
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
к. б. н., доцент, ФГБОУ ВО «Мордовский государственный
педагогический институт им. », г. Саранск
ОБ ОТРАСТАНИИ ЗЛАКОВ СРЕДНЕГО ПОВОЛЖЬЯ
Gorchakova A. Yu.
Candidate of Science, associate professor of Federal State – Financed Educational Insritution of Higher Professional Education M. E. Evsevjev Mordvian State Pedagogical Institute, Saransk
ABOUT THE REGROWTH OF CEREALS
АННОТАЦИЯ
В статье рассматриваются особенности отрастания злаков: дана схема размещения почек возобновления у злаков, определены типы побегов, образованных этими группами почек. Выделены группы злаков по характеру отрастания. Определено соотношение органов запаса при отрастании. Лето благоприятствует развитию жароустойчивых злаков, особи которых способны быстро разрастаться за счет укоренения в узлах столоновидных побегов. Определена динамика питательных веществ: протеина, моносахаров, дисахаров, крахмала. Установлено, что увеличение углеводов в органах запаса идет на первых порах за счет повышения моносахаров и дисахаров.
ABSTRACT
Features of aftergrowth of grasses are considered: the scheme of placement of buds of renewal of grasses is given, types of the sprouts that are formed by these groups of buds are defined. Groups of grasses are defined basing on nature of aftergrowth. The ratio of storage organs at aftergrowth is defined. Dynamics of nutrients is determined: protein, monosaccharides, disaccharides, starch. It is established that the increase of carbohydrates in storage organs takes place at the beginning due to increase of monosaccharides and disaccharides.
Ключевые слова: Poaceae, злаки, отрастание, почки возобновления, типы побегов, органы запаса, динамика питательных веществ.
Keywords: Poaceae, grasses, aftergrowth, renewal buds, types of sprouts, storage organs, dynamics of nutrients.
Злаки (Poaceae) составляют основу большинства лугов всех ботанико-географических зон благодаря высокому уровню адаптивности [8]. После скашивания или стравливания злаки отрастают, восстанавливают свою надземную массу. Подрастающая трава называется отавой, а свойство растений образовывать ее – отавностью. Благодаря отавности растений можно в течение лета использовать сенокос повторно, а пастбища многократно. На сенокосах и пастбищах отава в основном формируется благодаря отрастанию укороченных побегов или образованию из почек новых побегов. У растений с укороченными побегами после скашивания или стравливания остается большая поверхность листьев, находящихся вблизи поверхности почвы, по сравнению с растениями, имеющими удлиненные побеги.
Особенности отрастания злаков изучены еще недостаточно, имеющиеся публикации больше отражают особенности отрастания тропических и субтропических злаков [1; 4; 5; 9, с. 592]. В связи с этим особый интерес представляет исследование отрастания злаков, характеризующихся многообразием биоморф и составляющих во многих районах основную хозяйственно-ботаническую группу в урожае пастбищ [3; 10;12].
Целью наших исследований являлось изучение особенностей отрастания злаков. В задачу исследований входило выделение групп злаков по характеру отрастания и установление динамики питательных веществ, сопровождающих этот процесс.
МАТЕРИАЛ И МЕТОДЫ
Для эксперимента использовались важнейшие в хозяйственном отношении кормовые злаки. Маршрутные обследования, экспедиции, которые составили значительную часть нашей работы, проводились по территории Республики Мордовия. Также были проведены стационарные наблюдения за кормовыми и зерновыми злаками путем постоянного отбора проб и определения биометрических показателей. В некоторых опытах каждые 15 дней извлекаются целые растения (до 10−12 экземпляров каждого вида), описываются и материал фиксируется для детального изучения. За основу нами принимается методика (1954) [6]. Биохимические характеристики изучали по общепринятым методикам. Данные по динамике сахаров, протеина (их увеличение или уменьшение) служили в качестве основного показателя интенсивности обмена в растениях. Все показатели определяли по мере завершения формирования отдельных фитомеров (по закончившим рост листьям) до перехода растений в фазу кущения.
РЕЗУЛЬТАТЫ
Несмотря на большие видовые различия в формировании почек особями, прослеживается определенная закономерность в характере отрастания злаков, проявляющаяся прежде всего в долевом участии различных групп почек в возобновлении их травостоя, о чем свидетельствуют данные наших опытов в Мордовии (травостой срезали на высоте 6-8 см с последующим подсчетом числа образовавшихся побегов на 100 отросших в 5- 6-кратной повторности) в летний период (рисунок 1, таблица 1).
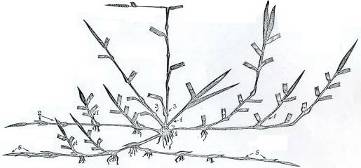
Рисунок 1 – Схема размещения почек возобновления у бореальных злаков: 1-5 – группы почек.
Таблица 1
Отрастание некоторых злаков в летний период.
Вид | Жизнен - ная форма | Побеги, образованные группами почек, в %, ( | ||||
1-й | 2-й | 3-й | 4-й | 5-й | ||
Dactylis glomerata L. | Р | 69 ± 4,9 | 11 ± 0,6 | 1 ± 0,2 | 19 ± 1,3 | ─ |
Festuca rubra L. | КР | 61 ± 1,0 | ─ | 8 ± 0,5 | 31 ± 0,9 | ─ |
Phleum pratense L. | Р | 2 ± 0,3 | 3 ± 0,4 | 48 ±1,3 | 47 ± 1,1 | ─ |
Deschampsia cespitosa (L.) Beauv. | П | 87 ± 0,9 | 4 ± 0,4 | 1 ± 0,3 | 8 ± 0,7 | ─ |
Bromopsis inermis (Leyss.) Holub, сорт «Пензенский-1» | К | 6 ± 0,5 | ─ | 18 ± 0,7 | 50 ± 1,3 | 26 ± 1,1 |
Calamagrostis epigeios (L.) Roth | К | 1 ± 0,1 | ─ | 30 ± 1,5 | 26 ± 0,7 | 43 ± 0,7 |
Agrostis stolonifera L. | КС | 1 ± 0,1 | 18 ± 0,3 | 67 ± 1,5 | 3 ± 0,3 | 11 ± 0,4 |
Примечание: К – корневищные; КС – корневищно-столонообразующие; КР – корневищно-рыхлодерновинные; Р – рыхлодерновинные; П – плотнодерновинные.
После отчуждения злаки отрастают по-разному: одни виды возобновляются главным образом почками надземными, другие – подземными. Например, злаки, формирующие корневища (Bromopsis inermis, Calamagrostis epigeios и др.), в основном отрастают почками подземных структур, а виды, имеющие в структуре особей большое количество укороченных и столоновидных побегов (Agrostis stolonifera L., Phleum pratense L. и т. д.), отрастают надземными группами почек. Апикальные почки имеют большое значение у 44 % изученных видов в Мордовии. Определяющая роль в формировании отавы принадлежит этим почкам у таких видов как Deschampsia cespitosa (L.) Beauv., Dactylis glomerata L., Phleum pratense и др., в структуре которых преобладают укороченные побеги. Значение боковых почек диагеотропных побегов в формировании отавы, несмотря на наличие этих структур у многих видов, небольшое. Значительная роль принадлежит им в отрастании столонообразующих злаков.
Боковые почки апогеотропных побегов имеют большое значение в формировании отавы у многих злаков. Например, у Agrostis stolonifera эти почки давали в Мордовии около 67 % побегов. Велика их роль в восстановлении травостоя у высорослых злаков, не образующих укороченных побегов (Bromopsis inermis, Calamagrostis epigeios и т. д.) – свыше 20 %. Значение почек зоны кущения в восстановлении травостоя злаков весьма большое, хотя доля участия в отрастании отавы сильно варьирует по видам. Наибольшое число побегов формируют почки зоны кущения у таких видов как Dactylis glomerata и др. На долю почек корневищ в формировании отавы высокорослых корневищных злаков приходится до 50 %.
В условиях Мордовии доля почек диагеотропных побегов в формировании отавы злаков была также невысокой. Это связано с более интенсивным захватом сопредельной территории каждой особью в связи с повышением температуры в летний период (до 300 и выше), улучшением условий вегетации для злаков с С4-типом фотосинтеза и ослаблением конкуренции со стороны вегетировавших ранней весной видов умеренных широт. Высокая температура лета благоприятствует развитию жароустойчивых злаков, особи которых способны быстро разрастаться за счет укоренения в узлах столоновидных побегов.
Анализ структуры отавы злаков показывает, что различия в отрастании отдельных видов связаны с особенностями их побегообразования: чем больше типов побегов в структуре особей, тем больше групп почек принимает участие в формировании отавы. Отрастание отдельных злаков после дефолиации зависит от доли укороченных и столоновидных побегов, ростовые почки которых находятся, как правило, ниже уровня отчуждения. Прекращение роста побегов наступало всякий раз, когда срез приходился ниже верхнего узла. Удаление верхней части побега, где концентрируется активная меристема, парализует его рост и даже приводит к гибели. В период роста материнского побега многие почки зоны кущения, а также надземных фитомеров покоятся и не раскрываются. Срезание верхушки приводит к переходу в рост ранее покоящихся боковых почек. Удаление апекса материнского побега активизирует также рост побегов из почек корневищ и столонов.
Таким образом, по характеру отрастания злаки можно разделить на следующие группы: 1) отрастающие при низком срезе в основном подземными почками, не формирующие укороченных побегов и после отчуждения теряющие всю листовую поверхность (корневищные виды); 2) формирующие укороченные побеги с длинными влагалищами (их апикальные почки размещаются на поверхности почвы и нередко отчуждаются), которые частично сохраняют ассимиляционную поверхность после среза и отрастают сохранившимися апикальными почками и почками зоны кущения (высокорослые рыхлодерновинные виды); 3) отрастающие в основном апикальными почками, выделяющиеся сохранением значительной части листовой поверхности после отчуждения (низкорослые пастбищные злаки).
Характер отрастания злаков определяется также количеством и составом питательных веществ в органах запаса, которые расходуются на дыхание и построение новых структур, на что указывают многие авторы [9; 12; 15]. Наиболее динамичными веществами являются углеводы стерни, которые представлены ассимилянтами и используются в качестве энергетического материала для образования новых органов. Гидролиз углеводов других органов происходит несколько позже. Интенсивность вегетативного возобновления злаков, как и других растений, определяется, особенно в первое время после дефолиации, количеством запасных веществ, накопленных каждым растением [7; 15]. Об этом наглядно свидетельствуют также наши исследования динамики протеина (рисунок 2), моносахаров, дисахаров и крахмала после скашивания злаков в летний период в Мордовии.
Сразу после скашивания наблюдается снижение растворимых углеводов сначала в стерне, а затем и в подземных органах. По мере роста листьев и усиления фотосинтеза увеличивается количество растворимых углеводов.
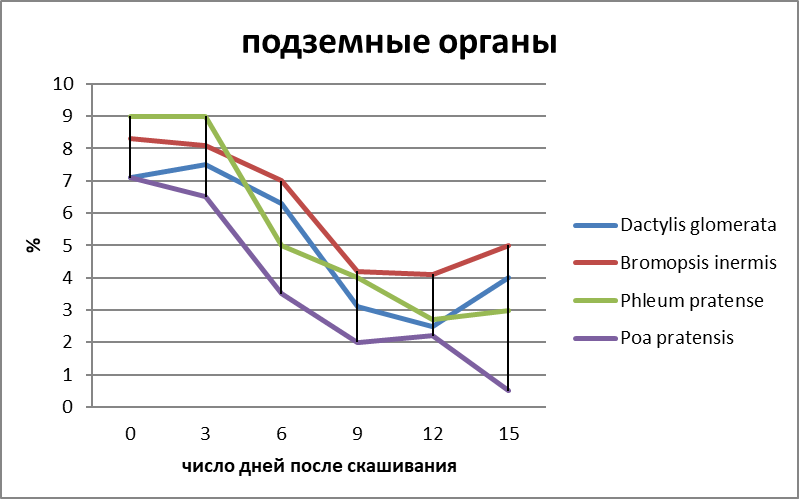
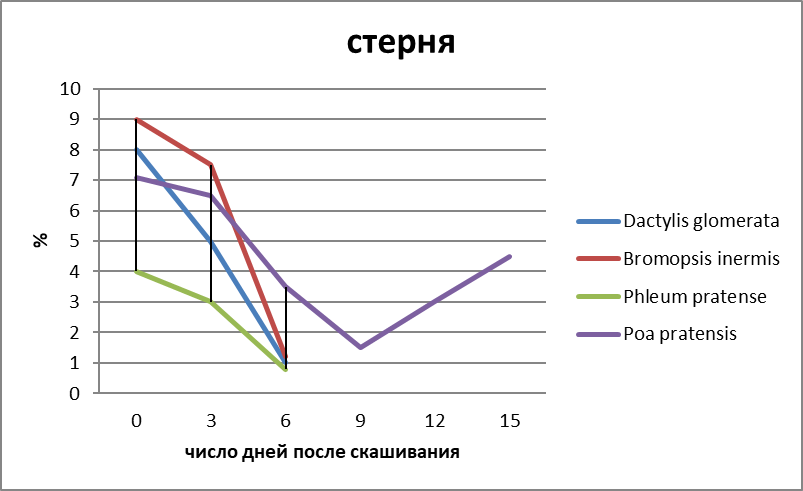
Рисунок 2 – Динамика протеина в запасающих органах некоторых злаков (Мордовия, 2011г., в %).
Первые три дня после отчуждения характеризуется резким снижением в содержании углеводов в надземных структурах всех видов, в то время как в подземной части их количество мало изменяется. В этот период, очевидно, интенсивно протекают процессы гидролиза и расходования углеводов на дыхание. Процессы роста проходят еще слабо – из апикальных почек у всех видов появляется только первый свернутый лист. Содержание протеина практически не меняется во всех структурах. Быстрой мобилизацией запасных веществ стерни, очевидно, можно объяснить раннее отрастание укороченных побегов и пробуждение сохранившихся боковых почек апогеотропных побегов.
Через 6 дней после скашивания в период развертывания первого листа в надземной части содержание моно - и дисахаров снижалось до минимума у всех видов, а протеина – в 2-3 раза; в подземных органах заметно уменьшилось количество крахмала, особенно у Dactylis glomerata. Слабое снижение моно - и дисахаров в корнях, очевидно, связано с усилением гидролиза крахмала. Отмечено также снижение протеина. Полученные данные указывают на то, что запасной материал продолжает расходоваться, поскольку продуктов фотосинтеза, образуемых в молодых листьях, явно еще недостаточно для формирования новых побегов. Через 9 дней после среза (развертывание второго листа) отмечено дальнейшее снижение количества крахмала, что связано с усилением расходов пластического материала на построение новых органов и отмиранием части старых корней. К 12 дню новые побеги всех злаков имели по 2-3 развернутых листа, а у Bromopsis inermis – 4; отмечено снижение водорастворимых сахаров и крахмала. Спустя 15 дней после скашивания у отрастающих побегов появились четвертый и шестой листья, и наметилась стабилизация в содержании углеводов, а у некоторых видов отмечено увеличение простых сахаров. Последнее, очевидно, связано с поступлением сахаров из надземных органов, где их образование заметно повысилось в связи с усилением фотосинтеза.
Различный ход изменения в содержании углеводов и их фракций по видам в период отрастания отавы можно объяснить в известной степени данными соотношения органов запаса у изучаемых видов (рисунок 3).
В среднем на 100 побегов у Phleum pratense образуется (по массе) больше корней и корневищ, чем у других видов. Поэтому стабилизация в содержании запасных углеводов у этого вида наметилась уже к 9-12 дню после среза. Если, например, Poa pratensis L. s. l. на 100 побегов приходится всего 115 г сухих веществ запасающих органов, то у Phleum pratense – 409.
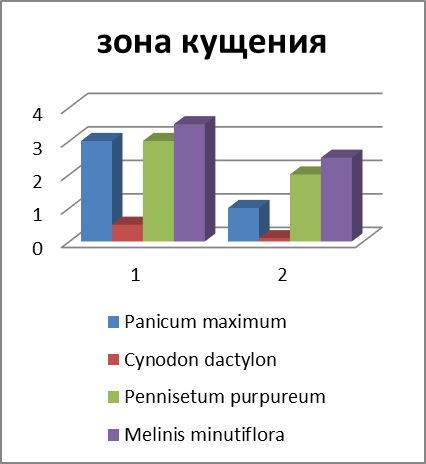
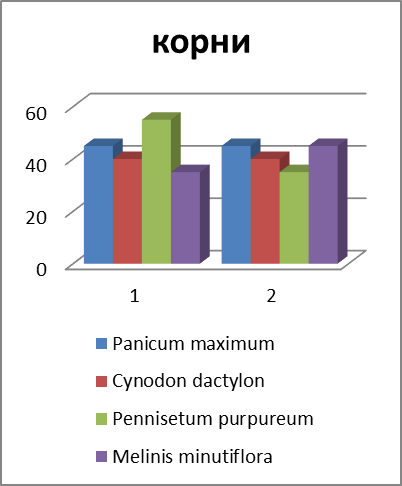
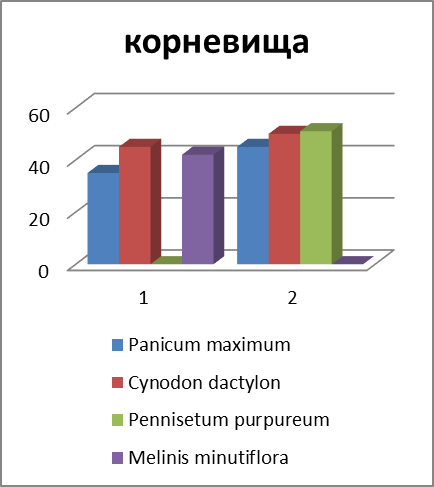
Рисунок 3 – Соотношение массы отдельных запасающих органов у некоторых злаков (Мордовия, 2011 г., в %).
Скашивание заметно влияет на соотношение органов запаса. Наибольшему изменению подвергаются показатели массы стерневых остатков, зоны кущения и корней. У Poa pratensis, у которой корни являются важнейшим источником запаса, через полмесяца после срезания масса активных корней снижается более чем в два раза; у Bromopsis inermis и Phleum pratense масса корней снижается менее заметно, что связано, очевидно, с продолжением активной деятельности корней, образованных корневищами. Высокий процент стерневых остатков у Poa pratensis объясняется продолжением их вегетации после среза в связи с образованием боковых побегов.
ОБСУЖДЕНИЕ
Отрастание растений зависит от их биологических свойств, времени скашивания (стравливания), условий произрастания, степени обеспеченности растений запасными питательными веществами. Анализ динамики углеводов в запасающих органах бореальных злаков показывает, что их расход в первый период после отчуждения (период интенсивного дыхания) выше, чем протеина; наиболее активно расходуются углеводы стерни; вступление растений в фазу интенсивного образования новых органов (через 6-9 дней после среза) связано с продолжением расходования углеводов и заметным снижением протеина в стерне и подземных органах; стабилизация углеводов в запасающих органах и их некоторое накопление по времени совпадает с образованием побегов с 3-4 листьями; увеличение углеводов в органах запаса идет на первых порах за счет повышения моносахаров и дисахаров. Таким образом, имеющиеся публикации, которые больше отражают особенности отрастания тропических и субтропических злаков [1; 2; 3; 4; 5; 9; 13; 14], дополняются нашими исследованиями злаков умеренной зоны.
ЗАКЛЮЧЕНИЕ
Наши исследования ограничились определением отрастание злаков особенностями их побегообразования и формирования отдельных групп почек. По местоположению и роли в образовании отавы после дефолиации травостоя почки выделены на пять групп: апикальные, боковые почки диагеотропных побегов, боковые почки апогеотропных побегов, почки зоны кущения и почки корневищ, различающиеся биометрическими и биохимическими характеристиками. Характерной чертой бореальных злаков является их высокая способность к отрастанию боковыми почками надземных апогеотропных побегов. Злаки, относящиеся к группе корневищно-столонообразующих и корневищных, отличаются широким участием в формировании отавы боковых почек апогеотропных побегов (корневищ и столоновидных). Возможными перспективами для дальнейших исследований является исследование отрастания злаков по сезонам года, в зависимости от высоты и частоты среза.
Список литературы
1. Онтогенетические спектры, эффективная плотность, классификация популяций растений / // Труды международной конференции по фитоценологии и систематике высших растений, посвященной 100-летию со дня рождения. – М. : МПГУ, 2001. – С. 62–63.
2. Морфологические особенности ветвления побегов у бореальных злаков / , // Труды Кубанского государственного аграрного университета. – 2011. – Т. 1. – № 30. – С. 81-84.
3. О сезонном развитии злаков Республики Мордовия / // Бот. журн. – 2013. – Т. 98. – № 5. – С. 605-621.
4. Численность и возрастной состав ценопопуляций лисохвоста лугового (Alopecurus pratensis L.) на лугах Архангельской области / // Труды международной конференции по фитоценологии и систематике высших растений, посвященной 100-летию со дня рождения. – М. : МПГУ, 2001. – С. 93–94.
5. Восстановительные сукцессии травяных сообществ в ландшафтах южной лесостепи (Воронежская область, музей-заповедник «Дивногорье») / , // Вест. СПбГУ. Серия 7 (геология, география). – 2009. – № 2. – С. 92–96.
6. О методах изучения ритмики сезонного развития растений в стационарных геоботанических исследованиях / // Уч. записки МГПИ им. . – 1954. – Т. 37. – Вып. 2. – С. 3–20.
7. Интенсивность начального роста проростков дикорастущих злаков Stipa krylovii Roshev. и Leymus chinensis (Trin.) Tzvel. в условиях Восточного Забайкалья / // Естественные и технические науки. – 2008. – №3. – С. 82–87.
8. Проблемы теоретической морфологии и эволюции высших растений / . – М., СПб : КМК, 2005. – 407 с.
9. Brown W. V. The organization of the grass shoot apex and systematic / W. V. Brown, W. H. Emery //American Journal of Botany. –1957. – Vol. 44 – Pp. 590-595.
10. Gorchakova A. Yu., 2012. An impact of soil processing on vegetative reproduction of Agrostis Stolonifera. International Journal оf Applied and Fundamental Research. http://www. /451-24040.
11. Gorchakova A. Yu. Revisiting of grass brunching / A. Yu. Gorchakova, I. S. Belyuchenko // Bothalia Journal [Электронный ресурс] : электрон. научный журнал. – 2014. – Vol. 44. – № 8. – Aug 2014. – Pp. 163–180. – Режим доступа: http:///beheer/index. php/acceptance/index/m108N/140596214 .
12. Gorchakova A. Yu. About features of grass brunching / A. Yu. Gorchakova, I. S. Belyuchenko // Ciencia e Teccnica vitivinicola [Электронный ресурс] : электрон. научный журнал. – 2014. – Vol. 29. – № 8. – Pp. 80–98. – Режим доступа:http://cienciatecnica. org/cien/index. php/acceptance/index/Mwb8W/1406482943 .
13. Humphreys L. btropical grass growth / L. R. Humphreys // Trop. Agric. – 1966. – Vol. 23. – Pp. 337-358.
14. Jones R. J. The Method of reproduction in Rhodes grass / R. J. Jones, A. J. Pritchard //Trop. Agric. – 1971. – Vol. 4(5). – Pp. 301-307.
15. Roberts M. R. Effects of soil organic layer removal on regeneration after clear cutting a northern hardwood stand in New Brunswick / M. R. Roberts, H. Dong // Canadian Journal of Forest Research. – 1993. – Vol. 23 – Pp. 2093-2100.
