

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Семена из WT и arp -/- растений высевали в агаризованную MС среду, содержащей ДНК-повреждающие агенты. Как показано на рисунке 4А и 4С, когда семена выращивали в среде содержащей Н2О2, не наблюдалось существенная разница в скоростях прорастания между растениями дикого типа и arp–/– растениями. Однако прорастание arp–/– растений было гораздо более чувствительным к MMS и t-BuO2H по сравнению с растениями дикого типа (Рисунок 4B и 4C). Эти результаты свидетельствуют о том, что дефицит ARP значительно повышает чувствительность растений A. thaliana к повреждениям ДНК, генерируемым MMS и t-BuO2H, но не к тем, которые генерируются Н2О2.
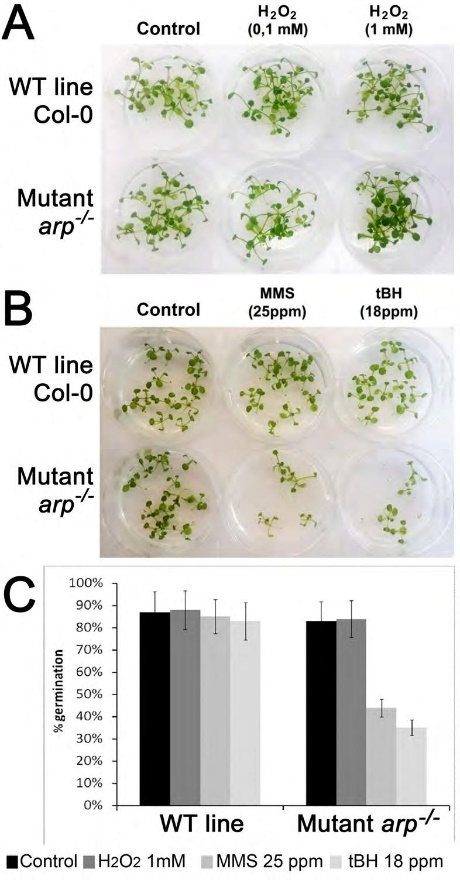
Агаризованные среды MS содержали разные концентрации H2O2 (A), MMS и t-BuO2H (B). Эффективность прорастания семян на 17 день (C)
Рисунок 4 – Чувствительность гомозиготного arp-/- мутантного растения A. thaliana к действию H2O2, MMS и t-BuO2H
Для изучения чувствительности мутантных растений к повреждению ДНК в ходе прорастания семян, мы инкубировали 6-дневные проростки дикого и arp-/- мутантного растения в среде, содержащей различные концентрации H2O2, MMS и t-BuO2H. Через 5-6 дней в среде с возрастающей концентрацией H2O2 не наблюдалось существенная разница в чувствительности к повреждению ДНК, вызванному H2O2, между растениями дикого типа и мутантом (Рисунок 5), хотя оба показали умеренное ингибирование роста при максимальной дозе H2O2 (5 мМ).
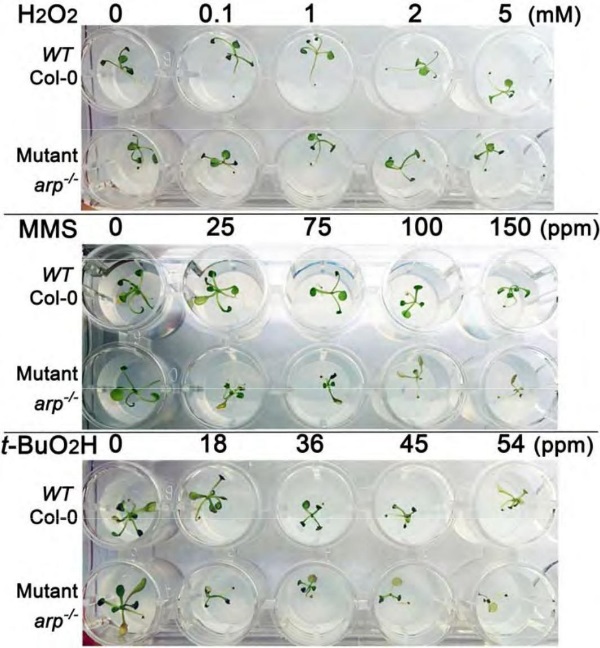
Рисунок 5 – Чувствительность гомозиготного arp-/- мутантного растения A. thaliana к действию H2O2, MMS и t-BuO2H
С другой стороны, растения дикого типа и arp-/- растения проявили дифференциальную чувствительность к MMS и t-BuO2H, при этом мутантные растения накаутные по АП-эндонуклеазе были значительно более чувствительными, чем растения дикого типа в указанном диапазоне концентрации ДНК-повреждающих агентов: 25-150 ppm для MMS и 18-54 ppm для t-BuO2H (Рисунок 5).
ЗАКЛЮЧЕНИЕ
В данной работе мы исследовали роль ARP в обеспечении устойчивости растений к действию генотоксических агентов. Мы показали, что гомозиготный arp–/– мутант более чувствителен к воздействию MMS и t-BuO2H, но не к H2O2, по сравнению с растениями дикого типа, что указывает на то, что ARP-катализируемая NIR активность требуется для репарации AП-сайтов, генерируемых экзогенными факторами и специфическими окисидативными повреждениями ДНК, индуцированными t-BuO2H в условиях in vivo. В этом отношении ARP напоминает бактериальную AP-эндонуклеазу Nfo, делеция которой также придает дифференциальную чувствительность мутантным по гену Nfo E. coli к ДНК-повреждающим агентам (Ishchenko A. A., 2006: 2564-69). Мы можем предположить, что ARP может индуцироваться в ответ на повреждение ДНК, вызванное экзогенными факторами. Так как NIR-дефицитные клетки чувствительны к воздействию t-BuO2H, но не к H2O2, можно предположить, что органический гидропероксид индуцирует специфические типы повреждения ДНК, которые удаляются по NIR механизму. При восстановлении t-BuO2H переходными металлами образуются алкоксильные радикалы (CH3)3CO•, которые, в свою очередь, инициируют перекисное окисление липидов (LPO). Интересно, что в отличие от H2O2, t-BuO2H продуцирует одноцепочечные разрывы ДНК в интактных клетках, но не в лизированных клетках, что указывает на то, что реакция с мембранными липидами участвует в формировании разрыва цепи ДНК. Действительно, высвобождение малонового диальдегида, продукта LPO, было обнаружено в клетках, инкубированных с t-BuO2H, но не с H2O2 (Baker M. A., 1991: 563-72). Реактивные альдегиды, образуемые из LPO, генерируют множество высокотоксичных экзоциклических аддуктов ДНК (Gros L., 2003: 219-29). Кроме того, распад t-BuO2H приводит к образованию трет-бутоксильных и углеродсодержащих метильных радикалов. Последний радикал может атаковать ДНК и продуцировать 8-метилгуанин (8meG) (Hix S., 1995:293-301). Остатки 8meG могут быть обнаружены in vivo в ДНК, выделенной из печени и желудка крыс, обработанных t-BuO2H (Hix S., 2000: P. 1056-4). E. coli 3-метиладенин ДНК-гликозилаза AlkA вырезает 8meG из дуплексной ДНК с высокой эффективностью из пар с аденином или тимином, но делает это очень плохо, когда 8meG находится в естественном контексте против цитозина (Gasparutto D., 2002: 437-47). В клетках млекопитающих обнаружена активность 8meG-ДНК гликозилазы, что свидетельствует о существовании альтернативных путей восстановления t-BuO2H индуцированных поражений ДНК. Исходя из этих наблюдений, мы выдвигаем гипотезу о том, что повреждения ДНК, образующиеся органическим пероксидом в растениях, являются субстратами для ARP-катализируемого NIR-пути.
Ранее было показано, что АП-эндонуклеаза инициированный NIR механизм работает совместно с BER механизмом в репарации геномной ДНК от потенциально мутагенных и летальных повреждений (Ishchenko A. A., 2006: 2564-69). В настоящем исследовании мы показали, что NIR функция сохраняется у двудольных растений, которые содержат гены кодирующие гомологи ARP белка. Биохимические и генетические данные показывают, что NIR механизм является универсальным и сохраняется у всех эукариот, включая дрожжи, нематоды, млекопитающие и цветковые растения. В заключение мы предполагаем, что катализируемый АП-эндонуклеазой NIR-механизм играет существенную роль в поддержании целостности генома, несмотря на наличие множества универсальных ДНК гликозилаз, которые способны устранить широкий спектр поврежденных оснований ДНК.
Литература
Cadet J., Douki T., Gasparutto D., Ravanat J. L. Oxidative damage to DNA: formation, measurement and biochemical features // Mutat. Res. – 2003. – Vol. 531. – P. 5-23. Krokan H. E., Bjoras M. Base excision repair // Cold Spring Harb. Perspect. Biol. – 2013. – Vol. 5. – P. a012583. Ischenko A. A., Saparbaev M. K. Alternative nucleotide incision repair pathway for oxidative DNA damage // Nature. – 2002. – Vol. 415. – P. 183-187. Hitomi K., Iwai S., Tainer J. A. The intricate structural chemistry of base excision repair machinery: implications for DNA damage recognition, removal, and repair // DNA Repair (Amst). – 2007. - Vol. 6. – P. 410-428. Gros L., Ishchenko A. A., Ide H., Elder R. H., Saparbaev M. K. The major human AP endonuclease (Ape1) is involved in the nucleotide incision repair pathway // Nucleic Acids Res. – 2004. – Vol. 32. - P. 73-81. Friedberg E. C., Walker G. C., Siede W., Wood R. D., Schultz R. A., Ellenberger T. DNA repair and mutagenesis // ASM Press. – 2006. Vanyushin B. F., Ashapkin V. V. DNA methylation in higher plants: past, present and future // Biochim Biophys Acta. – 2011. – Vol. 1809. – P. 360-368. He X. J., Chen T., Zhu J. K. Regulation and function of DNA methylation in plants and animals // Cell Res. – 2011. – Vol. 21. – P. 442-465. Zhu J. K. Active DNA demethylation mediated by DNA glycosylases // Annu. Rev. Genet. – 2009. – Vol. 43. – P. 143-166. Morales-Ruiz T., Ortega-Galisteo A. P., Ponferrada-Marin M. I., Martinez-Macias M. I., Ariza R. R., Roldan-Arjona T. Demeter and repressor of silencing 1 encode 5-methylcytosine DNA glycosylases // Proc. Natl. Acad. Sci. U. S.A. – 2006. – Vol. 103. - P. 6853-6858. Babiychuk E., Kushnir S., Van Montagu M., Inze D. The Arabidopsis thaliana apurinic endonuclease Arp reduces human transcription factors Fos and Jun // Proc. Natl. Acad. Sci. U. S. A. – 1994. – Vol. 91. – P. 3299-3303. Cordoba-Canero D., Roldan-Arjona T., Ariza R. R. Arabidopsis ARP endonuclease functions in a branched base excision DNA repair pathway completed by LIG1 // Plant J. – 2011. – Vol. 68. – P. 693-702. Joldybayeva B., Prorok P., Grin I. R., Zharkov D. O., Ishenko A. A., Tudek B., Bissenbaev A. K., Saparbaev M. Cloning and Characterization of a Wheat Homologue of Apurinic/Apyrimidinic Endonuclease Ape1L // PLoS One. – 2014. – Vol. 9. – P. e92963. Bissenbaev A. K., Ishchenko A. A., Taipakova S. M., Saparbaev M. K. Presence of base excision repair enzymes in the wheat aleurone and their activation in cells undergoing programmed cell death // Plant Physiol. Biochem. – 2011. – Vol. 49. - P. 1155-1164. Ishchenko A. A., Deprez E., Maksimenko A., Brochon J. C., Tauc P., Saparbaev M. K. Uncoupling of the base excision and nucleotide incision repair pathways reveals their respective biological roles // Proc. Natl. Acad. Sci. U. S. A. – 2006. - Vol. 103. P. 2564-2569. Baker M. A., He S. Q. Elaboration of cellular DNA breaks by hydroperoxides // Free Radic Biol Med. – 1991. - Vol. 11. – P. 563-572. Gros L., Ishchenko A. A., Saparbaev M. Enzymology of repair of etheno-adducts // Mutat. Res. – 2003. – Vol. 531. – P. 219-229. Hix S., Morais Mda S., Augusto O. DNA methylation by tert-butyl hydroperoxide-iron (II) // Free Radic Biol Med. – 1995. – Vol. 19. – P. 293-301. Hix S., Kadiiska M. B., Mason R. P., Augusto O. In vivo metabolism of tert-butyl hydroperoxide to methyl radicals. EPR spin trapping and DNA methylation studies // Chem Res Toxicol. – 2000. – Vol. 13. – P. 1056-1054. Gasparutto D., Dherin C., Boiteux S., Cadet J. Excision of 8-methylguanine site-specifically incorporated into oligonucleotide substrates by the AlkA protein of Escherichia coli // DNA Repair (Amst). – 2002. - Vol. 1. P. 437-447. Bradford M. M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding // Anal. Biochem. – 1976. – Vol. 72. – P. 248-254. Murphy T. M., Belmonte M., Shu S., Britt A. B., Hatteroth J. Requirement for abasic endonuclease gene homologues in Arabidopsis seed development // PLoS One. – 2009. – Vol. 4. – P. e4297. Foyer C. H., Noctor G. Redox sensing and signaling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria // Physiol. Plant. – 2003. – Vol. 119. – P. 355–364. Jaruga P., Dizdaroglu M. 8,5'-Cyclopurine-2'-deoxynucleosides in DNA: mechanisms of formation, measurement, repair and biological effects // DNA Repair (Amst). – 2008. – Vol. 7. – P. 1413-1425. Brooks P. J. The 8,5'-cyclopurine-2'-deoxynucleosides: candidate neurodegenerative DNA lesions in xeroderma pigmentosum, and unique probes of transcription and nucleotide excision repair // DNA Repair (Amst). – 2008. – Vol. 7. – P. 1168-1179.
References
Cadet J., Douki T., Gasparutto D., Ravanat J. L. (2003) Oxidative damage to DNA: formation, measurement and biochemical features. Mutat. Res., vol. 531, pp. 5-23. Krokan H. E., Bjoras M. (2013) Base excision repair. Cold Spring Harb. Perspect. Biol., vol. 5, pp. a012583. Ischenko A. A., Saparbaev M. K. (2002) Alternative nucleotide incision repair pathway for oxidative DNA damage. Nature, vol. 415, pp. 183-87. Hitomi K., Iwai S., Tainer J. A. (2007) The intricate structural chemistry of base excision repair machinery: implications for DNA damage recognition, removal, and repair. DNA Repair (Amst), vol. 6, pp. 410-28. Gros L., Ishchenko A. A., Ide H., Elder R. H., Saparbaev M. K. (2004) The major human AP endonuclease (Ape1) is involved in the nucleotide incision repair pathway. Nucleic Acids Res., vol. 32, pp. 73-81. Friedberg E. C., Walker G. C., Siede W., Wood R. D., Schultz R. A., Ellenberger T. (2006) DNA repair and mutagenesis. ASM Press. Vanyushin B. F., Ashapkin V. V. (2011) DNA methylation in higher plants: past, present and future. Biochim Biophys Acta., vol. 1809, pp. 360-68. He X. J., Chen T., Zhu J. K. (2011) Regulation and function of DNA methylation in plants and animals. Cell Res., vol. 21, pp. 442-65. Zhu J. K. (2009) Active DNA demethylation mediated by DNA glycosylases. Annu. Rev. Genet., vol. 43, pp. 143-66. Morales-Ruiz T., Ortega-Galisteo A. P., Ponferrada-Marin M. I., Martinez-Macias M. I., Ariza R. R., Roldan-Arjona T. (2006) Demeter and repressor of silencing 1 encode 5-methylcytosine DNA glycosylases. Proc. Natl. Acad. Sci. U. S.A., vol. 103, pp. 6853-58. Babiychuk E., Kushnir S., Van Montagu M., Inze D. (1994) The Arabidopsis thaliana apurinic endonuclease Arp reduces human transcription factors Fos and Jun. Proc. Natl. Acad. Sci. U. S. A., vol. 91, pp. 3299-303. Cordoba-Canero D., Roldan-Arjona T., Ariza R. R. (2011) Arabidopsis ARP endonuclease functions in a branched base excision DNA repair pathway completed by LIG1. Plant J., vol. 68, pp. 693-702. Joldybayeva B., Prorok P., Grin I. R., Zharkov D. O., Ishenko A. A., Tudek B., Bissenbaev A. K., Saparbaev M. (2014) Cloning and Characterization of a Wheat Homologue of Apurinic/Apyrimidinic Endonuclease Ape1L. PLoS One, vol. 9, pp. e92963. Bissenbaev A. K., Ishchenko A. A., Taipakova S. M., Saparbaev M. K. (2011) Presence of base excision repair enzymes in the wheat aleurone and their activation in cells undergoing programmed cell death. Plant Physiol. Biochem., vol. 49, pp. 1155-64. Ishchenko A. A., Deprez E., Maksimenko A., Brochon J. C., Tauc P., Saparbaev M. K. (2006) Uncoupling of the base excision and nucleotide incision repair pathways reveals their respective biological roles. Proc. Natl. Acad. Sci. U. S. A., vol. 103, pp. 2564-69. Baker M. A., He S. Q. (1991) Elaboration of cellular DNA breaks by hydroperoxides. Free Radic Biol Med., vol. 11, pp. 563-72. Gros L., Ishchenko A. A., Saparbaev M. (2003) Enzymology of repair of etheno-adducts. Mutat. Res. vol. 531, pp. 219-29. Hix S., Morais Mda S., Augusto O. (1995) DNA methylation by tert-butyl hydroperoxide-iron (II). Free Radic Biol Med. vol. 19, pp. 293-301. Hix S., Kadiiska M. B., Mason R. P., Augusto O. (2000) In vivo metabolism of tert-butyl hydroperoxide to methyl radicals. EPR spin trapping and DNA methylation studies. Chem Res Toxicol., vol. 13, pp. 1056-54. Gasparutto D., Dherin C., Boiteux S., Cadet J. (2002) Excision of 8-methylguanine site-specifically incorporated into oligonucleotide substrates by the AlkA protein of Escherichia coli. DNA Repair (Amst), vol. 1, pp. 437-47. Bradford M. M. (1976) A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem., vol. 72, pp. 248-54. Murphy T. M., Belmonte M., Shu S., Britt A. B., Hatteroth J. (2009) Requirement for abasic endonuclease gene homologues in Arabidopsis seed development. PLoS One, vol. 4, pp. e4297. Foyer C. H., Noctor G. (2003) Redox sensing and signaling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol. Plant, vol. 119, pp. 355–64. Jaruga P., Dizdaroglu M. (2008) 8,5'-Cyclopurine-2'-deoxynucleosides in DNA: mechanisms of formation, measurement, repair and biological effects. DNA Repair (Amst), vol. 7, pp. 1413-25. Brooks P. J. (2008) The 8,5'-cyclopurine-2'-deoxynucleosides: candidate neurodegenerative DNA lesions in xeroderma pigmentosum, and unique probes of transcription and nucleotide excision repair. DNA Repair (Amst), vol. 7, pp. 1168-79.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
