChemPort |
Dedkov, EI, Kostrominova, TY, Borisov, AB and Carlson, BM (2003). MyoD and myogenin protein expression in skeletal muscles of senile rats. Cell Tissue Res 311: 401–416. | PubMed | ISI | ChemPort |
Grounds, MD (1998). Age-associated changes in the response of skeletal muscle cells to exercise and regeneration. Ann NY Acad Sci 854: 78–91. | Article | PubMed | ChemPort |
Welle, S (2002). Cellular and molecular basis of age-related sarcopenia. Can J Appl Physiol 27: 19–41. | PubMed | ISI | ChemPort |
Shefer, G, Van de Mark, DP, Richardson, JB and Yablonka-Reuveni, Z (2006). Satellite-cell pool size does matter: defining the myogenic potency of aging skeletal muscle. Dev Biol 294: 50–66. | Article | PubMed | ISI | ChemPort |
Sharma, M, Kambadur, R, Matthews, KG, Somers, WG, Devlin, GP, Conaglen, JV, et al. (1999). Myostatin, a transforming growth factor-beta superfamily member, is expressed in heart muscle and is upregulated in cardiomyocytes after infarct. J Cell Physiol 180: 1–9. | Article | PubMed | ISI | ChemPort |
Rosenblatt, JD, Lunt, AI, Parry, DJ and Partridge, TA (1995). Culturing satellite cells from living single muscle fiber explants. In Vitro Cell Dev Biol Anim 31: 773–779. | Article | PubMed | ISI | ChemPort |
Partridge, TA (1997). Tissue culture of skeletal muscle. Methods Mol Biol 75: 131–144. | PubMed | ChemPort |
Yablonka-Reuveni, Z, Rudnicki, MA, Rivera, AJ, Primig, M, Anderson, JE and Natanson, P (1999). The transition from proliferation to differentiation is delayed in satellite cells from mice lacking MyoD. Dev Biol 210: 440–455. | Article | PubMed | ISI | ChemPort |
Oliver, MH, Harrison, NK, Bishop, JE, Cole, PJ and Laurent, GJ (1989). A rapid and convenient assay for counting cells cultured in microwell plates: application for assessment of growth factors. J Cell Sci 92: 513–518. | PubMed | ISI |
![]()
Suresh, A and Sodhi, A (1991). Production of interleukin-1 and tumor necrosis factor by bone marrow-derived macrophages: effect of cisplatin and lipopolysaccharide. Immunol Lett 30: 93–100. | Article | PubMed | ISI | ChemPort |
