
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ГБОу гимназия 1569 «созвездие» |
Живой свет: биолюминесценция |
Баранникова Анастасия, 10а класс
Научный руководитель:
Введение
При упоминании люминесценции или флуоресценции люди представляют себе что-то, излучающее свет Ї начиная от лампы и заканчивая абсолютно абстрактным сгустком света. Если же сделать акцент на том, что источник этого света живой, то в голове всплывают светящиеся организмы, будь то мышь или собака. Однако собака Баскервилей оказалась покрыта фосфором, а вот светящиеся мыши Ї вполне реальны, и созданы они благодаря генной модификации. Например, обитают такие животные в Научном Центре Биомедицинских Технологий в Подмосковье. Каждая мышь светится холодным зеленым светом под ультрафиолетовым излучением. Меченые стволовые клетки люминесцентных особей вводят другим мышам, больным сахарным диабетом второго типа, а затем исследуют их местонахождение и процессы, происходящие внутри организма.
Сейчас ученые используют GFP и его варианты и гомологи во многих методах для изучения организации и функционирования живых систем. Флуоресцентные белки, соединившиеся с интересующими белками, позволяют наблюдать за их локализацией, передвижением, оборотом и даже «старением» (т. е. временем, прошедшим после белкового синтеза), а также множеством других процессов, связанных с жизнью клетки. Нуклеиновые кислоты тоже могут быть помечены — с помощью РНК - или ДНК-связывающих белковых производных.
Многие лаборатории сосредотачивают свои усилия на идентификации и создании флуоресцентных белков с новыми характеристиками и улучшенными свойствами: разнообразие существующих в настоящее время белков покрывает практически весь видимый спектр цветов, что позволяет одновременно использовать несколько маркеров для разных объектов.
Сам белок, на основе которого и появляется у изначально нефлуоресцирующих существ данная способность – зеленый светящийся белок (Green Fluorescent Protein, GFP). В 2008 году Нобелевская Премия по химии была присуждена за открытие, служащее поистине ярким примером того, как исследования в одной области науки могут привести к прорыву в другой. Таким образом, изучение механизма свечения морского организма неожиданно дало ключ к огромному спектру инструментов для визуализации жизнедеятельности клетки.
Цель нашего исследования – изучение явления биолюминесценции и возможность его использования в будущем в различных областях науки.
Гипотеза исследования: Использование флуоресцентных веществ и соединений в будущем позволит усовершенствовать многие технологии.
Можно выделить следующие задачи нашего исследования:
- Выявить особенности люминесцентных органических соединений и объяснить природу их происхождения Исследовать способы использования биолюминесцентных ресурсов Сделать прогноз о развитии данной отрасли науки и ее востребованности Выделить из клеток белок PFP (англ. Purple fluorescent protein, фиолетовый флуоресцентный белок) и рассмотреть его способность к люминесценции Трансформировать E. coli (кишечную палочку), внедрив плазмиду, кодирующую белок-гомолог GFP (англ. Green fluorescent protein, зеленый флуоресцентный белок)
Глава 1. Теоретическая часть
История открытия биолюминесцентных белков
Всё началось с Осами Шимомуры и его исследования в области биолюминесценции – химических реакций в живом, результатом которых является выделение света. Изучая светящихся медуз Aequorea victoria в 1960х, он выделил биолюминесцентный протеин, излучающий в синем спектре. Но медуза излучала зеленый свет. Дальнейшие исследования показали, что этот синий свет поглощался другим протеином, позднее названным «зеленым флуоресцентным белком» (англ. green fluorescent protein, GFP, 1962 год), который в свою очередь продуцирует в зеленом спектре. Способность GFP поглощать в синем и излучать в зеленом спектрах оказалась зависимой исключительно от структуры белка, без каких-либо побочных факторов.
В 1988 Мартин Чалфи услышал о GFP впервые и понял, что его способность самостоятельно флуоресцировать может сделать его идеальным клеточным маркером в организмах-моделях, которые он изучал. Используя техники молекулярной биологии, Чалфи преуспел во внедрении гена GFP в червей C. Elegans (свободноживущая нематода). GFP производился в клетках, излучая свой зеленый свет без необходимости добавления каких-либо компонентов и без какого-либо видимого ущерба, нанесенного клеткам. Чалфи поместил ген GFP после промотера (отрезка цепи генетической информации, служащего командой к началу синтеза), активного в рецепторных нейронах круглых червей в гонадах материнского червя. Так как этот вид – гермафродиты, за этим последовало самовоспроизведение особи. GFP перешел в яйца, отложенные материнским червем. Из яиц появляются новые особи, уже несущие в своем генетическом коде информацию о GFP, но продуцировать зеленый свет могут только рецепторные нейроны. Когда они синтезируют GFP, они начинают светиться в ультрафиолетовом свете.
Дальнейшая работа подтвердила, что ген GFP возможно модифицировать. Данное открытие дало ученым возможность отслеживать локализацию некоторых веществ в живых организмах.
Роджер Тсьен первым изучил, как структура GFP продуцирует зеленый свет, а затем использовал эти знания, чтобы создать молекулы, продуцирующие с другой длиной волн, следовательно – в другом цвете. Со временем его команда добавила еще несколько природных флуоресцирующих молекул к общему количеству, расширив цветовой спектр, добавив в него желтый и несколько других оттенков. Однако, у него не вышло создать красный флуоресцентный протеин. Красный свет проходит сквозь кожу и другие биологические ткани намного легче других, что делает его идеальным для лабораторных исследований in vivo.
В 1999 году русские ученые выделили из кораллов и красный вариант, DsRED. Он больше и вытянутей GFP. Тсьену удалось уменьшить его размер. Из DsRED Тсьен также получил протеины с «вкусными» названиями: mPlum, mCherry, mStrawberry, mOrange и mCitrine.
С тех пор сложные биологические системы могут помечаться различными цветами, показывая всю глубину процессов, ранее скрытых от взора исследователей.
Десятки тысяч различных белков в живых организмах контролируют важные химические процессы в мельчайших деталях. Если этот белковый механизм нарушится вследствие мутаций или искусственных модификаций, то могут проявиться различные дисфункции и болезни, в дальнейшем приводящие в том числе и к летальному исходу. Вот почему для биохимии так важно определить индивидуальную роль каждого белка в жизнедеятельности живого организма.
В 2008 году Нобелевская премия по химии была присуждена этим трем ученым за открытие GFP и серию важных исследований в области его применения и развития технологий, позволивших использовать его как инструмент в биологических науках. Используя ДНК, ученые могут связывать GFP с не менее интересными, но невидимыми белками (приложение 5).
Среди важных исследований, в которых задействован зеленый флуоресцентный белок можно найти, например, разрушение нервных клеток при болезни Альцгеймера или создание инсулин-продуцирующих бета-клеток в печени эмбриона.
Структура флуоресцентных белков
GFP-белки весьма разнообразны по спектрам поглощения и флуоресценции. Несмотря на это, все они очень близки по своей структуре.
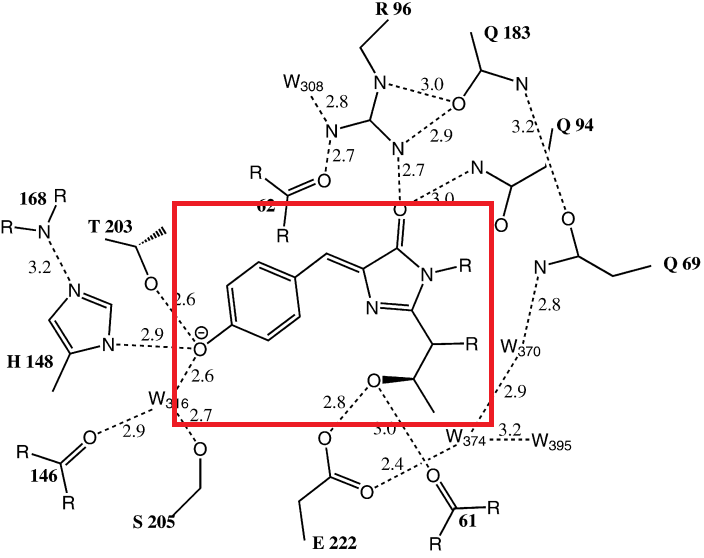
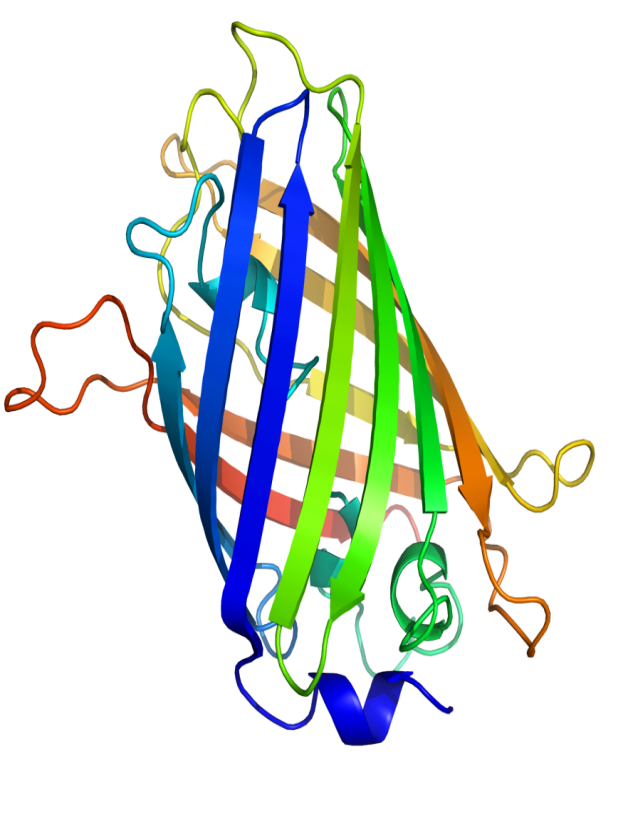
Первыми, в 1996 году, были установлены структуры GFP из A. victoria. Хотя GFP дикого типа и большинство мутантов были закристаллизованы в димерной форме, структуры некоторых мономерных белков также были установлены. GFP не является облигатным димером, это зависит от условий роста кристаллов [8].Он имеет уникальную структуру полого цилиндра, внешняя сторона которого образована одиннадцатью антипараллельными в-листами. Хромофор находится внутри структуры и связан б-спиральными участками, проходящими вдоль оси цилиндра. Короткие фрагменты б-спиралей и петли образуют также крышки, захватывающие цилиндр сверху и снизу [6]. Такая структура, характерная для цветных белков, зовется в-бочонком (приложение 3).
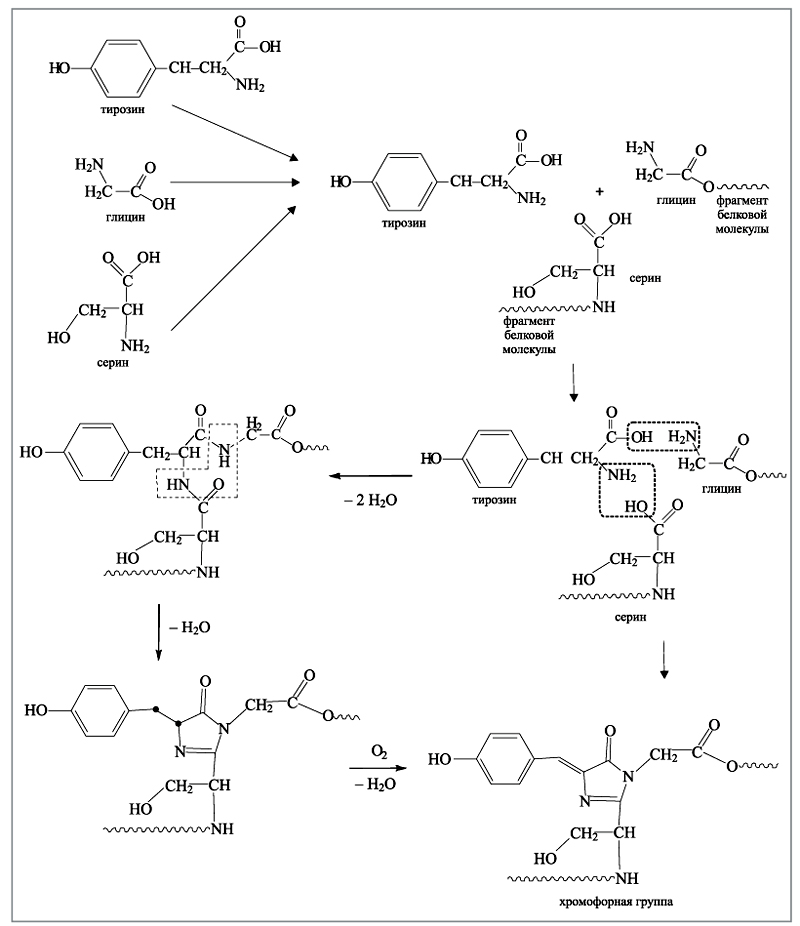
На первый взгляд кажется, что строение хромофорной группы достаточно сложное, и то, как она получается, непонятно. На самом деле ее образование – результат трех весьма простых последовательных стадий, в результате каждой из которых выделяется вода в качестве побочного продукта. Все происходит внутри пептидного цилиндра. Биохимики давно знают, что все белковые молекулы собираются внутри живого организма из известных аминокислот – органических молекул, содержащих аминогруппу NH2 и кислотную карбоксильную группу СООН. Эти аминокислоты имеют отличительный признак: между аминогруппой и карбоксильной группой находится всего один атом углерода, таким образом, общий вид всех природных аминокислот NH2–CR2–COOH. Органические заместители R у центрального атома углерода могут быть различными, в том числе и атомами водорода. При образовании белковых молекул концевая аминогруппа одной молекулы взаимодействует с концевой карбоксильной группой соседней молекулы с выделением воды:
NH2–CR2–COOH + NH2–CR'2–COOH —>NH2–CR2–CO–NH–CR'2–COOH + H2O
Именно таким образом наращивается цепочка белковой молекулы.
Хромофорная группа в GFP собирается из трех аминокислот: тирозина, глицина и серина, которые присутствуют в каждом живом организме. Три молекулы объединяются в единый реагирующий центр. Две аминокислоты из трех (глицин и серин) прикреплены (подвешены) к внутренней полости основного цилиндра. Далее все происходит следующим образом: гидроксильная группа тирозина реагирует с аминогруппой глицина, а аминогруппа тирозина взаимодействует с гидроксильной группой серина при этом выделяются две молекулы воды. Далее реагируют две аминогруппы и карбонильный кислород, образуется пятичленный цикл с двумя атомами азота и двойной связью (так называемый имидазольный цикл), вновь выделяется молекула воды. На последней стадии молекула кислорода отрывает по одному атому водорода от двух атомов углерода; выделяется молекула воды, и в итоге образуется хромофорная группа (приложения 1, 2).
Флуорофор = Ser65+Tyr66+Gly67
Флуоресцентные белки и фотосинтез у животных
Флуоресцентные белки найдены более чем в 125 видах живых существ. со своей группой исследователей выяснил, что флуоресцентные протеины могут вести себя как доноры электронов в возбужденном состоянии [2]. В их статье, опубликованной в Nature Chemical Biology, говорилось, что возбужденный зеленый хромофор (GFP) отдает электрон акцептору, создавая неустойчивое образование. Лукьянов прикрепил белок к металлическим бусинам и поместил такие бусины в раствор, содержащий различные электронакцепторы. При облучении белка голубым светом белок сначала флуоресцировал зеленым светом, затем постепенно становился красным, демонстрируя то, что он отдает электроны компонентам раствора. Если вблизи нет электронных акцепторов, то эта промежуточная форма теряет цвет. Однако, если они есть и они вступают реакцию, то GFP краснеет. Это явление называется реддингом (приложение 6). Чтобы понять, в каком из процессов, связанных с передачей электронов, задействован GFP, исследователям необходимо выяснить, кому именно он передает электрон.
Это революционная идея, так как ранее считалось, что флуоресцентные протеины пассивно поглощают свет и продуцируют его, вызывая явление биолюминесценции. Однако Лукьянов предполагает, что «природной функцией GFP может оказаться донорство электронов – аналогично донорству хлорофилла в процессе фотосинтеза.
Глава 2. Исследовательская часть
2.1 Эксперимент 1: Выделение плазмидной ДНК из бактерий E. coli
Нами был проведен ряд экспериментов по выделению плазмидной ДНК бактерии, трансформации бактерий и последующему выделению экспрессированного белка на базе межфакультетского центра Московского Государственного Университета имени в городе Пущино под руководством Чернышева Сергея Вячеславовича.
Материалы и оборудование для выделения плазмидной ДНК из E. coli:
Штамм E. coli XL1–Blue (StratageneTM), трансформированный плазмидой pBR322, микроцентрифуга, шейкер (приложение 7)
Растворы:
- Раствор I для выделения плазмидной ДНК: 50мМ глюкоза; 100мМ Tris–HCl, pH 8,0; 10мМ ЭДТА. Хранить при +4 С, перед использованием добавить лизоцим до 5 мг/мл. Раствор II. 0,2N NaOH; 1% SDS. Готовить свежий раствор необходимо перед использованием, допускается лишь непродолжительное хранение при комнатной температуре в плотно закрытой пластиковой посуде. Раствор III. 3М ацетат калия, рН 5,0. На 100 мл: 5M ацетат калия – 60 мл; CH3COOH ледяная – 11,5 мл; H2O – 28,5 мл. Смесь фенол–хлороформ Смесь хлороформ–изоамиловый спирт Ампициллин. Раствор 100 мг/мл в воде.
Методика [1]
1. Вырастить культуру клеток E. coli с плазмидой pBR322 в 5 мл питательной среды 2YT содержащей 100 мкг/мл антибиотика ампициллина, в течение 16–24 ч на термостатируемой роторной качалке-шейкере при температуре 37 С и скорости 200–300 об/мин.
2. Осадить клетки из культуральной среды. Для этого поместить 1,5 мл «ночной культуры» E. coli в 1,7 мл эппендорф и центрифугировать в течение 2 мин при 10000 об/мин. Удалить супернатант выливанием через край. Повторить осаждение в эту пробирку еще 2 раза.
3. Центрифугировать 10 с дополнительно, отобрать остатки культуральной среды микропипеткой.
4. Ресуспензировать осадок в 200 мкл раствора I с помощью микропипетки или вортекса. Оставить при комнатной температуре на 5 мин.
5. Добавить 400 мкл раствора II, сразу же резко встряхнуть, перевернуть 5 раз. Происходит лизис бактерий и щелочная денатурация ДНК. Поместить образцы на лед (0 С) на 5 мин.
6. Добавить 300 мкл холодного раствора III, 5 раз перевернуть пробирку и оставить на льду на 5 мин.
7. Центрифугировать в течение 5 мин при максимальной скорости центрифуги для осаждения хромосомной ДНК.
8. Супернатант, содержащий плазмидную ДНК, перенести микропипеткой в новую пробирку, содержащую 500 мкл изопропанола, смешать переворачиванием пробирки, оставить на 10 мин на столе.
9. Осадить плазмиду центрифугированием в течение 10 мин при максимальной скорости центрифуги, супернатант затем вылить через край, а остатки после короткого центрифугирования отобрать микропипеткой.
10. Плазмидный осадок растворить в 100 мкл TE-буфера (можно 200 мкл).
11. Раствор плазмидной ДНК необходимо подвергнуть дальнейшей очистке – провести фенольную депротеинизацию образца. Для этого добавить в пробирку равный объем смеси фенол–хлороформ, хорошо перемешать и разделить фазы центрифугированием (10 мин, 10000 об/мин).
12. Перенести верхнюю водную фазу в новую пробирку, добавить равный объем смеси хлороформ–изоамиловый спирт (помимо денатурации протеинов она удаляет остатки фенола), хорошо перемешать и разделить органическую и водную фазы центрифугированием в течение 3 мин.
13. Отобрать верхнюю водную фазу, добавить 1/10 объема холодного раствора 3М ацетата калия, pH 5,0 и высадить ДНК этанолом. Для этого добавить в пробирку 1 мл холодного (–20 С) этанола и поместить образец на лед или в морозильник не менее чем на 15 мин. На этой стадии можно прерваться: под спиртом очищенная ДНК, в принципе, может храниться годами. Но все три формы плазмиды, судя по опыту, перейдут в линейную форму.
14. Осадить плазмидную ДНК центрифугированием (10 мин, 10000 об/мин).
15. Добавить к осадку 1 мл 70% этанола для промывки осадка ДНК и центрифугировать 3 мин.
16. Слить супернатант, перевернуть пробирки на фильтр и подсушить осадок на воздухе в течение 0,5 ч. Растворить в 50–100 мкл воды. На электрофорез следует взять 5–10 мкл образца.
17. Провести при необходимости обработку образца рибонуклеазой для удаления РНК. Следует добавить к полученному препарату плазмиды 1/100 объема раствора РНКазы А (10 мг/мл в 10 мМ Tris–HCl (рН 7,4), 15 мМ NaCl) и выдержать 0,5–1 ч при 37 С.
2.2 Эксперимент 2: Трансформация плазмидной ДНК бактерий
Трансформация – направленный перенос в клетку чужеродной генетической информации с помощью изолированной ДНК. Трансформация бывает естественной (например, у Baccilus subtilis) или искусственная.
Стадии:
Контакт с поверхностью; Проникновение ДНК в клетку; Соединение трансформирующей ДНК с соответствующим фрагментом хромосомы реципиента; Репликация новой информации.Способы трансформации:
- Химический; Замораживание-оттаивание; Электропорация – способ трансформации при помощи электрического поля с использованием электропоратора; Трансфекция – способ введения нуклеиновой кислоты в клетки невирусным методом; Баллистическая трансформация – введение генетического материала путем «обстреливания» растительной ткани мельчайшими частицами с нанесенными на х поверхность ДНК; Агробактериальная трансформация (у растений) – метод введения в клетки целевых генов при помоци бактерий; Трансформация протопластов – лишенных клеточной стенки растительных клеток.
Методы трансфекции:
- Ca-P трансфекция; Микроинъекция – введение нуклеиновой кислоты в клетку с помощью микроманипулятора и микропипетки; Водорастворимые катионные полимеры; Липосомная трансфекция – способ введения нуклеиновых кислот в клетку в составе липосом – сферических оболочек, состоящих из фосфолипидов. Баллистическая трансфекция Электропорация
1980г – Дж. Шелл, Марк Ван Монтегю – открытие агробактериальной трансформации. Agrobacterium tumefaceus вызывает так называемый «бактериальный рак» у растений. Имеют Ti - и Ri-плазмиды.
Плазмиды – автономно реплицирующиеся внехромосомные генетические элементы; двухцепочечные кольцевые замкнутые молекулы ДНК.
1946г – Дж. Ледерберг и Э. Тайтем – открытие конъюгации.
Конъюгация – однонаправленный перенос части генетического материала при непосредственном контакте клеток.
Донор выпускает половой пиль; Пиль соединяет Донора с Реципиентом; В мобильной плазмиде происходит однонитевой разрыв и одна цепь переходит в другую клетку; Обе клетки достраивают вторую цепь плазмиды и образуют половые пили.Плазмиды при проведении трансформации E. coli попадают только в очень небольшую часть (0,01–5%) клеток. Для отбора трансформированных клеток, несущих рекомбинантную ДНК, используют селективные маркеры, например, гены устойчивости к антибиотику. После осуществления процедуры трансформации бактерии высеивают на питательный агар, содержащий ампициллин. При этом выживают и образуют колонии только те клетки, в которые попала плазмида и синтезируется активная в-лактамаза. Эти клоны E. coli называют трансформантами. Другие векторы несут гены устойчивости к канамицину, тетрациклину. Таким образом, получившие рекомбинантную плазмиду, т. е. трансформированные или изменённые по сравнению с исходными клетки приобретают способность размножаться на питательных средах, содержащих антибиотики. В то время как на остальные клетки бактериальной культуры антибиотики оказывают бактериостатическое или бактерицидное действие. Разработанные позднее методы электротрансформации клеток обладают более высокой эффективностью по сравнению с «химической» или Ca2+ - трансформацией. Эти методы требуют специального оборудования для «электропробоя» мембран клеток – электропораторов.
Приготовление компетентных клеток E. coli
Приведённая процедура предполагает получение компетентных клеток E. coli путём их инкубации при низкой температуре в растворе, содержащем катионы Ca2+. Чтобы повысить уровень 50 трансформации помимо CaCl2 в раствор добавляют различные вещества. Приобретшие компетентность «кальциевые клетки» следует использовать сразу. Если по каким-то причинам это сделать невозможно, то 1–2 дня максимум они могут храниться в холодильнике при +4 С, пока полностью не утратят компетентность. Поэтому их лучше быстро заморозить в жидком азоте и хранить при –70 С. Все работы с компетентными клетками проводят на холоде, используя поддоны с мелкоколотым льдом или снегом.
Материалы и оборудование
Штамм E. coli XL1–Blue, диметилсульфоксид (ДМСО).
Растворы
- Среда SOB. 2% триптон; 0,5% дрожжевой экстракт; 10мМ NaCl; 2,5мМ KCl; 10мм MgCl2; 10мМ MgSO4.
- Буфер TB. 10мМ HEPES (N-2-гидроксиэтилпиперазин-N’-2-этансульфо-новая кислота, можно исп. PIPES или BES); 15мМ CaCl2; 55мМ MnCl2; 250мМ KCl.
Методика
1. Рассеять бактерии штрихом на чашку с селективной агаризованной средой SOB, выращивать при 37 С в течение ночи.
2. Несколько колоний E. coli (10–12) диаметром приблизительно 2–3 мм поместить в 40 мл среды SOB в колбе на 0,5–1 л. Растить при интенсивном встряхивании на качалке при скорости вращения 200–300 об/мин до оптической плотности OD600 = 0,6 при температуре 37 С. Обычно на это требуется 2–2,5 ч. Все дальнейшие манипуляции проводить на льду.
3. В 50 мл центрифужную пробирку поместить 30 мл культуры клеток, оставить при 0 C на 10–15 мин.
4. Центрифугировать 10 мин, 3000 об/мин, +4 C. Слить супернатант.
5. Центрифугировать 20 с, 3000 об/мин, +4 C. Удалить жидкость пипеткой.
6. Суспензировать в 10 мл буфера TB, оставить при 0 С на 10–15 мин.
7. Повторить п. 4 и п. 5.
8. Ресуспензировать осадок клеток в 2 мл буфера TB.
9. Добавить криопротектор ДМСО до 3,5% (70 мкл), оставить при 0 С на 10– 15 мин.
10.Повторить п. 4. Ресуспензировать клетки в остатках жидкости.
11.Разлить по 100 мкл в охлажденные 1,7 мл пробирки. Использовать «кальциевые клетки» сразу или заморозить в жидком азоте.
Примечание: При выращивании бактерий при температуре 37 С получается компетентность среднего уровня. При росте культуры E. coli при 18 С компетентность клеток получается выше, но время роста до нужной плотности удлиняется до нескольких суток. [1]
Трансформация E. coli
Для трансформации E. coli используют компетентные клетки и очищенную плазмидную ДНК или «лигазную смесь». ДНК добавляют к клеткам и выдерживают на льду некоторое время для сорбции ДНК на клеточной поверхности. Затем проводят кратковременный «тепловой шок» при 42 С, при котором ДНК проникает внутрь клеток E. coli, добавляют питательный бульон и рассеивают бактерии на агаризованной питательной среде, содержащей антибиотик. В результате экспрессии плазмидных генов устойчивости к антибиотикам трансформированные клетки приобретают способность расти на таких средах. На плотной питательной среде вырастают за 1–2 суток хорошо видимые колонии (≈ 104 –105 клеток), но только из тех клеток, которые получили плазмиду. Т. е. клоны-трансформанты. Не все трансформанты являются рекомбинантами, т. е. могут быть трансформированы исходным («пустым») вектором, если не применялись специальные процедуры для предотвращения такой возможности.
Методика
Посеять «ночную культуру» бактериальной питательной среде LB 1/20 «ночной культуры» инокулировать 7-10 мл свежей среды Охладить клетки во льду, центрифугировать Клеточный осадок перенести в чистый эппендорф, добавить 100мМ раствораВ ходе данных экспериментов мы последовательно выделили плазмидную ДНК, а затем трансформировали E. coli, использовав уже рекомбинантные плазмиды, кодирующие ген PFP. Спустя некоторое время на чашке Петри выросла одна заметная бактериальная колония диаметром около 4мм (приложение 10), которая при поглощении определенной длины волн начинала экспрессировать мягкий фиолетовый свет. Затем экспрессированный белок был также выделен нами из бактериальных клеток с использованием порошка солей алюминия для механического разрушения клеточных мембран (приложения 8, 9).
Выводы
В результате нашего исследования нам удалось установить природу и особенности строения органических флуоресцентных соединений, таких как зеленый и прочие флуоресцентные белки.
Также нами были рассмотрены способы использования биолюминесцентных ресурсов, такие как, например, флуоресцентная микроскопия сверхвысокого разрешения.
В будущем ученые делают ставку на то, что широкое использование всего спектра специфичных протеинов позволит им наблюдать за процессами жизнедеятельности организмов и вылечивать многие болезни, используя знания о механизмах их распространения. Помимо этого исследования возникновения самого явления биолюминесценции могут раскрыть больше подробностей о происхождении всех организмов в общем и в частности явлении «животного фотосинтеза».
В ходе экспериментов нами были сначала выделены плазмиды из бактериальных клеток, а затем выведена колония клеток, содержащих в себе рекомбинантные плазмиды и экспрессирующих флуоресцентные белки. Эти же белки, но уже выделенные из бактериальных клеток в ходе эксперимента, были рассмотрены нами в ультрафиолетовом спектре.
Заключение
Полученные данные могут быть использованы в различных отраслях биотехнологии и медицины. Например, для быстрого тестирования гнойных ран, так как пробы на чувствительность к антибиотикам дорогостоящи и занимают много времени, маркировки штаммов бактерий, таких как бактерия ботулизма, или плесневых грибов.
Зеленый флуоресцентный белок (GFP) является прекрасным маркером для визуализации живых клеток и организмов. Особенностью флуоресцентных белков является способность формировать флуорофорную группу автокаталитически, без привлечения внешних кофакторов и ферментов, т. е. они могут светиться внутри клетки сами по себе. Независимое созревание, в сочетании с низкой токсичностью и высокой стабильностью, позволяет использовать флуоресцентные белки в самых разных организмах, в которых исходно они не синтезировались.
Преимущество бактериофитохромов заключается в том, что их кофактор (например, биливердин) участвует в нормальном метаболизме гема млекопитающих. Визуализация различных процессов в этих организмах in vivo важна для таких задач как количественная и неинвазивная оценка роста и метастазирования опухоли, изучение экспрессии генов, отслеживание бактериальной инфекции и т. д. Вкратце, — можно наблюдать за этими процессами, поместив флуоресцентный белок под определенный промотор, который активируется только в конкретных условиях, или в опухолевые клетки; тогда регистрация сигнала от белка будет означать, что промотор активен, или что клетки трансформировались, или что определенный молекулярный путь запустился.
Визуализация глубоких тканей с помощью GFP-подобных белков, к сожалению, сильно затруднена из-за сильного поглощения света гемоглобином и меланином кожи. Оптимальный флуоресцентный белок для имиджинга in vivo должен обладать максимумами поглощения и флуоресценции в дальнекрасной-инфракрасной области, в которой поглощение света тканью минимально.
Список литературы
Молекулярная биология. Практическое руководство: Учеб. – Саратов: Издательство «Саратовский источник», 2013. – 84 с. ехнологии фотоактивируемых флуоресцентных белков - https://postnauka. ru/video/39997 G. H. Patterson, J. Lippincott-Schwartz. A Photoactivatable GFP for Selective Photolabeling of Proteins and Cells || Science, 2002 H. Yokoe, T. Meyer. Spatial Dynamics of GFP-tagged Proteins Investigated by Local Fluorescence Enhacement || Nature biotechnology, 1996 J. Haseloff, B. Amos. GFP in Plants || Elsevier Current Trends, 1995 Green Fluorescent Protein Site - http://www. conncoll. edu/ccacad/zimmer/GFP-ww/GFP-1.htm N. G. Gurskaya, A. F. Fradkov, A. Terskikh GFP‐like chromoproteins as a source of far‐red fluorescent proteins || FEBS …, 2001 - Wiley Online Library Protein Data Bank - https://pdb101.rcsb. org/motm/42Приложение 1. Структура флуорофора
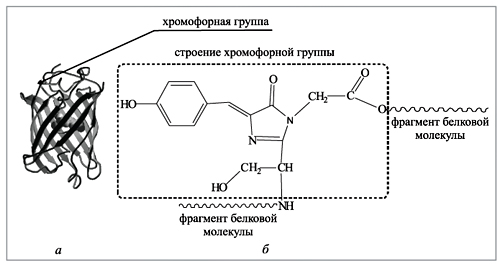
а – молекула GFP, полипептидные цепи условно показаны в виде лент и жгутов; хромофорная группа, обеспечивающая флуоресценцию, показана в виде шаровой структуры; б – строение хромофорной группы
Приложение 2. Синтез хроматофорной группы из трех аминокислот
Приложение 3. Структура GFP
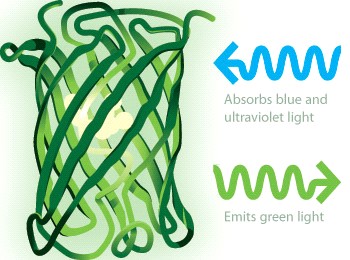
Приложение 4. Флуоресцирующие черви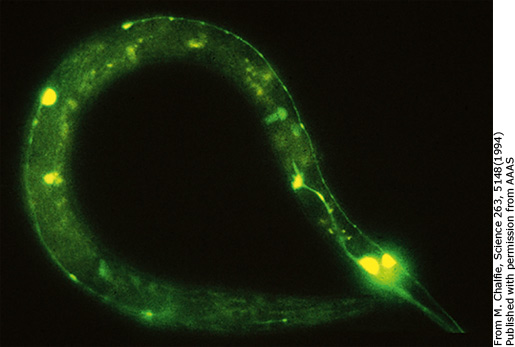
Приложение 5. Использование GFP как метки в молекулярной биологии
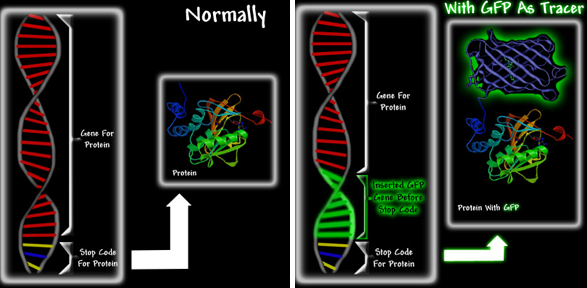
Приложение 6. Донорно-акцепторное изменение GFP

Приложение 7. Оборудование для проведения экспериментов: центрифуга и микропипетки
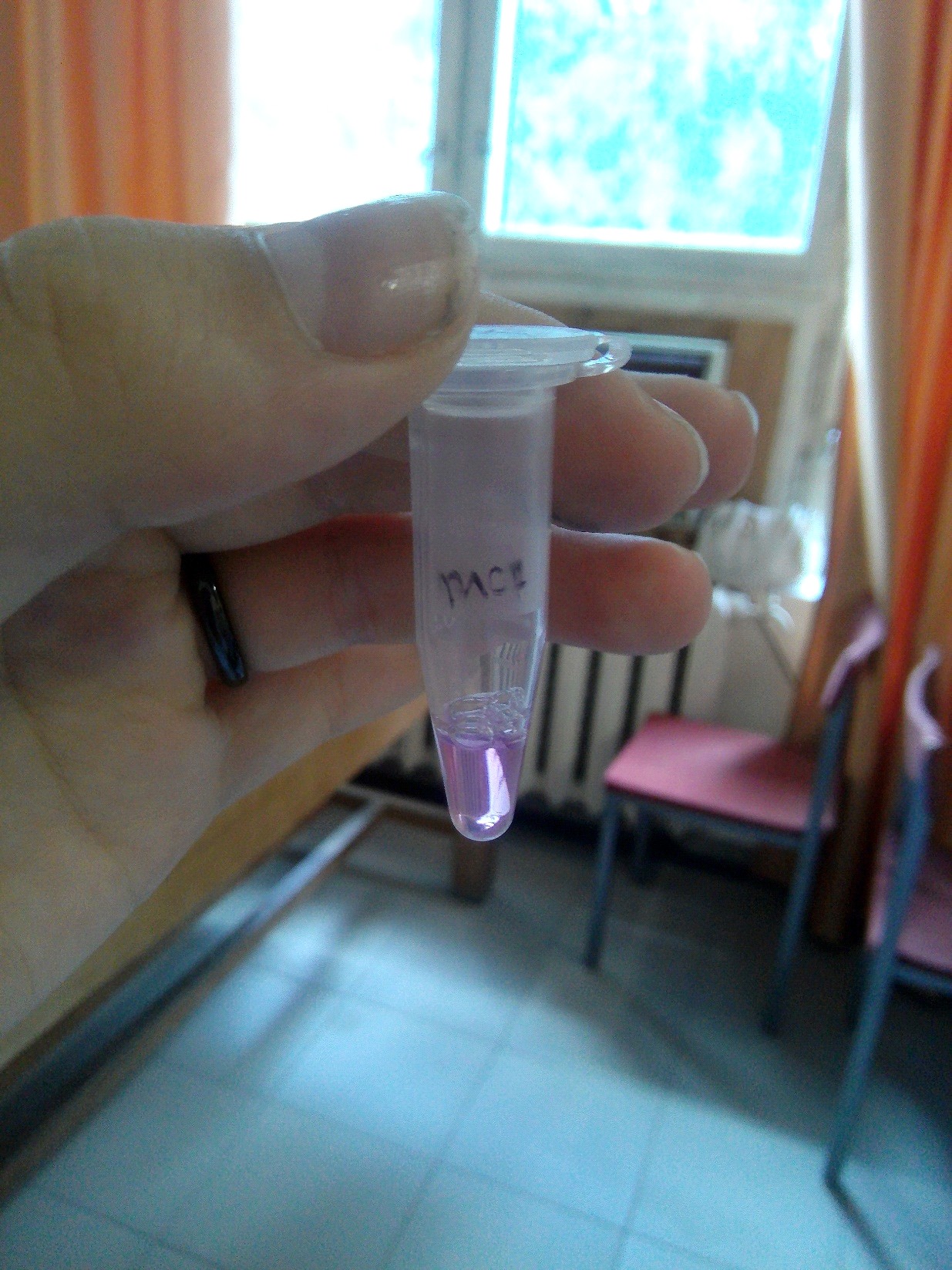
Приложение 8. Эппендорф с раствором PFP

Приложение 9. Эппендорфы с белками-гомологами GFP под ультрафиолетом
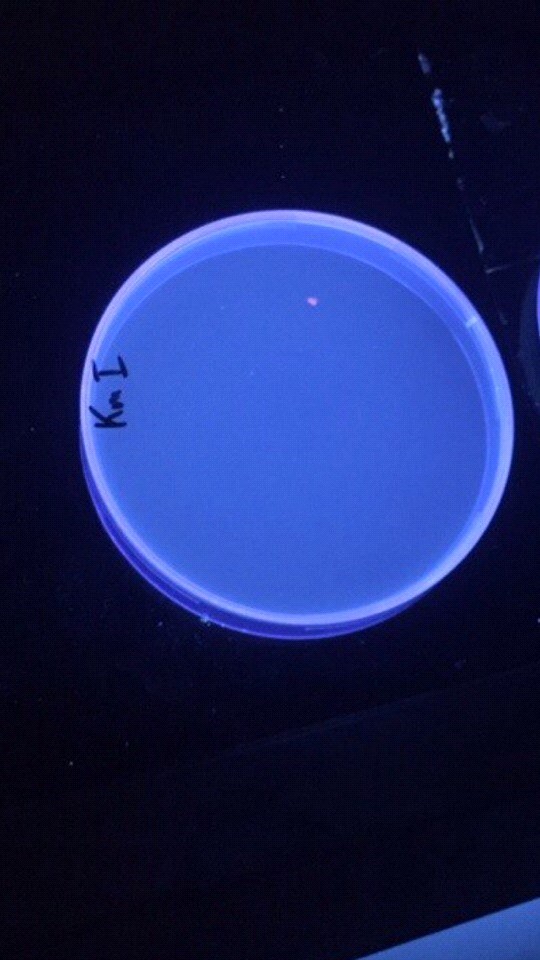
Приложение 10. Чашка Петри с выращенной культурой бактерий E. coli, экспрессирующих фиолетовый флуоресцентный белок
