

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
УДК 631.527:633:1 На правах рукописи
БУЛАТОВА КУЛЬПАШ МАНСУРОВНА
Биологические основы селекции
зерновых колосовых культур
06.01.05 – Селекция и семеноводство
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
Республика Казахстан
Алматы, 2009
Работа выполнена в ТОО «Казахский научно-исследовательский институт земледелия и растениеводства
» АО «КазАгроИнновация»
Научные консультанты: доктор биологических наук, профессор, академик НАН РК, УААН и РАСХН ,
доктор биологических наук, профессор
Официальные оппоненты: доктор биологических наук,
профессор, академик НАН РК, ,
доктор биологических наук, профессор ,
доктор биологических наук, профессор, академик РАЕН,
Ведущая организация: Казахский национальный аграрный университет
Защита состоится 5 июня 2009 года в 14.00 часов на заседании диссертационного совета ОД 55.05.01 при Казахском НИИ земледелия и растениеводства
Республика Казахстан, Алматинская область, Карасайский район, п. Алмалыбак, .
С диссертацией можно ознакомиться в библиотеке Казахского НИИ земледелия и растениеводства.
Автореферат разослан ____ ____________________ 2009 г.
Ученый секретарь
диссертационного совета,
доктор с-х. наук
ВВЕДЕНИЕ
Актуальность проблемы Пшеница и ячмень являются основными зерновыми колосовыми культурами, возделываемыми в Казахстане. Посевные площади этих культур составляют в Республике более 15 млн. га. Одним из главных факторов увеличения валового сбора зерна является селекционный путь - создание высокоурожайных сортов с комплексом ценных хозяйственно-биологических признаков и свойств.
Успех в решении этой важнейшей для сельскохозяйственной отрасли Республики задачи зависит от уровня генетического разнообразия исходных коллекций, гибридных популяций, методологической базы отбора желаемых генотипов в ходе селекционного процесса.
Значимость биологических методов анализа для решения проблем генофонда, селекции и семеноводства сельскохозяйственных культур бесспорна, однако их практическое применение все еще не на должном уровне. Разработка, совершенствование и внедрение надежных и эффективных методов биологического анализа в изучение генетических ресурсов, процесс отбора ценных форм, сохранение биологических свойств сорта в ходе семеноводства являются актуальными задачами для отечественной селекции и необходимыми условиями повышения продовольственной безопасности страны.
В национальном генофонде Республики сосредоточено более 30 тыс. образцов пшеницы, 3 тысяч коллекционных номеров ячменя, представленных такими категориями, как: коммерческие и местные сорта, дикие сородичи, признаковые коллекции. Генетическое разнообразие коллекционного материала не изучено, следовательно, не использовано в достаточной мере. В то же время, образцы с редкими и новыми аллелями признаков могут дать начало селекции высокоурожайных, устойчивых к неблагоприятным условиям среды, ценных по другим хозяйственно-ценным показателям сортов. Выявление таких форм и их использование в селекции сопряжено с изучением большого числа коллекционных образцов, целенаправленного подбора компонентов для скрещиваний, поэтапной оценки селекционных линий. Успешная реализация этих задач невозможна без использования достижений генетики, молекулярной биологии, физиологии, и других наук в биологическом анализе растений на этапах селекционного процесса зерновых колосовых культур. Особую значимость в их числе приобретают методы, разработанные на основе специфичности состава запасных белков, ферментов, накоплении определенных метаболитов на разных стадиях, условиях развития генотипа и сопряженности их характеристик с проявлением хозяйственно-ценных признаков, что актуально для селекции сельскохозяйственных культур.
Работа была выполнена в рамках государственных программ: гг. «Разработать новые и усовершенствовать существующие схемы, методы селекции и на их основе создать новые высокопродуктивные, устойчивые к болезням, вредителям, неблагоприятным факторам среды и с высоким качеством продукции сорта и гибриды пшеницы, ячменя, овса, кукурузы, сорго, сои, гороха с разработкой экологических основ промышленного семеноводства». № гос. регистрациии 0194РК00028; гг. «Создать новые с использованием методов генетики, биотехнологии и других биологических дисциплин устойчивые к стрессовым факторам среды, высокопродуктивные сорта и гибриды зерновых, зернобобовых, сорговых культур». № гос. регистрации 0196РК00292; гг. «Сбор, интродукция, комплексное изучение и выявление доноров с желательными хозяйственно-ценными признаками, организация, поддержание существующих коллекций зерновых, зернобобовых, крупяных, технических и лекарственных культур на Юге и Юго-Востоке Казахстана», № гос. регистрации 0101РК00226;
гг. «Разработка биохимических подходов для дифференциации генотипов пшеницы по устойчивости к желтой ржавчине», № гос. регистрации 0106РК00730.
Цель работы – Разработать и усовершенствовать методические основы биологического анализа растений в селекции пшеницы и ячменя.
Для достижения цели необходимо было решить следующие задачи:
-определить вариабельность состава запасных белков пшеницы, установить генетический контроль новых субъединиц белков, усовершенствовать систему регистрации коллекционных и селекционных образцов, мутантных линий, сортов по генетико-биохимическим параметрам, провести регистрацию генетических ресурсов и выявить ценные для селекции генотипы;
-разработать систему регистрации и идентификации линий, биотипов и сортов, оценить разнообразие генетических ресурсов ячменя по полиморфности белок-кодирующих локусов;
-выявить биологические особенности жаро-засухоустойчивых и скороспелых форм ячменя, разработать на этой основе методы отбора желаемых генотипов в коллекционных и селекционных питомниках;
- выделить коллекционные образцы – источники и доноры хозяйственно-ценных признаков для селекции пшеницы и ячменя; на основе разработанных методов и приемов биологического анализа, отобрать перспективные селекционные линии.
Научная новизна Впервые для идентификации коллекционного и селекционного материала пшеницы и ячменя предлагается комплекс биологических тестов, включающих:
- уточненную номенклатуру высокомолекулярных субъединиц глютенина;
- сорта – дифференциаторы для идентификации, отбора и паспортизации коллекционных и селекционных линий пшеницы;
- совершенствованную систему регистрации образцов ячменя по гордеинам с генетико-биохимической характеристикой их компонентов.
Проведена генетико-биохимическая идентификация по белковым формулам 1831 образца генофонда зерновых колосовых культур, установлено генетическое разнообразие коллекций, выделены источники редких и ценных для селекции генов.
Предложен комплекс биологических тестов для отбора скороспелых и засухоустойчивых форм ячменя в поэтапной оценке в селекционных питомниках.
Предложена схема биологических тестов в селекции пшеницы и ячменя по основным хозяйственно-ценным признакам.
Практическая ценность работы
Проведена идентификация и регистрация генофонда пшеницы и ячменя по блокам компонентов запасных белков зерна, результаты опубликованы в 2-ух каталогах.
На основе спектров запасных белков ведется паспортизация сортов пшеницы и ячменя, передаваемых на Государственное сортоиспытание;
С участием соискателя переданы на Государственное сортоиспытание 2 сорта озимой мягкой и 1 сорт яровой твердой пшеницы (от 5 до 10% участия в авторстве);
Созданы константные линии озимой пшеницы с эффективными генами устойчивости к желтой ржавчине, 3 из которых находятся в питомнике конкурсного сортоиспытания;
Для селекции на засухоустойчивость и скороспелость создано 233 гибридных и 34 перспективных по комплексу ценных признаков и свойств линий гордеиновых биотипов ячменя, часть из которых испытывается в питомниках конкурсного сортоиспытания. Линия 61/7 показала значительное превышение по урожайности над стандартным сортом.
Положения, выносимые на защиту:
-усовершенствованная система регистрации коллекционных и селекционных образцов пшеницы и ячменя по белковым формулам;
-идентифицированные коллекции генофонда ячменя и пшеницы, их генетическое разнообразие, ценные источники хозяйственно-биологических признаков;
-комплекс биологических критериев в дифференциации и отборе скороспелых и засухоустойчивых форм ячменя;
-перспективные линии и сорта пшеницы и ячменя, как практический результат биологического анализа генофонда и использования ценных источников в селекции по основным хозяйственно-ценным признакам.
Апробация работы Результаты исследований были представлены и доложены на 27 научно-практических конференциях, совещаниях и симпозиумах, в т. ч. на межд. сов. «Генбанк растений и его использование в селекции», (Алматы, 1995), межд. науч. пр. конф. «Аграрная наука на рубеже веков» (Акмола, 1997), 2-ой межд. конф. «Проблемы экологии АПК и охрана окружающей среды» (Алматы,1998), межд. науч. конф. «Природные соединения – регуляторы метаболизма и адаптации растений» (Алматы, 1999), межд. науч. конф. «Физиолого-биохимические и генетические основы устойчивости и продук-тивности раст.» ( Алматы, 1999 ), мат. межд. научно-теоретической конф. «Стратегия земледелия и растениеводства на пороге ХYI века.» ( Алма-ты,1999), на 8-м Международном симпозиуме по ячменю (Adelaide, South Australia, 2000), на 4-ой межд. науч. конф. «Проблемы экологии АПК и охрана окружающей среды» (Щучинск, 2002), на международном симпозиуме “Biotechnology Approaches for Exploitation and Preservation of Plant Resources” (Ялта, 2002), межд. науч. конф. «Современное состояние проблем и достижений в области генетики и селекции» (Алматы, 2003), первой центрально-азиатской конференции по пшенице, (Алматы, 2003), 6-ой межд. науч. практ. конф. . «Научное обеспечение устойчивого развития АПК Казахстана, Кыргызстана, Монголии, России, Таджикистана и Узбекистана», (Павлодар, 2003), межд. научн. конф. «Стратегия научного обеспечения АПК РК в отраслях земледелия, растениеводства и садоводства. Реальность и перспективы», (Алматы, 2004), межд. научн. конф. «Достижения аграрной науки в области земледелия, селекции и растениеводства», (Алмалыбак, 2004), межд. конф. «Развитие ключевых направлений сельскохозяйственной науки в Казахстане: селекция, биотехнология, генетические ресурсы», (Астана, 2004), 3-ем Московском Международном конгрессе «Биотехнология: состоя-ние и перспективы развития» (Москва, 2005), Республиканской научно-теоретической конференции «Сейфуллинские чтения – 3», посвященной 50-летию основания КазГАТУ, Астана, 2007, 10-ой международной конференции по научному обеспечению азиатских территорий (Улан-Батор, 2007), на XI межд. научно-практ. конф. «Развитие АПК азиатских территорий»- (Новосибирск.-2008), международной конференции «Современные методы использования генетических ресурсов в селекции ячменя и овса» (С. Петербург, 2009).
Публикации: по материалам диссертации опубликовано 76 научных работ (в списке приведено 46), в том числе 2 каталога, подана 1 заявка на получение патента РК (№ гос. регистрации 2007/0593.1).
Структура диссертации диссертационная работа изложена на 270 страницах печатного текста, состоит из введения, основной части, включающей обзор литературы, объекты, методы и условия проведения исследований, результаты исследований, а также заключения, списка использованных источников, включающего 533 наименования, содержит 95 таблиц и 39 рисунков, 4 приложения.
Автор выражает особую признательность научным консультантам: академику и д-ру биол. наук за ценные советы и предложения при подготовке диссертации.
ОСНОВНАЯ ЧАСТЬ
1 Обзор литературы В обзоре дана оценка современного состояния по использованию методов биологического анализа растений в селекции и семеноводстве сельскохозяйственных культур по проблемам: генетические марке-ры в изучении и регистрации генофонда; молекулярные маркеры в решении проблем генофонда, селекции и семеноводства; ферменты как системы белковых марке-ров; запасные белки семян как молекулярные маркеры; методы и приемы диагнос-тики засухоустойчивости растений; биохимические особенности засухоус-тойчивых форм растений; скороспелость как механизм адаптации растений к засухе.
2 Объекты, методы и условия проведения исследований
2.1 Условия проведения исследований Аналитические исследования проводились в лаборатории КазНИИЗиР, полевые исследования велись в агроэкологических зонах Алматинской (полевые стационары КазНИИЗиР), Западно-Казахстанской (полевые стационары Актюбинская СОС), Акмолинской (полевые стационары КазНИИЗХ им. А.И. Бараева), Павлодарской (полевые стационары Павлодарского НИИСХ) областей. Агротехника на селекционных посевах была общепринятой для соответствующих зон.
2.2 Объекты исследований: 811 образцов генофонда Triticum aestivum L., включающих сорта озимого и ярового типа развития, коллекционные номера зарубежной селекции, линии международных питомников; номера селекционных питомников яровой мягкой пшеницы: СП-2 и КП – 307 и 299 номеров, соответственно; регенерантные линии дигаплоида АДГ 147 – производные сорта Саратовская 52, изогенные линии с генами устойчивости к желтой ржавчине; 264 коллекционных образца Triticum durum; 752 образца генофонда ячменя, гибридные линии ячменя F3 – F 6 поколений.
2.3 Методы исследований: Полевые оценки и структурный анализ растений проводили по стандартным методикам, применяемым для характеристики селекционного материала. Гибридизацию проводили «твелл» методом (Borlaug, в модификации Уразалиева, Шегебаева, 1981). Глютенины в составе суммарных белков эндосперма экстрагировали 0,0625 М трис - НСl буфером, рН 6,8, содержащим 5% меркаптоэтанола, 3% додецилсульфата натрия, 10% глицерина, 0,1% красителя бромфенолового синего. Система разделения соответствовала модифицированному методу Лэммли,1970 (, 1985). Компонентный состав проламинов определяли путем электрофореза экстрагированной 70% этанолом белковой фракции в полиакриламидном геле в кислой системе по прописи и др. (. 1989). Количественное соотно-шение компонентов в белковом спектре определяли путем сканирования электро-фореграмм с последующим расчетом площадей пиков. Засухоустойчивость стан-дартных сортов ячменя определяли по числу проросших семян после прогревания и проращивания в растворе сахарозы (Методические указания ВИР под. ред. 1982), по уровню свободного пролина в проростках материал оценивался методом Бейтса с сотр. (Bates et. al., 1973). Общая активность пероксидазы замерялась на спектрофотометре СФ-26 и СПЕКОЛ-11 по скорости реакции окисления бензидина (по Бояркину). Перевод активности в нанокаталы проводили по и др.,1977. Электрофоретическое разделение пероксидаз осуществляли в трис-глициновом буфере при рН 8,3. Изозимы пероксидазы выявляли бензидиновым реагентом в ацентатном буфере при рН 4,7 с последующим переносом в 0,1 % водный раствор перекиси водорода (, 1975). Относительную активность индивидуальных изоформ определяли хронометрированием (Liu E. M. 1973). Оценка проростков озимой пшеницы на устойчивость к расам желтой ржавчины в условиях теплицы по методике Gassner, Straib (1929), полевая устойчивость растений к видам ржавчины по шкале Peterson и др. (1948) определялась в лаборатории «иммунитета растений» НИИПББ НЦБ МОН РК. Математическая обработка данных, в т. ч. и кластеризация методом UPGMA осуществлялись при помощи пакета программ Statistica V 5.0. Оценку генетического разнообразия коллекций определяли согласно способа Nei в модификации Hintum & Elings, 1991, цитировано по Morgunov A. I. et. al. 1993.
В автореферате имеются следующие сокращения и обозначения: Glu A1, - B1, - D1 локусы – глютенинкодирующие локусы; ВМСГ –высокомолекулярные субъединицы глютенина. НМГ–низкомолекулярный глютенин; Hrd А,-B,–F-гордеинкодирующие локусы; ДДСNaПААГ–полиакриламидный гель с додецил-сульфатом натрия; ОЭП – относительная электрофоретическая подвижность; СИММИТ - Международный центр улучшения пшеницы и кукурузы; ИКАРДА – Международный Центр по сельскохозяйственным исследованиям в засушливых ре-гионах; КАСИП–Казахстанско-Сибирский питомник; WWERYT – Восточно – Европейский питомник сортоиспытания озимой мягкой пшеницы; КСИ - питомник конкурсного сортоиспытания; ГСИ – государственное сортоиспытание.
3 Результаты исследований
3.1 Совершенствование способов идентификации и отбора генотипов пшеницы и ячменя по биохимическим тестам Возросший в последнее время интерес к субъединичному составу глютенинов пшеницы связан с функциональной значимостью конкретных белков в детерминации хлебопекарных свойств. На этой основе рекомендованы градации отдельных высокомолекулярных субъединиц глютенина в соответствии с их вкладом в так называемую оценку по глютенину - Glutenin quality score (Payne et. al., 1987). В то же время, в ходе исследований выявляются редкие и новые варианты субъединиц глютенина пшеницы, изучение и оценка селекционной ценности которых может в значительной степени повысить эффективность маркерной селекции. Кроме того, отмечается некоторое несоот-ветствие прогнозной оценки качества по глютенинам с результатами традиционных методов оценки технологических показателей. Причиной этого может являться различие в аллельном состоянии глютенинкодирующих локусов, не выявляемое при фракционировании продуктов их биосинтеза при определенных условиях. Уточнение генетического контроля и идентификация новых и редких белковых субъединиц, выявляемых в ходе изучения коллекционного и селекционного материала необходимы как для достоверной оценки образцов, так и для расширения генетической основы создаваемых сортов путем изучения ценности генотипов со специфическими вариантами аллелей и включения их в селекционный процесс.
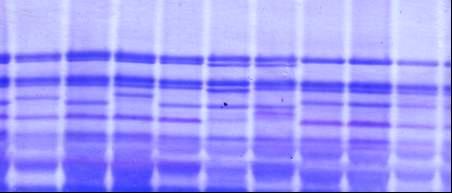
3.1.1 Новые и редкие аллели локусов, контролирующих биосинтез высокомолекулярных субъединиц глютенина пшеницы В электрофоретическом спектре запасных белков мягкой пшеницы (геном АА ВВ DD), при фрак-ционировании белковых проб стандартным методом Laemmli, как правило, в зоне ВМСГ, контролируемых локусом Glu B1, проявляются 2 компонента. Разделение белков в модифицированных нами условиях выявляет еще один компонент (рисунок 1, дорожка №2, отмечен белой точкой), которого нет в спектре ВМСГ твердой пшеницы (дорожка №1). Аналогичный вариант субъединицы (а) характерен для многих сортов озимой пшеницы Казахстанской селекции, в том числе и Богарной 56 (дорожка №8), для известного сорта Безостая 1. Дополнительный компонент (b) проявляется и в спектре яровой мягкой пшеницы Саратовская 52, Саратовская 29 и многих других сортов мягкой пшеницы селекции НИИСХ Юго-Востока РФ (дорожка 4, черная точка), с той лишь разницей, что он имеет меньшую электро-форетическую подвижность. Отсутствие сопутствующей субъединицы в спектре ВМСГ твёрдой пшеницы, не имеющей D геном, ее наличие у дигаплоидной линии с мутацией, приведшей к исчезновению Вх7 субъединицы, обнаружение в спектре ВМСГ дикого сородича Aegilops cylindrica (дорожка №7, показана стрелкой) аналогичной субъединицы свидетельствуют о том, что ее контроль осущест-вляется D геномом. Выявлено 4 варианта сопутствующих субъединиц, контролируемых 1D хромосомой: a, b, c, d. (носители Безостая 1, Саратовская 29, Бабило, Сурхак Ишкашимский, соответственно). Для более точной оценки генетического контроля сопутствующих субъединиц были использованы нуллисомные линии сорта Чайниз Спринг (nullisome 1D tetrasome 1B – дорожки № 3,4 на рисунке 2, nullisome 1D tetrasome 1A –дорожки №5,6, nullisome 1B tetrasome 1D – дорожка №7). Электрофорез запасных белков в модифицированных условиях показал, что биосинтез высокомолекулярной субъединицы, сопутствующей субъединице глютенина 7 в центральной части зоны ВМСГ (показано стрелкой) контролируется 1D хромосомой. Анализ частоты встречаемости субъединиц 7 в сочетании с обнаруженной «а» субъединицей в составе глютенинов мягкой пшеницы из разных селекционных центров показал, что доля образцов, имеющих в составе высокомолекулярных глютенинов субъединицы 7 и «а» наиболее значительна в наборе районированных и проходящих испытание сортов Казахстана (90,5%), среди сортов, созданных в селекционно-генетическом институте – национальном центре семеноведения и сортоизучения УААН (95,7%) и Мироновском институте пшеницы им (100%), что является косвенным подтверждением ее значимости в детерминации хлебопекарных свойств, поскольку сорта Казахстанской селекции и этих селекционных учреждений, как правило, высококачественны. В сорте озимой мягкой пшеницы Богарная 56 выявлены биотипы, идентичные по спектру проламинов, но различающиеся по составу ВМСГ: у одного из них (типичного) субъединица 7 сопряжена с сопутствующим вариантом а, а у другого - с вариантом субъединицы b. По данным таблицы 1 , видно, что типичный биотип, имеющий в своем составе дополнительную субъединицу а, характеризуется более высокими показателями качества в сравнении с другим биотипом. Полученные данные свидетельствуют о том, что при полном следовании условиям фракционирования высокомолекулярных глютенинов в ДДС-Na ПААГе по Laemmli (1970) микрогетерогенность субъединиц нивелируется и в спектре насчитывается ограниченное число компонентов. Модификация метода, предложенная нами (Булатова, 1985), позволяет дифференцировать варианты субъединицы 7, контролируемой локусом Glu B1 и выявить в зоне электрофоретического спектра ВМСГ, традиционно считающейся месторасположением продуктов экспрессии локуса Glu B1, 4 варианта специфичных субъединиц, кодируемых D геномом. Присутствие определенных вариантов этих субъединиц сопряжено с показателями качества мягкой пшеницы, в связи с чем, их изучение и дискриминация с помощью модифицированного метода необходимы в оценке материала генофонда и в селекции высококачественных образцов. Генотипы пшеницы с сочетанием субъединиц 7 - «а» и 7 – «b» встречаются с достаточной частотой, тогда как образцов с вариантами субъединицы 7, сопряженной с «c» и «d» в генофонде практически нет, за исключением староместных сортов Западного Памира: Бабило и Сурхак Ишкашимский. С целью выявления селекционной значимости специфичных аллелей с участием этих сортов были получены, отобраны по урожайности и адаптивности к условиям Юго Востока Казахстана и переданы селекционерам 45 линий F3-F4 поколений. У значительной части сортов яровой пшеницы селекции НИИСХ Юго-Востока по результатам фракциони-рования в 10 %-ном полиакриламидном геле по прописи Laemmli (1970) субъединичный состав глютенинов был определен как 2*; 7+9; 2+12; (Morgunov A. I. et. al., 1990, и др. 1991). Оценка качества по глю-тенину для таких сортов в соответствии с ранжированием Payne и др. (1987)
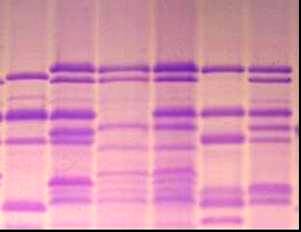


9 10
1- Наурыз 2; 2- Стекловидная 24; 3- дигаплоид Саратовской 52 с null мутацией в локусе Glu B1; 4-дигаплоид Саратовская 52 (контроль); 5 - Бабило; 6 - Ишкашимский Сурхак, 7,9,10 - образцы Aegilops cylindrica, Алматинская обл.; 8 - Богарная 56 (обозначения даны в тексте).
Рисунок 1 – Спектры высокомолекулярных глютенинов Tr. Aestivum L., T. durum Desf. и Aegilops cylindrica.
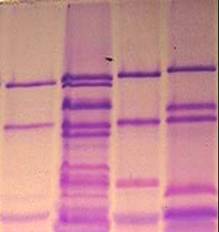
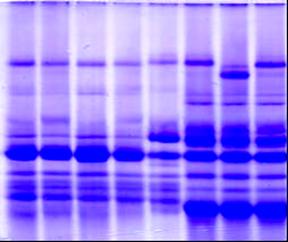
|

1,2 – Чайниз Спринг; 3,4 – нуллисомик по 1D и тетрасомик по 1 B хромосомам; 5,6 - нуллисомик по 1D и тетрасомик по 1 А хромосомам; 7 - нуллисомик по 1B и тетрасомик по 1D хромосомам.
Рисунок 2 – Электрофоретические спектры глютенинов у нулли-тетрасомных линий сорта Чайниз Спринг.
равна 7, в то же время эти сорта известны как наиболее высококачественные среди яровых пшениц. В связи с этим, ряд авторов придерживается мнения, что показатель качества по оценке ВМСГ не всегда совпадает с характеристикой хлебопекарных показателей сортов яровой пшеницы, полученных классическими методами ( Д и др. 1991, 2001). Отдельные случаи несоответствия хлебопекарных показателей качества муки бальной оценке по субъединицам глютенина лежат за пределами состава ВМСГ (Obukhova L. et. al., 2003). Причиной несоответствия реальных качественных показателей с бальной оценкой по составу ВМСГ может являться и различие в аллельном состоянии глютенинкодирующих локусов, не выявляемое по продуктам их биосинтеза при определенных условиях электрофореза.
Таблица 1 – Показатели качества глютениновых биотипов сорта озимой мягкой пшеницы Богарная 56 с сочетанием 7 - «а» и 7 – «b» субъединиц глютенина.
Биотип с ВМСГ: | Показа-тель седимента-ции, мл | Содержа-ние сырой клейкови-ны, % | Качество клейко-вины, балл | Число падения, сек. | Содержа-ние белка, % | Показа-тель ИДК |
2*;7+а+9;5+10 | 54±1,5 | 44,6±1,18 | 4,6±0,09 | 424±17,6 | 15,5±0,36 | 95,4±2,2 |
2*;7+b+9;5+10 | 48,4±1,4 | 47,8±3,2 | 3,4±0,2 | 409±21,3 | 14,7±0,36 | 104,1±3,8 |
НСР 0,95 | 4,31 | 7,17 | 0,46 | 58,05 | 1,07 | 9,23 |
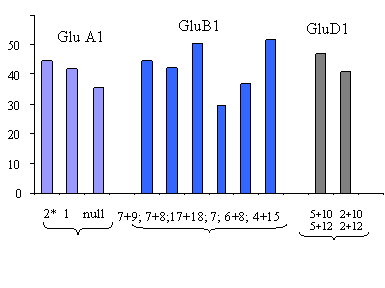
Анализ состава ВМСГ сорта Саратовская 29 и ряда других сортов (таблица 2), проведенный в модифицированных условиях, показал, что локусом Glu D 1 детерминируется вариант ВМСГ, в котором “х” субъединица не совпадает по подвижности с положением субъединицы 2: она располагается на уровне ВМСГ 5, однако отличается от нее по интенсивности проявления. Следовательно, она не является продуктом аллели «d» и контролируется новой, ранее не описанной аллелью локуса Glu 1D. Состав ВМСГ этих сортов в связи с полученными данными регистрируется как 2*; 7+9; 5** +12, а не 2*; 7+9; 2 +12, как опубликовано указанными в таблице 2 авторами. Ряд исследователей предполагает, что линии с субъединицей у-типа 12 характеризуются лучшим качеством, нежели таковые с субъединицей 10 (Rodriguez-Quijano M.,et. al., 2000). Преимущество субъединицы х-типа 5 по вкладу в качество зерна доказано давно. По-видимому, сочетание таких ценных показателей в одном генотипе и обеспечивает высокое качество саратовских сортов и созданных с их участием форм.
Еще один редкий аллельный вариант глютенинкодирующего локуса Glu B1, был выявлен в ходе изучения генетических изменений у регенерантных линий мягкой пшеницы по локусам, контролирующим биосинтез запасных белков зерна – глиадинов и глютенинов. В спектре высокомолекулярных глютенинов линии АД54, в сравнении с исходной линией выявлено отсутствие одной из высокомолекулярных субъединиц, классифицируемой по каталогу Payne P., Lawrence G. (1983) как х 7 ( рисунок 3), контролируемой локусом Glu B1. Мутантная линия с нулевой аллелью и ее исходная форма были изучены по ряду признаков урожайности и качества. Сравнение элементов урожайности показало превышение линии АД-54 над контрольным растением по ряду признаков урожайности, таких как: масса зерен с главного колоса и масса 1000 зерен (на 18,7% и 10,3%, соответственно). По ряду технологических показателей регенератная и исходная линия не отличались, однако по уровню седиментации АД-54 уступала исходной форме (29,5 мл. у регенеранта и 35,0 мл. у исходной формы). Результаты свидетельствуют, что отсутствие одной из ВМСГ, контролируемых локусом Glu B1 негативно отражается на показателях качества зерна и муки.
Таблица 2- Состав ВМСГ сортов яровой мягкой пшеницы по прописи Laemmli (1970) и в модифицированных условиях.
Сорт | Пропись Laemmli | Качес-тво (балл) | Модифицированные условия | ||||
Состав субъединиц по локусам | |||||||
Glu A1 | Glu B1 | Glu D1 | Glu A1 | Glu B1 | Glu D1 | ||
Казахстанская раннеспелая | 2* | 7+9 | 2+123 | 7 | 2* | 7+9 | 5**+12 |
Целинная 3с | 2* | 7+9 | 2+123 | 7 | 2* | 7+9 | 5**+12/5+10 |
Целинная 92 | 2* | 7+9 | 2+123 | 7 | 2* | 7+9 | 5**+12 |
Целинная 98 | 2* | 7+9 | 2+123 | 7 | 2* | 7+9 | 5**+12 |
Целинная Юбилейная | 2* | 7+9 | 2+123 | 7 | 2* | 7+9 | 5**+12 |
Акмола 2 | 2* | 7+9 | 2+123 | 7 | 2* | 7+9 | 5**+12 |
Омская 9 | 2* | 7+9 | 2+12¹ | 7 | 2* | 7+9 | 5**+12 |
Омская 18 | 2* | 7+9 | 2+12¹ | 7 | 2* | 7+9 | 5**+12 |
Омская 28 | 2* | 7+9 | 2+122 | 7 | 2* | 7+9 | 5**+12 |
Саратовская 29 | 2* | 7+9 | 2+12¹ | 7 | 2* | 7+9 | 5**+12 |
Саратовская 42 | 2* | 7+9 | 2+12¹ | 7 | 2* | 7+9 | 5**+12 |
Саратовская 55 | 2* | 7+9 | 2+12¹ | 7 | 2* | 7+9 | 5**+12 |
Примечание: ¹- Д и др. 1991; ²- , , 1999; ³ - 2001. |
|
5 7 1 9 12 2 |

 А В
А В1-ВМСГ; 2-НМСГ+глиадины; А – АД 54; В – АД 52
Рисунок 3 – Гаметоклональная изменчивость пшеницы по спектру высокомолекулярных субъединиц глютенина
Аналогичное ухудшение качественных показателей зерна и муки наблюдается у сортов пшеницы с «нуль» мутацией по локусу Glu A1 (Burnouf et. al., 1980, Payne et. al., 1987). Выявленную сопряженность между нуль аллелью глютенинкодирующих локусов и низкими технологическими показателями зерна и муки следует учитывать при браковке селекционного материала. Гибридные популяции, полученные с участием носителя редкой аллели локуса Glu B1, могут использоваться в разработке QTL продуктивности и качества.
3.1.2 Стандартные образцы мягкой и твердой пшеницы для определения состава ВМСГ сортов, селекционных линий, образцов генофонда В результате многолетних исследований и анализа образцов мягкой и твердой пшеницы были отобраны мономорфные образцы, носители различных вариантов аллелей глюте-нинкодирующих локусов, необходимые для идентификации селекционного и кол-лекционного материала по составу ВМСГ. В таблицах 3 и 4 приведен состав глюте-нинов сортов дифференциаторов, используемых для идентификации образцов пшеницы.
Таблица 3 - Состав ВМСГ сортов - дифференциаторов твердой пшеницы
№ п/п | Наименование сорта | Glu 1 локусы | ||
Glu A1 | Glu В1 | Glu В3 | ||
1. | Наурыз 1 | 0 | 7+8 | 1 |
2. | Наурыз 2 | 1 | 7+8 | 2 |
3. | Президент | 0 | 6+8 | 1 |
4. | Ватан | 0 | 20 | 2 |
5. | Каргала 3 | 0 | 13+16 | 2 |
6. | Гордеиформе 254 | 0 | 14+15 | 1 |
7. | Харьковская 17 | 2* | 7+8 | 2 |
Таблица 4 - Состав ВМСГ сортов - дифференциаторов мягкой пшеницы
№ п/п | Наименование | Glu 1 локусы | ||
Glu A1 | Glu B1 | Glu D1 | ||
1. | Безостая 1 | 2* | 7+9 | 5+10 |
2. | Богарная 56 (типичный биотип) | 2* | 7+а +9 | 5+10 |
3. | Богарная 56 ( биотип) | 2* | 7+b +9 | 5+10 |
4. | Саратовская 29 | 2* | 7+9 | 5+12 |
5. | Стекловидная 24 | 2* | 7+8 | 5+10 |
6. | Красная Звезда | 0 | 7+8 | 2+10 |
7. | Память 47, Алтын масак | 2* | 17+18 | 5+10 |
8. | Янбаш | 2* | 20 | 5+10 |
9. | Ани 435 | 0 | 13+16 | 2+10 |
10. | Bluegille | 1 | 13+19 | 2+12 |
11. | (TX71A562.6*4/AMI)*4/LARGO | 2* | 14+15 | 5+10/2+10 |
12. | Кокбидай | 2* | 7 | 2+10 |
13. | Brock | 0 | 6+8 | 2+12 |
14. | Кзыл-бас | 0 | 6+8 | 2+10 |
15. | Регенерант Саратовской 52 | 2* | 0+9 | 5+12 |
На их основе проведена идентификация 1082 образцов генофонда КазНИИЗИР, в т. ч.657-озимой мягкой пшеницы из 12 коллекционных наборов, 141 - яровой мягкой пшеницы из 4 наборов, 284-твердой пшеницы из 6 наборов по составу высокомолекулярных глютенинов. Их белковые формулы приведены в изданном нами каталоге (, и др.,2008).
3.1.3 Каталог блоков компонентов гордеина для регистрации коллекционных форм ячменя Идентификация генотипов ячменя осуществляется исследователями стран СНГ в соответствии с методическими указаниями по сортовой идентификации ячменя на основе эталонных спектров проламинов (Гаврилюк и др.1975), на основе составляющих сорт проламиновых биотипов (Перуанский и др., 1985), на основе блоков компонентов, контролируемых определенными аллелями гордеинкодирующих локусов (Поморцев и др., 1994). Однако ни одно из них в полной мере не отвечает возросшим в настоящее время потребностям в анализе коллекционного и селекционного материала. Так, использование крахмального геля, формирование трубчатых гелей для электрофореза в одних методических рекомендациях, система регистрации компонентов белка у других не позволяет быстро и эффективно интерпретировать полученные электрофореграммы. Блочная номенклатура более трудоемка на начальных стадиях отработки, но по своей применимости в идентификации, регистрации и дифференциации образцов генофонда, селекционных линий превосходит номенклатуру, основанную на регистрации компонентов по относительной электрофоретической подвижности. В то же время форма записи белковой формулы на основе характеристики каждого компонента в пределах a-, b - и w- зон (ОЭП, интенсивность) бывает необходима при сравнении генетически измененных путем биотехнологии, мутагенезом форм с контрольными образцами для выяснения и указания деталей изменчивости а также при передаче сорта в государственное сортоиспытание. Схема перехода от белковых формул, составленных по относительной электрофоретической подвижности компонентов в геле и их локализации в пределах фракций, к блочной номенклатуре пшеницы предлагалась рядом исследователей (Sasek, Cherny, 1985, Абсаттарова, 2002). Для практического применения эта система недостаточна, поскольку авторами изучено ограниченное разнообразие блоков компонентов глиадин-кодирующих локусов. На основе анализа запасных белков большого набора разнообразного сортового и коллекционного материала, нами разработан и предложен для практического использования каталог, включающий 30 вариантов блоков компонентов гордеина локуса Hrd A, 36 вариантов блоков компонентов гордеина локуса Hrd В, 13 вариантов блоков компонентов гордеина локуса Hrd F, которые кроме цифровой идентификации охарактеризованы по относительной электрофоретической подвижности (ОЭП) составляющих их компонентов. Запись белковой формулы можно вести в двух вариантах: краткой, например для сорта Донецкий 8 (№1 на рисунке 4) она будет выглядеть: 5 1 2, для сорта Донецкий 9 (№4 на рисунке 4)– Белковая формула этих же сортов с указанием позиций компонентов в геле записывается так : (Донецкий 8) a 5/69; 4/71; 3/74; b 5/50; 4/53;3/56;3/57;2/58;1/61; w 7/17;6/19;5/21;4/23;3/25; (Донецкий 9): a 5/68; 4/71; 3/73; b 5/50; 4/54; 3/57; 2/58; 2/60 w 8/15; 7/16; 6/18; 5/21; 4/24; На основе разработанного каталога составлены гордеиновые формулы 757 образцов генофонда ячменя, документированные в издании «Каталог генофонда ячменя по спектру гордеина» (, , 2007).
|
|
|



9
HRD А5 (1); HRD А1 (2,3,4,6,8); HRD А3 (5); HRD А2 (7); HRD А11 (9);HRD В1 (1,2,3); HRD В3 (4); HRD В4 (5); HRD В2 (6-9).
HRD F2 (1); HRD F3 (2,3); HRD F4 (4,5); HRD F1(6-9).
Рисунок 4 – Наиболее распространенные варианты блоков компонентов гордеина
3.1.4 Биохимические маркеры скороспелости ячменя Длина вегетационного периода является основным фактором получения хорошего урожая ячменя в условиях горной и предгорной зон Юго-Востока Казахстана (, 1997). Биохимические механизмы, лежащие в основе морфофизиологических процессов, обеспечивающих опережающее развитие скороспелых сортов и закономерности их наследования, изучены недостаточно.
Нами установлено, что в фазе восковой и полной спелости скороспелые формы имеют более высокий уровень свободного пролина в сравнении с позднеспелыми образцами (рисунок5). Качественные и количественные изменения в спектре гор-деина показали, что по мере прорастания зерна происходит деградация компонентов запасных белков g, b и a зон, причем активнее и на более ранних этапах расходуется b50 компонент, медленнее остальных деградирует компонент w зоны с ОЭП 18, (рисунок 6). Отмечено, что у скороспелого генотипа даже в эндосперме зерна на стадии 7-ми суточного прорастания относительное содержание компонента w6/18 составляет 77,5% от изначального уровня в покоящемся зерне, тогда как у позднеспелого генотипа его содержание на этом этапе резко снижено и составляет лишь 16,4% от изначального уровня. Практически полный распад быстроподвижных компонентов a, b и g зон у позднеспелой формы отмечается уже на стадии 3-х суточного развития, тогда как у раннеспелой формы компоненты этих зон проявляются еще достаточно четко. Одной из причин более замедленного распада гордеина у скороспелых форм может быть пул свободных аминокислот покоящегося зерна, не включенных в протеины во время созревания.

 |
М. – молочная спелость; В. – восковая спелость; П. – полная спелость
Рисунок 5- Накопление свободного пролина в зерне скороспелого и позднеспелого генотипов ярового ячменя на стадиях созревания.
Биохимический анализ зерна на стадия покоя показал, что все скороспелые формы характеризовались более высоким содержанием неинкорпорированной в белки аминокислоты пролин. Полученные результаты свидетельствуют, что повышенное содержание свободного пролина в зерне ячменя на стадии полной спелости, а также интенсивность распада запасного белка эндосперма целесообразно использовать в дифференциации селекционных линий, коллекционных образцов и сортов.
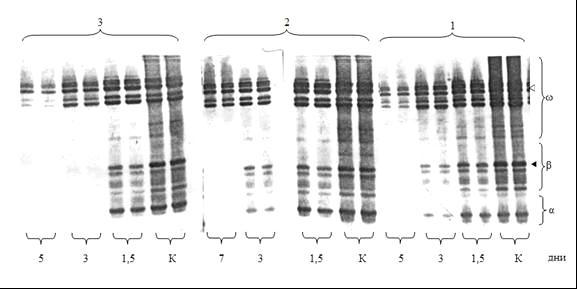

1 – S-26-3 (скороспелый); 2 – Б3. кл. 21(скороспелый); 3 – 33/86-11 (позднеспелый), v - w18, ƒ- b50.
Рисунок 6 - Качественные и количественные изменения в спектре гордеина при прорастании зерна ячменя.
3.1.5 Диагностика и отбор засухоустойчивых образцов ячменя Разработка биохимических тестов засухоустойчивости проводилась путем изучения:
а) влияния высоких температур на генотип (жаростойкость);
б) влияния дефицита влаги в корнеобитаемой среде на генотип (устойчивость к дефициту влаги);
в) совместного влияния двух (а и б) стрессовых факторов на генотип.
Воздействие внешних факторов вызывает в растительном организме изменения, проявляющиеся на различных уровнях его организации. Нами изучалось содер-жание свободной аминокислоты пролин в проростках ярового ячменя и характер изменения активности фермента пероксидазы этих же объектов под действием повышенных температур. С целью разработки методических условий отбора устойчивых к стрессам генотипов ярового ячменя содержание свободного про-лина определялось на проростках с различными сроками развития: 7, 10- и 12-дневных образцах двух перспективных номеров из питомника конкурсного сортоиспытания: 30/83-17 (неустойчивый к жаре), 110жароустойчивый). Уровень аминокислоты у опытных образцов, подвергшихся влиянию высоко-температурного стресс-фактора (1 часовое воздействие t +450C), резко возрастал при всех изученных сроках развития, однако наибольшего значения он достигал у 10-дневных проростков (рисунок 7). Тестирование проростков 45 образцов ярового ячменя позволило ранжировать их по относительному накоплению пролина и определить размах колебаний по значениям ее концентрации в опытных и кон-трольных вариантах, выделить перспективные в плане жаростойкости образцы и использовать их для дальнейших исследований. Анализ характера накопления свободного пролина в проростках биотипов ярового ячменя Сауле до (t+250C) и после воздействия высокотемпературного стресса (1 час при t+450C) показал (таблица 5), что интенсивность его аккумуляции наиболее высока у биотипов №4, №5, №7, №8 и №9. Доля первых трех биотипов является преобладающей в сорте, предназначенном для возделывания в неполивных условиях.
Известно, что под влиянием стрессовых факторов в клетках растений усиливается биологическое окисление и накопление токсичных продуктов обмена. В регуляции этих процессов участвует антиоксидантный фермент пероксидаза. Воздействие нарастающего температурного стресса на проростки выявило термо-стабильность изопероксидазы С100 у жаростойких генотипов ячменя. Для дифферен-циации устойчивых к дефициту влаги форм были подобраны следующие условия: воздействие на 10 дневные проростки 10 мМ раствора полиэтиленгликоля (ПЭГ) с молекулярной массой 17000 в течение 72 часов. При воздействии ПЭГа на проростки ячменя (имитация дефицита влаги) повышается общая пероксидазная активность и идет перераспределение активности в сторону быстропод-вижных изоформ с ОЭП 90 и 93. Совместное воздействие высокотем-пературного стресса и дефицита влаги на проростки также вызывало повышение уровня свободного пролина, тогда как изменение активности изоформ пероксидазы было неоднозначным. Реакция растений ячменя на засу-ху оценивалась не только на стадии проростков в условиях ее искусственной имитации, но и в фазу трубкования в полевых условиях. В результате проведенных исследований установлено, что большая интенсивность накопления свободного пролина у ячменя на стадии проростков во время стресса наблюдалась у засухоустойчивых форм, причем эта тенденция сохранялась на всех изученных этапах развития. Общая активность пероксидазы проростков и активность ее изоформ также варьировали в зависимости от устойчивости растительных объектов
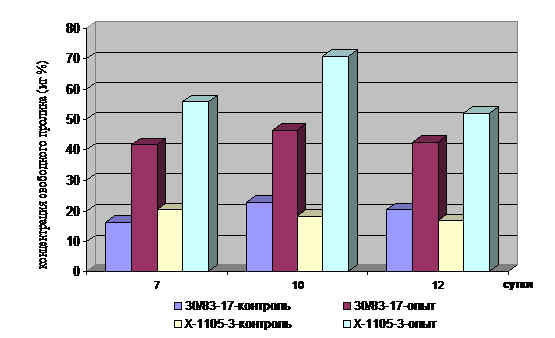 |
Рисунок 7 - Накопление свободного пролина у разновозрастных проростков ячменя в условиях температурного стресса
Таблица 5 - Интенсивность накопления свободного пролина в проростках биотипов ячменя сорта Сауле до (контроль, К) и после (опыт, О) воздействия высокотемпературного стресса
№п/п гордеи-новых биоти-пов сорта Сауле | Содержание свободного пролина в пророст-ках при t+250C (К), мг/% | Содержание свободного пролина в проростках при 1 час возд. t+450C (О), мг/% | *N (O/K) |
1 | 5,6±0,2 | 18,0±1,1 | 3,2 |
2 | 4,7±0,3 | 19,9±1,5 | 4,2 |
3 | 3,3±0,1 | 16,0±1,0 | 4,8 |
4 | 5,2±0,2 | 28,1±0,9 | 5,6 |
5 | 4,3±0,7 | 30,1±2,0 | 7,6 |
6 | 4,5±0,2 | 15,8±0,8 | 3,5 |
7 | 4,6±0,2 | 24,0±2,3 | 5,2 |
8 | 3,2±0,3 | 16,4±0,8 | 5,3 |
9 | 3,3±0,3 | 23,6±0,1 | 7,2 |
НСР 0,95 | 0,67 | 4,13 | |
*N-кратность повышения концентрации свободного пролина в проростках опытных образцов по отношению к таковой контрольных. |
к засухе. На фоне сходной реакции проростков ячменя как устойчивых, так и неустойчивых к засухе и ее элементам по изменению концентрации свободного пролина, выявлены специфические ответные реакции у устойчивых к высокотемпературному стрессу генотипов. Так, усиление температурного стресса вызывало повышение активности быстромигрирующих в полиакриламидном геле катодных изопероксидаз, у неустойчивых форм, тогда как у устойчивых образцов ячменя аналогичные изоформы проявляли стабильность. На основе полученных научных результатов оформлена заявка на патент «Способ двухступенчатой оценки жаростойкости зерновых», № госрегистрации 2007/0593.1, суть которой состоит в том, что проростки испытуемых и стандартного (относительно которого ведут селекцию) генотипов подвергают в камере с освещением лампой накаливания в 500 вт воздействию температурой (450-480 С) в течение часа, другую партию растений (контрольная) оставляют при температуре 20-250 С, в половине обеих партий проростков определяют концентрацию свободного пролина и по их соотношению производят первичную браковку генотипов, у второй части не выбракованных генотипов определяют относительную активность катодной изопероксидазы с ОЭП 100 и генотипы у которых соотношение активностей фермента с ОЭП 100 опытных и контрольных растений ниже единицы, относят к жаростойким, а генотипы с показателями выше единицы, но ниже показателя стандартного генотипа относят к перспективным жаростойким формам.
3.2 Идентификация коллекционных образцов генофонда зерновых колосовых культур Идентификация сорта, линии, генотипа зерновых колосовых – важнейшее звено в организации мероприятий по сбору, хранению и воспроизводству коллекционного материала, по отбору и контролю перспективных линий в селекционных питомниках, установлении типичности сорта в процессе семеноводства.
3.2.1 Идентификация и оценка генетического разнообразия коллекционных образцов мягкой пшеницы В результате анализа состава ВМСГ 16 коллекционных наборов пшеницы показано, что глютенинкодирующий локус Glu А1 представлен 3-мя аллелями (таблица 6), кодирующими субъединицы 1, 2* (a, b) и «молчащую» аллель (с), при которой в спектре отсутствует самая медленно подвижная субъединица. По локусу Glu 1В выявлены носители 9 аллелей (a, b, c, d, e, f, g, h, i), из которых аллель “с”, кодирующая субъединичную пару 7+9 и аллель “b”, ответственная за биосинтез субъединичной пары 7+8 наиболее распространены и встречаются в генофонде с частотой 55,7% и 26,9% соответственно. Локус Glu D1 представлен в изученном материале 4-мя аллелями, из которых наиболее распространены -“d” и “e” (встречаются с частотой 35,5% и 30,5%, соответственно). Анализ состава глиадинов позволил выявить по присутствию блока компонентов глиадина Gli 1B3 образцы, имеющие пшенично-ржаную транслокацию1BL/ 1RS. Генетическое разнообразие (H) изученных коллекций по числу и частоте встречающихся аллелей 3-х глютенинкодирующих локусов варьировало от 0,28 до 0,64. Более 70% изученного материала генофонда мягкой пшеницы КазНИИЗ являются носителями ценных в отношении качества аллелей локусов Glu A1 и Glu B1, более 50% - локуса Glu D1. Результатом использования методов белкового маркирования в селекции мягкой пшеницы явилось соавторство в передаче на ГСИ сортов озимой мягкой пшеницы «Кызылбидай 70», с составом высокомолекулярных субъединиц глютенина: 2* (GluA1); 7+9 (GluB1); 5 +10 (GluD1) и «Казахстан-16», с составом ВМСГ типичного биотипа: 2* (GluA1); 7+9 (GluB1); 5 +10 (GluD1), сопутствующего (№2): 2*(GluA1); 7+8 (GluB1);5 +10 (Glu D1).
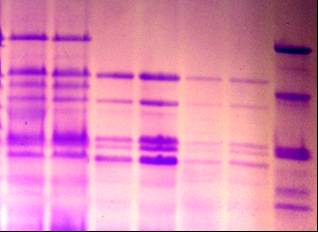
3.2.2 Идентификация и оценка генетического разнообразия коллекционных образцов Triticum durum В генофонде твердой пшеницы, выявлены носители 4 аллельных вариантов локуса Glu - A1 (таблица 7, рисунок 8), из них “null” аллель (отсутствие самой медленно-подвижной субъединицы) и аллель, контролирующая биосинтез ВМСГ 1, наиболее распространены (59,8% и 23,5% соответственно).
|
9 10 |
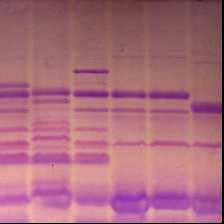

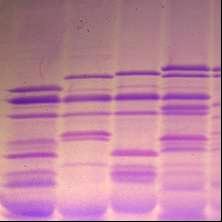

ВМСГ, кодируемые локусом Glu 1A (указаны стрелкой): 2* (1,7); 1* (8); 1 (3,9), и локусом Glu 1В ( треугольник): 7+8 (1,3,4,5,7,8,9); 13+16 (2);, мягкая пшеница Стекловидная 24 с составом ВМСГ 2*; 7+8; 5+10 (10).
Рисунок 8 – Высокомолекулярные субъединицы глютенина твердой пшеницы.
По локусу Glu В1 среди образцов твердой пшеницы выявлены носители 5 аллель-ных вариантов (d, b, e, f, h), ответственных за биосинтез субъединиц : 6+8; 7+8; 20; 13+16; 14+15, соответственно. Генотипы с субъединицами 7+8 наиболее распро-странены в генофонде твердой пшеницы КазНИИЗ и составляют 59,1%, на долю но-сителей субъединичной пары 6+8, сопряженной с высоким качеством макарон, приходится 14,8% образцов. Более 80% образцов генофонда твердой пшеницы явля-ются носителями ценных в отношении качества аллелей локуса Glu В1. Более 50% образцов коллекций генофонда твердой пшеницы характеризуется присутствием ценного варианта низкомолекулярных глютенинов, кодируемых локусом Glu-B3. Наиболее высоким уровнем генетического разнообразия характеризуются: коллек-ционный фонд отдела зерновых колосовых культур КазНИИЗ и коллекция яровой твердой пшеницы Шалкарской ОС (0,54), наименее выражен этот показатель у питомника стран ЦАЗ (0,39). Результатом использования молекулярного маркирования в селекции твердой пшеницы явилось соавторство в передаче в 2008г. на ГСИ сорта яровой твердой пшеницы «Ертол» с составом высокомолекулярных и низкомолекулярных глютенинов: 1 (Glu A1); 7+8 GluB1); 2 (Glu B3).
Таблица 6 -Разнообразие аллелей глютенинкодирующих локусов в коллекционных образцах генофонда пшеницы
№ колл. наборов1 | Н | Локусы ВМСГ | |||||||||||||||
Glu A1 | Glu B1 | Glu D1 | |||||||||||||||
0 c | 1 a | 2* b | 7+9 c | 7+8 b | 6+8 d | 17+18 i | 13+16 f | 14+15 h | 13+19 G | 20 e | 7 a | 5+10 d | 2+10 e | 5+12 ? | 2+12 a | ||
1 | 0,41 | 4,2 | 95,8 | 67,3 | 24,5 | 2,0 | 6,1 | 50 | 25,9 | 18,5 | 5,5 | ||||||
2 | 0,59 | 36,5 | 63,4 | 50 | 25,0 | 4,0 | 10,0 | 8 | 2,0 | 41,8 | 45,5 | 5,5 | 7,3 | ||||
3 | 0,63 | 22,8 | 13,0 | 64,3 | 48,7 | 32,5 | 11,5 | 4,2 | 1,6 | 0,5 | 0,5 | 0,5 | 33,5 | 34,5 | 17,7 | 14,4 | |
4 | 0,64 | 44,7 | 2,6 | 52,6 | 36,7 | 43,3 | 10,0 | 10,0 | 32,4 | 44,1 | 14,7 | 8,8 | |||||
5 | 0,53 | 13,6 | 86,3 | 63,6 | 15,9 | 11,4 | 9,1 | 27,3 | 31,8 | 20,4 | 20,4 | ||||||
6 | 0,64 | 30,0 | 2,5 | 67,5 | 47,5 | 13,7 | 12,6 | 10,0 | 2,5 | 13,7 | 23,8 | 31,3 | 18,8 | 26,3 | |||
7 | 0,44 | 8,7 | 91,3 | 50,0 | 50,0 | 25 | 8,3 | 50 | 16,7 | ||||||||
8 | 0,39 | 12,5 | 87,5 | 87,5 | 12,5 | 37,5 | 25,0 | 12,5 | 25,0 | ||||||||
9 | 0,51 | 73,7 | 26,3 | 29,4 | 5,9 | 52,9 | 5,9 | 5,9 | 23,5 | 11,8 | 64,7 | ||||||
10 | 0,48 | 20 | 4,0 | 76,0 | 16.7 | 56.7 | 6,7 | 6,7 | 3,3 | 3,3 | 3,3 | 3,3 | 78,6 | 14,3 | 7,1 | ||
11 | 0,55 | 6,2 | 93,8 | 38,1 | 38,1 | 23,8 | 42,1 | 31,6 | 5,3 | 21,1 | |||||||
12 | 0,49 | 12,9 | 4,1 | 83,0 | 72,7 | 21,7 | 2,8 | 2,8 | 38,5 | 40,6 | 7 | 14,0 | |||||
13 | 0,41 | 3,8 | 96,1 | 70,4 | 29,6 | 15,4 | 20,9 | 35,8 | 28,2 | ||||||||
14 | 0,31 | 3,6 | 96,4 | 91,2 | 7,0 | 1,8 | 38,6 | 10,5 | 43,9 | 7 | |||||||
15 | 0,59 | 22,9 | 11,5 | 65,6 | 55,7 | 39,3 | 1,6 | 3,3 | 44,3 | 16,4 | 18,0 | 21,3 | |||||
16 | 0,28 | 71,4 | 28,6 | 100 | 28,6 | 71,4 | |||||||||||
Генофонд в целом | 20,5 | 5,3 | 74,1 | 55,7 | 26,9 | 6,6 | 6,0 | 0,9 | 0,1 | 0,2 | 0,6 | 2,9 | 35,5 | 30,5 | 19,5 | 14,4 | |
1 №1- сорта озимой мягкой пшеницы Казахстанской селекции; №2 - сорта озимой мягкой пшеницы стран ЦАЗ ; № 3- образцы озимой мягкой пшеницы питомника восточно-европейского сортоиспытания ;№4 – перспективные линии озимой мягкой пшеницы; №5 - образцы озимой мягкой пшеницы института пшеницы и подсолнечника г. Добруджа (Болгария); №6 –коллекционные образцы озимой мягкой пшеницы США; №7 - образцы озимой мягкой пшеницы селекционно-генетического института – национального центра семеноведения и сортоизучения УААН ; №8 - образцы озимой мягкой пшеницы Мироновского института пшеницы им УААН; №9- образцы озимой мягкой пшеницы международного института пшеницы, г. Кембридж (Англия); №10- сорта и линии озимой мягкой пшеницы Красноводопадской CОС (Казахстан); №11- устойчивые к ржавчинным болезням линии озимой мягкой пшеницы НИИ проблем биологической безопасности НЦБ МОН РК; №12 - перспективных по качественным показателям линии озимой пшеницы; №13 – сорта яровой мягкой пшеницы; №14 – образцы яровой мягкой пшеницы питомника 1-ый КАСИП; №15 – образцы яровой мягкой пшеницы 4-ого КАСИП; №16- староместные сорта яровой мягкой пшеницы Западного Памира (Таджикистан). | |||||||||||||||||
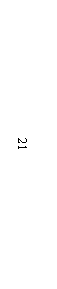
Таблица 7 - Разнообразие аллелей глютенинкодирующих локусов коллекционных образцов генофонда твердой пшеницы
№ колл. набо- ров1 | Кол-во об-раз-цов | Н | Локусы ВМСГ и НМГ | ||||||||||
Glu –A1 | Glu-B1 | Glu-B3 | |||||||||||
«0» | 1 | 1* | 2* | 6+8 | 7+8 | 20 | 13+ 16 | 14+ 15 | НМГ 1 | НМГ 2 | |||
1 | 11 | 0,39 | 100 | - | - | - | 21,4 | 28,6 | 50 | - | - | 33,3 | 66,7 |
2 | 30 | 0,49 | 47,1 | 44,1 | 2,9 | 5,9 | 8,3 | 77,8 | 5,6 | 8,3 | - | 38,8 | 61,3 |
3 | 16 | 0,53 | 64,8 | 35,2 | - | - | 11,1 | 61,1 | - | 27,8 | - | 43,8 | 56,2 |
4 | 115 | 0,54 | 71,0 | 23,7 | - | 5,3 | 22,3 | 48,3 | 16,8 | 7,7 | 4,9 | 42,2 | 57,8 |
5 | 92 | 0,54 | 44,7 | 17,0 | 1,1 | 37,2 | 7,5 | 70,2 | 9,6 | 9,6 | 3,2 | 43,6 | 56,4 |
Генофонд в целом | 59,8 | 23,5 | 0,8 | 15,9 | 14,8 | 59,1 | 13,3 | 9,5 | 3,4 | 42,4 | 57,6 | ||
1 №1 - сорта озимой твердой пшеницы стран ЦАЗ; №2- образцы яровой твердой пшеницы питомника 1-ый КАСИП; №3- образцы яровой твердой пшеницы питомника 4-ый КАСИП; №4 - образцы мировой коллекции ВИР; №5- коллекционные образцы твердой пшеницы Шалкарской ОС (Казахстан); |
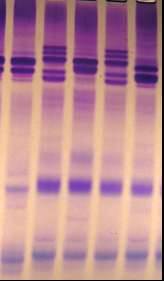
3.2.3 Идентификация и регистрация генофонда ячменя по запасным белкам зерна Генофонд ячменя КазНИИЗиР, изученный по комплексу хозяйственно-ценных признаков, к настоящему времени насчитывает около 900 образцов, из которых 757 были идентифицированы на основе спектра гордеина. По результатам электрофореза запасных белков у 10 коллекционных наборов, обозначения которых приведены в сноске таблицы 10, выявлено 12 вариантов a - зоны белкового спектра (HRDF), 33 варианта b - зоны (HRDB) и 29 вариантов блоков компонентов гордеина w - зоны (HRDA, таблицы 8-10). Наиболее характерные для генофонда блоки HRDA: 1, 2, 3, 6, 9; блоки компонентов HRDB: 2,1,3, 4, 5, 7, 6, 24; блоки HRDF: 1, 2, 5, 4, 7. Степень генетического разнообразия коллекций генофонда ячменя варьирует от 0,52 до 0,88. Значительная часть образцов коллекции (24,1%) полиморфна, в том числе 12,9% по всем 3 гордеинкодирующим локусам, 7,0% - по 2ум и 4,2% - по 1 локусу. Полиморфность локусов и, следовательно, гетерогенность образцов коллекций следует учитывать в ходе их репродуцирования при закладке на хранение, а также при подборе пар для гибридизации в селекционном процессе. Специфичность глютелина ячменя на уровне сорта, биотипа, линии может оказаться полезной в распознавании генотипов с идентичной гордеиновой формулой. Анализ состава запасных белков эндосперма коллекционных образцов ячменя, проведенный методом ДСН - электрофореза, выявил 3 варианта ВМСГ, обозначенных нами D1, D2 и D3 (рисунок 9, дорожки № 6,8,7, соответственно). Субъединицы D1 и D2 наиболее распространены у сортов, биотипов и коллекционных форм, тогда как субъединица D3 была выявлена лишь у единичных образцов.
Таблица 8 - Генетическое разнообразие коллекционных образцов генофонда ячменя КазНИИЗиР по локусу HrdA
№ колл. набора | Варианты блоков компонентов гордеина, частота встречаемости (%) | ||||||||||||||||||||||||||||
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 17 | 18 | 19 | 20 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 32 | |
1 | 44,7 | 25,5 | 25,5 | 0 | 0 | 0 | 2,1 | 2,1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
2 | 75,4 | 14 | 5,3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3,5 | 1,7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
3 | 38,5 | 13,8 | 29,3 | 0 | 6,4 | 0,9 | 1,8 | 0,9 | 0 | 1,8 | 4,6 | 0 | 1,8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
4 | 41,7 | 10,3 | 7,9 | 0,6 | 5,4 | 6 | 1,2 | 1,5 | 1,8 | 4,8 | 2,4 | 0,9 | 3,6 | 3,3 | 0,6 | 1,2 | 0,3 | 3 | 0 | 0,6 | 0,3 | 0,6 | 1,5 | 0 | 0 | 0 | 0,3 | 0 | 0 |
5 | 52,2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4,4 | 4,4 | 13 | 0 | 0 | 0 | 0 | 8,7 | 4,3 | 13 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
6 | 13 | 0 | 4,4 | 8,6 | 0 | 4,4 | 0 | 4,4 | 17,3 | 4,4 | 4,4 | 8,7 | 0 | 0 | 0 | 0 | 8,7 | 17,3 | 0 | 0 | 4,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
7 | 39,1 | 0 | 0 | 4,4 | 0 | 13 | 0 | 17,3 | 13 | 4,4 | 0 | 0 | 0 | 0 | 0 | 0 | 4,4 | 0 | 0 | 0 | 4,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
8 | 29,4 | 0 | 0 | 2 | 0 | 0 | 0 | 5,9 | 49 | 7,8 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 |
9 | 19,6 | 1,9 | 10,1 | 7,6 | 0 | 7,6 | 0 | 0 | 18,4 | 8,2 | 1,9 | 1,3 | 0 | 0 | 0 | 0,6 | 3,2 | 5,1 | 0,6 | 1,9 | 2,5 | 0 | 5,7 | 0,6 | 1,3 | 0,6 | 0,6 | 0,6 | 0 |
10 | 28,2 | 15,5 | 1,4 | 0 | 0 | 8,5 | 0 | 22,5 | 4,2 | 1,4 | 0 | 2,8 | 0 | 0 | 0 | 1,4 | 0 | 8,5 | 0 | 0 | 0 | 1,4 | 1,4 | 0 | 1,4 | 0 | 0 | 0 | 1,4 |
Гено-фонд | 36,9 | 8,4 | 9,3 | 2,1 | 2,9 | 5 | 0,8 | 3,7 | 8,4 | 4,6 | 2,4 | 1,1 | 1,9 | 1,4 | 0,4 | 0,9 | 0,7 | 3,4 | 0,9 | 0,6 | 0,8 | 0,4 | 1,8 | 0,1 | 0,4 | 0,1 | 0,4 | 0,1 | 0,1 |
![]()
Таблица 9- Генетическое разнообразие коллекционных образцов генофонда ячменя КазНИИЗиР по локусу HrdB
№ колл. набора | Варианты блоков компонентов гордеина, частота встречаемости (%) | ||||||||||||||||||||||||||||||||
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | |
1 | 25,5 | 38,3 | 10,6 | 17 | 0 | 0 | 0 | 4,3 | 0 | 0 | 0 | 2,1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2,1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
2 | 20,7 | 62,1 | 8,6 | 6,9 | 0 | 0 | 0 | 0 | 1,7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
3 | 25,2 | 22,6 | 13,9 | 25,2 | 0,9 | 0,9 | 4,3 | 4,3 | 1,7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,9 |
4 | 19,4 | 19,8 | 12,5 | 6,4 | 7,8 | 6,3 | 9,3 | 2,9 | 4,1 | 2,9 | 1,5 | 0,9 | 0,9 | 0 | 0 | 0 | 0 | 0,3 | 0,3 | 0 | 0 | 0 | 1,2 | 0,6 | 0,6 | 0,3 | 1,2 | 0,3 | 0 | 0 | 0,6 | 0 | 0 |
5 | 0 | 8,7 | 47,8 | 0 | 21,7 | 0 | 4,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4,4 | 0 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | 0 | 0 |
6 | 0 | 12,5 | 16,7 | 8,3 | 25 | 8,3 | 12,5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 16,7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
7 | 0 | 17,4 | 26,1 | 4,4 | 0 | 21,7 | 8,6 | 4,4 | 0 | 0 | 0 | 0 | 4,4 | 4,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8,6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
8 | 4 | 28 | 0 | 8 | 0 | 4 | 2 | 2 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 48 | 0 | 0 | 0 | 0 | 0 | 0 |
9 | 3,3 | 7,2 | 10,5 | 1,3 | 20,4 | 9,9 | 15,1 | 0,7 | 0 | 5,9 | 2,6 | 0 | 1,3 | 2,6 | 0,7 | 0,7 | 0,7 | 0,7 | 0 | 0,7 | 0,7 | 0,7 | 9,2 | 0,7 | 0 | 0 | 1,9 | 1,9 | 0,7 | 0 | 0 | 0 | 0 |
10 | 5,3 | 26,7 | 5,3 | 2,7 | 1,3 | 10,7 | 2,7 | 1,3 | 4 | 1,3 | 0 | 0 | 0 | 2,7 | 0 | 0 | 0 | 6,7 | 0 | 0 | 0 | 0 | 24 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5,3 | 0 |
Гено-фонд | 14,4 | 22,2 | 12,1 | 8,1 | 7,7 | 6 | 7,6 | 2,3 | 2,2 | 2,3 | 1 | 0,4 | 0,7 | 0,8 | 0,1 | 0,1 | 0,1 | 0,8 | 0,1 | 0,2 | 0,1 | 0,1 | 4,8 | 0,3 | 0,2 | 0,1 | 3,4 | 0,4 | 0,1 | 0,3 | 0,2 | 0,4 | 0,1 |
|
1-Донецкий 10, 2-Оренбургский 15, 3-№ 000 из Эфиопии, 4- Карабалыкский 43, 5 –Донецкий 8; 6- № № № 000 из Сирии, идентичные по спектру гордеина)
Рисунок 9- Спектр глютелинов коллекционных образцов ячменя
Таблица 10 - Генетическое разнообразие коллекционных образцов генофонда ячменя КазНИИЗиР по составу гордеинов локуса HrdF
№ коллек-ционного набора | Варианты блоков компонентов гордеина, частота встречаемости (%) | |||||||||||
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 10 | 11 | 12 | 13 | |
1 | 40,9 | 27,3 | 18,2 | 11,4 | 2,3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
2 | 61 | 8,5 | 20,3 | 6,7 | 3,4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
3 | 15,2 | 37,5 | 25,9 | 14,3 | 6,25 | 0 | 0,9 | 0 | 0 | 0 | 0 | 0 |
4 | 19,4 | 20,3 | 25,5 | 12,8 | 14,8 | 0 | 7 | 0,3 | 0 | 0 | 0 | 0 |
5 | 8,7 | 0 | 21,7 | 47,8 | 21,7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
6 | 13 | 13 | 21,7 | 17,4 | 26,1 | 0 | 8,7 | 0 | 0 | 0 | 0 | 0 |
7 | 8,7 | 13 | 30,4 | 26,1 | 4,4 | 0 | 13 | 4,4 | 0 | 0 | 0 | 0 |
8 | 28 | 12 | 56 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 |
9 | 7 | 12,1 | 35 | 9,6 | 22,3 | 0,6 | 6,4 | 3,2 | 1,9 | 0,6 | 0,6 | 0,6 |
10 | 27,4 | 13,7 | 9,6 | 5,5 | 30,1 | 0 | 10,9 | 2,7 | 0 | 0 | 0 | 0 |
Всего | 20,9 | 18,7 | 26,8 | 12 | 14,3 | 0,1 | 5,5 | 1,0 | 0,3 | 0,1 | 0,1 | 0,1 |
№1- сорта селекции КазНИИЗиР; №2 и №3 - перспективные селекционные линиии конкурсных питомников КазНИИЗиР 1993 и 2005 годов, соответственно; №4- номера мировой коллекции ВИР; №5- линии ИКАРДА для зон с низким уровнем осадков; №6 - линии ИКАРДА для зон с низким уровнем осадков и холодной зимой; №7- линии ИКАРДА для зон с умеренным уровнем осадков; №8- голозерные шестирядные формы ИКАРДА; №9 - линии ИКАРДА; №10 - линии Иранского происхождения. |
3.2.4 Полиморфные сорта ячменя как источники и доноры селекционно-ценных признаков Внутрисортовой полиморфизм пшеницы и ячменя по составу запасных белков: проламинов и глютелинов, позволяет дифференцировать их на биотипы, объединенных сходством компонентного состава. Соотношение биотипов в сорте меняется в зависимости от условий возделывания, в связи с чем контроль за его биотипным составом по запасным белкам приобретает особую значимость при его семеноводстве и использовании в селекции на конкретные признаки в качестве исходной формы ( 1983, , 1985). Сорт
ярового ячменя Сауле, крупяного направления, был дифференцирован нами на основе качественной характеристики электрофоретического спектра гордеина семян на 9 биотипов (рисунок 10), из которых биотипы № 1,2, 4, 5, и 7 являлись наиболее многочисленными в процентном соотношении (таблица 11). В дальнейшем доля биотипов №1, 2, а также биотипов №6 и 8, изначально незначительных в количественном отношении, снизилась и составила в 2007 г. менее 5% каждого, тогда как биотипы №3 и 9 практически перестали обнаруживаться в популяциях сорта с 1996г. Доля биотипов № 4, 5 и 7 в сорте увеличилась, причем биотип №5 стал преобладающим в процентном отношении в сорте. Этот биотип является наиболее скороспелым и определяет скороспелость сорта в целом. Биотипы различаются по показателям урожайности (таблица 12), качества зерна (таблица 13), устойчивости к стрессовым факторам. По результатам лабораторных оценок биотипы № 4,7,8,9 являются жароустойчивыми, биотип №5 – жароустойчивым и скороспелым. Линии этих и других биотипов сорта Сауле использовались в селекции ячменя на засухоустойчивость и скороспелость.
|
|
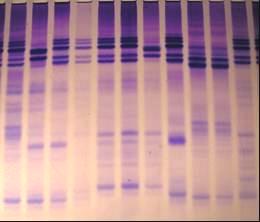

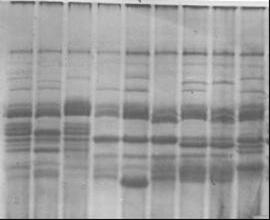

а в
Рисунок 10 - Белковые спектры (а-гордеины; в-глютелины и гордеины) биотипов сорта Сауле. Стрелкой указана субъединица D1.
В сорте ярового ячменя Арна пивоваренного направления выявлено 5 гордеи-новых биотипов, доля типичного биотипа варьировала по годам от 53,3 до 67,9%. Уровень типичного биотипа сопряжен с урожайностью (г=0,69), содержанием крахмала и экстрактивностью (г=0,7 и 0,62 соответственно), в связи с чем при вовлечении сорта Арна в скрещивания, необходимо предварительное маркирование индивидуальных растений и использование генотипов типичного биотипа. Контроль за направленностью сдвига в соотношении проламиновых биотипов в полиморфных сортах ярового ячменя позволяет прогнозировать изменение урожайных, техноло-гических, кормовых достоинств сорта и принимать своевременные меры по сохра-нению оптимального соотношения внутрисортовых групп. Закладку образцов поли-морфных сортов на хранение в генофонд следует вести с учетом соотношения
биотипов в сорте и контролировать при репродукции таких образцов полноценность популяции. Сохранение отдельных биотипов в коллекции предла-гается вести репродуцированием как отдельного образца (, , 2002). Биотипы сорта Сауле переданы в отдел генофонда полевых культур и отдел зернофуражных культур.
Таблица 11- Биотипный состав сорта Сауле по гордеину.
№ био-типа | Блоки компонентов | Cодержание биотипа в популяции (%) | |||||||||||
HRD | 1989 | 1990 | 1993 | 1996 | 1998 | 1999 | 2000 | 2001 | 2002 | 2007 | |||
A | B | F | |||||||||||
1 | 1 | 8 | 2 | 9,8 | 10.0 | 4.2 | 10,4 | - | 4.5 | 6,5 | 8.3 | 4.2 | 1.8 |
2 | 3 | 4 | 2 | 18.3 | 17.0 | 16,6 | 6,3 | 6,0 | 2.3 | - | 5.6 | 4.2 | 3.5 |
3 | 1 | 4 | 2 | 8.5 | 8.0 | 16,6 | - | - | - | - | - | - | |
4 | 1 | 3 | 4 | 18.3 | 20.0 | 25,0 | 4,2 | 10,0 | 13.6 | 4,4 | 12.5 | 8.3 | 12.3 |
5 | 1 | 1 | 3 | 17.0 | 16.0 | 8,3 | 20,8 | 42,0 | 36.4 | 50,0 | 38.9 | 43.8 | 54.4 |
6 | 3 | 1 | 3 | 6.1 | 6.6 | 12,5 | 6,3 | - | 6.8 | 2,2 | 4.2 | 8.3 | 3.5 |
7 | 1 | 2 | 1 | 13.4 | 13.0 | 8,3 | 37,5 | 40,0 | 31.8 | 23,9 | 23.6 | 18.8 | 21.1 |
8 | 7 | 8 | 2 | 4.9 | 5.3 | - | 14,6 | 2,0 | 4.5 | 13,0 | 5.6 | 4.2 | 3.5 |
9 | 1 | 12 | 2 | 3.7 | 4.0 | 8,3 | - | - | - | - | - | - | - |
Таблица 12- Элементы урожайности биотипов Сауле (1998 г.)
№ биоти- па | Высота растений, см. | Продуктив-ная кустис-тость, шт. | Длина колоса, см. | Число зерен в колосе, шт. | Число зерен с растений, шт. | Масса зерен с растений, г. |
1 | 55,40±6,65 | 2,17±0,23 | 6,80±0,47 | 17,43±1,96 | 34,60±8,34 | 1,37±0,43 |
2 | 59,33±5,86 | 2,10±0,21 | 7,20±0,47 | 17,43±2,23 | 34,20±8,43 | 1,40±0,36 |
3 | 58,93±5,32 | 2,23±0,22 | 7,03±0,38 | 17,93±2,97 | 36,73±9,97 | 1,67±0,48 |
4 | 58,27±6,76 | 2,13±0,32 | 6,90±0,50 | 17,60±2,30 | 34,57±8,84 | 1,37±0,43 |
5 | 55,67±6,54 | 2,20±0,25 | 6,83±0,62 | 17,93±1,65 | 35,70±7,56 | 1,57±0,34 |
6 | 52,27±5,36 | 2,13±0,27 | 6,33±0,54 | 14,23±2,32 | 34,10±7,27 | 1,37±0,30 |
7 | 54,07±8,20 | 2,33±0,19 | 7,00±0,79 | 17,50±3,36 | 37,77±10,2 | 1,70±0,52 |
8 | 58,03±5,20 | 3,43±1,19 | 7,07±0,38 | 18,20±1,39 | 36,23±7,71 | 1,63±0,35 |
9 | 59,80±6,80 | 2,23±0,20 | 6,63±0,44 | 17,30±2,75 | 33,97±9,07 | 1,53±0,43 |
Сауле | 55,70±4,47 | 2,07±0,33 | 7,00±0,17 | 16,90±2,55 | 35,73±9,54 | 1,53±0,43 |
НСР | 3,49 | 1,12 | 0,65 | 3,04 | 3,94 | 0,29 |
Таблица 13- Качественные показатели зерна биотипов сорта Сауле
№ биотипа | Содержание св. пролина в зерне, мг/% | Содержание белка,% | Содержание крахмала,% | Экстрактивность,% |
1 | 52,4±5,3 | 11,2±0,3 | 53,7±1,1 | 75,5±0,8 |
2 | 60,6±5,9 | 11,8±0,8 | 52,5±1,4 | 74,7±1,3 |
3 | 65,0±11,6 | 12,6±0,7 | 52,6±0,6 | 74,4±0,2 |
4 | 40,4±2,9 | 10,5±0,3 | 56,2±0,5 | 77,9±0,5 |
5 | 84,4±6,5 | 10,8±0,4 | 55,1±0,6 | 76,7±0,6 |
6 | 38,6±3,7 | 10,7±0,3 | 53,5±0,5 | 76,6±1,1 |
7 | 45,0±2,3 | 12,0±0,5 | 51,3±1,5 | 74,8±1,3 |
8 | 67,9±2,7 | 12,2±0,6 | 52,5±0,8 | 74,4±0,8 |
9 | 61,1±0,7 | 11,8±0,3 | 52,7±0,6 | 74,1±0,9 |
3.3 Биологический анализ в отборе перспективных линий пшеницы и ячменя с хозяйственно-ценными признаками
3.3.1 Высокомолекулярные субъединицы глютенина в оценке хлебопекарных показателей качества мягкой пшеницы Содержание белка, твердозерность, показатель седиментации, состав ВМСГ и, при необходимости, глиадинов являются биологическими тестами, используемыми на ранних этапах селекции пшеницы на качество зерна и муки при ограниченности биологического материала. Нами изучена взаимосвязь между этими и другими технологическими показателями на материале восточно-европейского регио-нального питомника и сортовом наборе мягкой пшеницы селекции КазНИИЗиР. Показано преимущество форм с определенными ВМСГ по таким технологическим показателям как твердозерность и седиментационный тест. Так, образцы с ВМСГ 2*, 1 (аллели “a” и”b” локуса Glu А1), 7+8, 7+9, 17+18, 14+15 (аллели “b”, “c”, “i”, “h” локуса Glu В1) и 5+10, 5+12 (аллели “d” и “нов.” локуса Glu D1) имели более высокие значения этих показателей. В группе образцов с указанными аллелями выявлена более тесная взаимосвязь между содержанием белка и показателем седиментации (варьирование от 0,56 до 0,63). По вкладу в оценку качества по глютенину “Glu 1 score” эти субъединицы высокоранжируются (от 2 до 3 баллов). Проявление уровня седиментации в зависимости от состава ВМСГ (рисунок 11) показало, что наиболь-
шее значение этого пока-зателя было присуще носите-лям аллелей “a” и “b” локуса Glu A1, “i” – локуса GluB1 и “d” - локуса Glu D1, вклад которых в оценку качества по глютенину определен в 3 и 4 балла. В то же время для групп, у которых отсутствует самая медленноподвижная субъединица, контролируе-мая локусом Glu A1 («null» аллель), субъединицами локуса Glu B1 являются: |
|
7 и 6+8, локуса Glu D1 – 2+10/2+12 характерно наименьшее значение показателя седиментации, что согласуется с их низким ранжированием по вкладу в качество, отмеченным ранее (Payne P. I., et. al., 1987). У носителей указанных аллелей твердозерность более тесно сопряжена с седиментационным тестом (r = 0,87-0,92). Носители аллели “d” локуса Glu В1, у которых в спектре ВМСГ присутствует пара субъединиц 6+8, характеризовались низким содержанием белка и клейковины. Рекомендуется оценку технологических свойств зерна и муки селекционных образцов пшеницы вести в пределах групп, предварительно дифференцированных по составу ВМСГ, с учетом сопряженности тех или иных показателей качества.
3.3.2 Белковые маркеры в создании перспективных линий озимой пшеницы с эффективными генами устойчивости к желтой ржавчине Известен ряд генов устойчивости к желтой ржавчине, эффективных в различных регионах Центральной Азии и Закавказья, в т. ч. и ген Yr15, имеющий происхождение от дикого сородича пшеницы Triticum dicoccoides Koern. и локализованный на коротком плече хромосомы 1В в 7сМ от центромеры (McIntosh RA et. al., 1996). Анализ показал, что изогенная линия сорта яровой мягкой пшеницы Aroona с геном Yr15 имела специфичную пару ВМСГ: 14+15 (рисунок 12, отмечены фигурной скобкой), контролируемых локусом Glu B1, который, как известно, находится на длинном плече хромо-сомы 1В и также тесно сцеплен с центромерой (Payne P. I. ey. al., 1982). С целью определения ценности специфичных субъединиц в маркировании устойчивости к желтой ржавчине, обусловленной присутствием гена Yr 15,
|

9
1-Aroona/Yr8; 2-Aroona/Yr17; 3-Aroona/Yr15; 4-Aroona; 5-Богарная 56 (стандарт); 6-Саратовская 29; 7-Avoset; 8-Avoset/Yr15; 9-Avoset /Yr8.
Рисунок 12 - изогенные линии сортов яровой мягкой пшеницы Aroona и Avoset с генами устойчивости к желтой ржавчине
из комбинаций озимых мягких пшениц Красноводопадская 210 х Aroona Yr15; и Стекловидная 24 х Aroona Yr15 были выделены и оценены по проростковой устойчивости линии F4 поколения. Значительная часть линий с вариантами субъединиц 14+15 оказались иммунными или устойчивыми к возбудителям желтой
Таблица 13 – Сопряженность ВМСГ локуса GluB1 c устойчивостью к желтой ржавчине
Гибридная комбинация | Общее число линий | Число иммунных линий | Число линий с ВМСГ 14+15 | % линий с ВМСГ 14+15 от числа иммунных |
Красноводопадская 210 хYr15 | 20 | 12 | 10 | 83,3 |
Стекловидная 24 х Yr15 | 22 | 18 | 9 | 50,0 |
ржавчины (таблица 13). На основе контролируемого переноса гена устойчивости к желтой ржавчине Yr15 нами cозданы 52 устойчивые и иммунные к возбудителям болезни гибридные линии мягкой пшеницы. По результатам полевых и лабораторных оценок из гибридной комбинации Стекловидная 24 х Aroona Yr15 выделена иммунная к желтой ржавчине линия №34, с благоприятным в отношении качества сочетанием высокомолекулярных субъединиц глютенина и перспективная по продуктивности (таблица 14). Выделенные линии переданы отделу зерновых колосовых культур КазНИИЗиР. Сорт озимой мягкой пшеницы Шарора, несущий
Таблица 14 - Элементы продуктивности перспективных линий озимой мягкой пшеницы, устойчивых к ржавчинным болезням.
Происхождение линии | Высота растений, см. | Продуктивная куст., шт. | Длина гл. коло-са, см | Число зерен в гл. кол. шт. | Число зерен в раст., шт. | Масса зерен с гл. ко-лоса.,г. | Масса зерен с раст.,г. | Масса 1000 зерен, г. |
Yr 15* | 89,0±6,23 | 3,9±0,23 | 6,2±0,50 | 25,5±1,4 | 86,3±6,90 | 1,0±0,1 | 3,5±0,18 | 44,6±2,3 |
Ст24**хYr15 | 119,5±9,5 | 4,2±0,38 | 8,5±0,60 | 33,0±2,8 | 102,6±9,23 | 1,5±0,1 | 4,6±0,29 | 46,2±2,5 |
Ст. 24 | 130,4±11,7 | 3,9±0,20 | 8,5±0,77 | 39,3±2,9 | 129,1±7,23 | 2,0±0,1 | 6,5±0,46 | 52,6±3,3 |
(ст.)Алмалы | 113,0±7,9 | 2,8±0,25 | 9,2±0,69 | 35,9±2,4 | 86,2±6,21 | 1,7±0,1 | 3,9±0,28 | 44,6±2,9 |
НСР 0,95 | 27,88 | 0,98 | 2,08 | 9,35 | 32,18 | 0,67 | 2,12 | 6,06 |
*-изогенная линия с геном Yr15; **- Стекловидная 24 |
пшенично-ржаную транслокацию, был использован в качестве источника гена Yr9 в скрещивании с неустойчивым к ржавчине сортом Стекловидная 24. На основе маркирования гибридных линий блоком глиадина 1B3 созданы 2 имунные к желтой ржавчине перспективные линии, которые также проходят испытание в питомнике КСИ отдела зерновых колосовых культур.
3.3.3 Биологические тесты в селекции засухоустойчивых линий ячменя
Путем скрещивания засухоустойчивых и скороспелых биотипов сорта Сауле и коллекционных образцов ячменя с высокоурожайными но неустойчивыми формами, были получены гибридные популяции, из которых выделено 355 высокоурожайных линий. По результатам тестирования на константность по спектру гордеина для дальнейшей селекционной проработки были отобраны гомозиготные линии, из числа которых проведен отбор урожайных гибридных линий со способностью к интенсивному накоплению свободного пролина при высокотемпературном и осмотическом стрессах. С 2005 в контрольном и конкурсном питомниках отдела зернофуражных культур и кукурузы КазНИИЗиР, в экологическом сортоиспытании находились 7 перспективных линий, из них линии № 25, 30 и 31 (таблица 15) выделились по результатам экологического испытания, линия 61/7 вошла в число перспективных сортообразцов и находится в
настоящее время в питомнике конкурсного сортоиспытания отдела
Таблица 15 - Биохимические показатели засухоустойчивости и скороспелости перспективных линий ячменя
№ ли-нии | Комбинации | Интенсивность накопления св. пролина в проростках при | Содержание св. пролина в зерне, мг/% | |
высоко - темпера-турном стрессе | дефиците влаги | |||
25 | Б4 кл.51 х Бота | 1,5 | 4,5 | 117,6 |
30 | Б4 кл.51 х Бота | 1,5 | 4,8 | 104,8 |
31 | К1 34хИран4470 | 1,6 | 5,3 | 84,4 |
НСР 0,95 | 18,94 |
Таблица 16 –Характеристика перспективной линии ячменя 61/7 питомника конкурсного сортоиспытания, 2007г.
Сорта | Длина вегетационного периода, дни | Высота растений, см. | Продуктивная кустистость, шт. | Длина колоса, см. | Число зерен в колосе, см. | Масса 1000 зерен, г. | Урожайность, ц/га | Содержание белка, % |
Арна (ст.) | 66 | 77,6 | 1,7 | 9,3 | 26 | 48,0 | 39,5 | 12,5 |
№ 61/7 | 66 | 79,3 | 1,8 | 9,0 | 25 | 50,0 | 48,5 | 11,1 |
НСР 0,95 2,2 |
Таблица 17- Этапы и виды биохимических тестов в селекции засухоустойчивых и скороспелых форм ячменя
![]()
Питомники | ||||||||
Селекционная задача | Коллекционный | Гибрид-ный | СП1 | СП2 | КП | КП | КСИ | КСИ |
Подбор исходных форм | Гетеро-зигот-ность | Константность | Выделение перспектив-ных линий | Оценка засухоустойчивости и скороспелости | Выделение перспек-тивных линий в КСИ | Сортоиспытание, выделение лучших линий | Сортоис-пытание, выделение лучших линий | |
Исполь-зуемые биохими-ческие показатели | Интенсивность накопления свободного пролина, активность изоперокси- дазы С100, спектр гордеина | Спектр гордеи-на | Спектр гордеина | Спектр гордеина, содержание белка, содержание св. пролина в зерне | Спектр гордеина, содержание св. пролина в проростках и зерне, активность пероксидазы и ее катодных изоформ. | Спектр гордеина | Спектр гордеина | Спектр гордеина |
зернофуражных культур и кукурузы (таблица 16). Линия получена из гибридной комбинации, исходными формами для которой были: линия биотипа №5, скороспелого и жароустойчивого, и линия чувствительного к высокотем-пературному стрессу биотипа №2 сорта ярового ячменя Сауле.
В таблице 17 приведен комплекс биохимических тестов, проявивших эффективность в селекции засухоустойчивых и скороспелых форм ячменя.
В результате использования биохимических маркеров и тестов при оценке и подборе исходного материала, проработке селекционных линий, их экологического испытания, селекционерам для дальнейшего селекционного отбора переданы 233 гибридные линии, созданные с участием тестированных на жаро-засухо-устойчивость в лабораторных условиях исходных форм.
ЗАКЛЮЧЕНИЕ
1. Установлено генетическое разнообразие коллекций генофонда Triticum aestivum L., идентифицировано наличие носителей 3-х аллелей (a, b и с) глютенинкодирующего локуса Glu А1 с преобладанием генотипов с «b» аллелью. По локусу Glu 1В в генофонде выявлены носители 9 аллелей (a, b, c, d, e, f, g, h, i), наиболее распространены носители аллелей “с”, и “b” (частота встречаемости 55,7% и 26,9% соответственно). Локус Glu D1 представлен в коллекциях 4-мя аллелями, из которых наиболее распространены “d” и “e” (встречаются с частотой 35,5% и 30,5%, соответственно). 20% образцов озимой пшеницы регионального питомника стран ЦАЗ и 19,9% Восточно-Европейского питомника имеют пшенично-ржаную транслокацию 1BL/ 1RS. Генетическое разнообразие (H) изученных коллекций варьирует от 0,28 у староместных сортов Западного Памира до 0,64 - коллекция образцов озимой пшеницы из США.
Показано, что селекционерами Казахстана используется 62,5% имеющегося в генофонде разнообразия генотипов по составу ВМСГ, для селекции на высокую урожайность, устойчивость к болезням в последнее время привлекается материал с пшенично-ржаной транслокацией.
2. Генофонд Triticum durum также разнообразен по составу ВМСГ, контролируемых локусом Glu - A1. Наряду с аллелями «a,» и «с», наиболее распространенными в коллекциях, (59,8% и 23,5% соответственно), выявлены образцы с редкими вариантами высокомолекулярных субъединиц глютенина 1* и 2*. Последняя не идентична ВМСГ 2* мягкой пшеницы. По локусу Glu В1 твердой пшеницы выявлены носители 5 аллельных вариантов (d, b, e, f, h), ответственных за биосинтез субъединиц: 6+8; 7+8; 20; 13+16; 14+15, соответственно. Генотипы с субъединицами 7+8 наиболее распространены и составляют 59,1% в генофонде.
На долю носителей субъединичной пары 6+8, сопряженной с высоким качеством макарон, приходится 14,8% образцов. Более 50% образцов коллекции характеризуется присутствием в составе глютенинов низкомолекулярных глютенинов НМГ-2, кодируемых локусом Glu-B3, более предпочтительных при отборе селекционного материала на качество.
Из всего набора изученных сортообразцов наиболее высоким уровнем генетического разнообразия выделяется коллекционный фонд отдела зерновых культур КазНИИЗиР и коллекция яровой твердой пшеницы Шалкарской ОС (0,54), наименее выражен этот показатель у регионального питомника стран ЦАЗ (0,39).
3. Выявлены новые, редкие высокомолекулярные субъединицы глютенина, эффективные для отбора ценных форм мягкой пшеницы: “a”, “b”, “c” и “d”; субъединичные пары глютенинов 5**+12; 0+ 9 мягкой пшеницы, установлен их генетический контроль и сопряженность с показателями урожайности и качества. Получены ценные гибридные популяции и выделены линии для расширения генетической основы донорного материала с целью создания новых сортов пшеницы.
4. Разработан каталог, включающий 30 вариантов блоков компонентов гордеина локуса Hrd A, 36 вариантов блоков компонентов гордеина локуса Hrd В, 13 вариантов блоков компонентов гордеина локуса Hrd F, которые кроме нумерации варианта аллели содержат информацию по ОЭП составляющих их компонентов в пределах a-, b - и w- белковых зон. Каталог позволяет вести запись белковых формул двумя способами: с указанием вариантов блоков компонентов гордеина и по позициям компонентов в пределах субфракций. На его основе составлены гордеиновые формулы 757 образцов генофонда ячменя, документированные в нашем изданном труде «Каталог генофонда ячменя по спектру гордеина» ( М, , 2007 г.).
5. В генофонде ячменя выявлены носиаллелей локуса Hrd A , с частотой встречаемости от 36,9% до 0,1%, 33 аллелей локуса Hrd B, с варьированием от 22,2% до 0,1% и 12 аллелей локуса Hrd F, частота которых колеблется от 20,9% до 0,1% . Генетическое разнообразие коллекций варьирует от 0,52 до 0,88. Значительная часть генофонда ячменя (24,1%) полиморфна по гордеинкодирующим локусам, в том числе по всем 3 гордеинкодирующим локусам - 12,9% , по 2ум - 7,0% - и 4,2% - по 1 локусу. Выявлено 3 варианта ВМСГ ячменя: D1, D2 и D3. Наиболее распространены субъединицы D1 и D2, тогда как субъединица D 3 выявлена лишь в единичных образцах Сирийского происхождения.
6. Установлена неспецифическая, характерная для всех генотипов ячменя реакция на высокую температуру, дефицит влаги, совместное действие стрессовых факторов различной интенсивности и длительности воздействия, проявляющаяся в повы-шении уровня свободной аминокислоты – пролина. У жаростойких форм наблюдается более интенсивное накопление осмопротектора в проростках и стабильность относительной активности катодной изопероксидазы С100. На основании этих закономерностей подана заявка на получение патента РК «Способ двухступенчатой оценки жаростойкости зерновых», № гос. Регистрации 2007/0593.1.
7.Определен биохимический состав зерна скороспелых образцов ячменя и характер утилизации гордеина при прорастании. Установлено, что эндосперм скороспелых форм характеризуется высоким уровнем свободного пролина, обеспечивающим опережающее развитие генотипа. При этом наблюдается замедленный и более продолжительный распад запасных белков в прорастающем зерне скороспелых генотипов.
8.Определено преимущество образцов мягкой пшеницы с ВМСГ 2*, 1 (аллели “a” и”b” локуса Glu А1), 7+8, 7+9, 17+18, 14+15 (аллели “b”, “c”, “i”, “h” локуса Glu В1) и 5+10, 5**+12 (аллели “d” и “нов.” локуса Glu D1) по показателям зерна и муки: твердозерность и седиментационный тест. В группе образцов с указанными аллелями выявлена более тесная взаимосвязь между содержанием белка и показателем седиментации (варьирование от 0,56 до 0,63). У образцов с «null» аллелью локуса Glu A1, ВМСГ 7 и 6+8 локуса Glu B1 и 2+10; 2+12 локуса Glu D1 значение показателя седиментации варьирует от 29,5 до 41,1 мл. У носителей указанных аллелей твердозерность тесно сопряжена с седиментационным тестом (r от 0,87 до 0,92).
9. В селекции пшеницы на устойчивость к желтой ржавчине установлена специфичность ВМСГ изогенной линии сорта Aroona с эффективным в регионе ЦАЗ геном Yr15 и пшенично-ржаная транслокация сорта озимой пшеницы Шарора, маркированная блоком глиадина 1В3. На основании этого созданы высокоустойчивые к патогену перспективные линии мягкой пшеницы.
10. Выделены по продуктивности, длине вегетационного периода, жароустойчивости, и способности к длительному хранению в генофонде 34 линии 9 биотипов ячменя, которые использованы в создании ценных форм.
11. Разработаны новые подходы в проведении биологических анализов исходного и селекционного материала зерновых колосовых культур. Созданы и переданы на Госсортоиспытание в соавторстве, 2 сорта озимой мягкой и 1 сорт яровой твердой пшеницы. В старших звеньях селекционного процесса проходят испытание 4 перспективные линии ячменя, 3 устойчивые к желтой ржавчине линии озимой мягкой пшеницы. Созданы 233 жаро-засухоустойчивые линии ячменя, среди них линия 61/7, превысившая по урожайности на 9ц/га или 23% стандартный сорт Арна, 52 линии мягкой пшеницы с генами расоспецифической устойчивости к желтой ржавчине (Yr9 и Yr15), 45 линий пшеницы F3-F4 поколения с редкими аллелями глютенинкодирующих локусов.
Предложения для практической селекции и семеноводства
1. В целях идентификации и регистрации образцов генофонда, селекционного материала и оценки типичности семян, присущей для сортов зерновых колосовых культур, рекомендуются разработанные серии сортов дифференциаторов.
2. При подборе исходного материала для программы гибридизации рекомендуется использовать генетико-биохимические формулы запасных белков, представленные в изданиях: «Каталог генофонда ячменя по спектру гордеина» (, , 2007 г.) и «Генетическое разнообразие коллекционных образцов генофонда твердой пшеницы НПЦЗиР по составу высокомолекулярных и низкомолекулярных глютенинов» (, и др., 2008 г.).
3. Для селекции на жаро-засухоустойчивость и скороспелость предлагается разработанный и внедренный в селекционный процесс комплекс биологических тестов, а также жароустойчивые и скороспелые биотипы ячменя и созданные с их участием 233 константные линии.
4. Для создания ржавчиноустойчивых сортов озимой пшеницы предлагаются устойчивые и иммунные к возбудителям желтой ржавчины гибридные линии мягкой пшеницы с эффективными генами расоспецифической устойчивости Yr15 и Yr9, характеризующиеся комплексом хозяйственно-ценных признаков.
5. Оценку технологических свойств зерна и муки селекционных образцов пшеницы рекомендуется вести в пределах групп, предварительно дифференцированных по составу ВМСГ, с учетом сопряженности определенных показателей качества; Генотипы с субъединицами глютенина 2*, 1 - локуса Glu A1; 7+8, 7+9, 17+18, 14+15 локуса Glu В1; 5+10, 5**+12, вариант “a” локуса Glu D1, 14+15 локуса Glu В1 мягкой пшеницы и 2*; 1* локуса Glu А1 твердой пшеницы предпочтительны при отборе на качество зерна и муки.
Список опубликованных работ по теме диссертации
1 , Перуанский ячменя в идентификации сортов и биотипов // Вестник с-х. науки Казахстана.-1988.- №8.-С. 30-33.
2 Булатова показатели жаростойких форм ярового ячменя //Вестник с.-х. науки Казахстана.-1991.-№6.- С.37-41.
3 Абугалиева качества зерна пшеницы на основе белковых маркеров /Тезисы респ. науч.-практ. конф. посвящ. 35-летию Актюб. с.-х. оп. Ст «Пути увеличения и улучшения качества с.-х. продукции в Казахстане. Актюбинск. -1992.-С.60-61.
4 , , Новохатин между компонентами глиадина, субъединиц глютенина и качеством зерна // Доклады РАСХН.- 1993.-, №4-С.9-13.
5 Булатова показатели биотипов ярового ячменя // Мат. межд. сов. «Генбанк растений и его использование в селекции».-Алматы.-1995.- С171-174.
6 Булатова белки ячменя в изучении гибридной популяции // Сб. науч. тр. «Научная деятельность Казахского НИИ земледелия им. -60 лет». – КАЗНИИЗ.- 1996.- Алматы..- «Бастау».- С.121-126.
7 , Сариев изменчивость ярового ячменя и биохимический контроль ее направленности //Сборник научных трудов «Повышение эффективности селекции полевых культур.-Алматы: РНИ «Бастау».-1997.-С.22-29.
8 ,Булатова М. А., Карбозов высокомолекулярных глютенинов озимых пшениц //Вестник с.-х. науки Казахстана. -1997. -№6. -С.54-58.
9 Булатова образцов ярового ячменя генофонда КазНИИЗа по составу запасных белков // Вестник с.-х. науки Казахстана.-1997.-№10.- С.16-22.
10 Булатова генотипов ячменя к повышенным температурам на стадии проростков // Сб. науч. тр. «Генетические основы селекции зерновых культур»- Алматы.- «Бастау».- 1998.- С. 68-78.
11 Булатова субъединица глютенина 7 в коллекционных образцах мягкой пшеницы // Сб. науч. тр.«Генетические основы селекции зерновых культур.».- Алматы.- «Бастау».- 1998.- С.194-198.
12 , Булатова показатели урожайности ярового ячменя в условиях Западного Казахстана // Сб. науч. тр. «Физиология и биохимия – неотьемлемое звено селекции». - Алматы. -1998.-С. 37-42.
13 , , Булатова особенности коллекционных форм ячменя //Мат. межд. научно-теоретич. конф. «Стратегия земледелия и растениеводства на пороге ХХI века.».- Алматы. -1999.
14 Bulatova K. M. Free Proline Content in Barley Seeds – Predictive Index of Early Maturity // Proceed. of 8th International Barley Symposium.- 2000.-P.211.
15 Булатова К., Состав запасных белков яровой мягкой пшеницы Казахстанской и Таджикской селекции // Тез. докл. межд. конф. «Развитие горных регионов Центральной Азии в XXI веке».- 2001.-Хорог, Горный Бадахшан, Таджикистан.-С.38-39.
16 Absattarova A., S. Baboyev, K. Bulatova M. Karabayev, M. Koishibayev, A. Kohmetova, V. Kuklacheva, A. Morgunov, S. Rsaliev, A. Sarbayev, R. Urasaliev, M. Yessimbekova, C.R. Wellings. Improvement of yellow rust resistance in Kazakhstan and Uzbekistan through sub – regional cooperation //First Regional Yellow Rust Conference for Centaral & West Asia and North Africa.- 2001.- P.34-41.
17 , , Джиенбаева К Состав запасных белков озимой пшеницы регионального питомника ЦАЗ //Вестник региональной сети по внедрению сортов пшеницы и семеноводству. -2002.-№2.-C.70-74.
18 , , Джиенбаева -биохимические особенности яровой пшеницы в связи с засухоустойчивостью // Вестник науки Акмолинского аграрного университета им. С. Сейфуллина.-2002.- Т.3, №6.- С.117-123.
19 , Булатова чужеродного генетического материала на стабильность локуса GLU B1 пшеницы //Мат. первой центрально-азиатской конференции по пшенице, - Алматы.-2С.41.
20 Булатова В. И., Цыганков и оценка генетического разнообразия сортимента яровой мягкой пшеницы западноказахстанских экотипов по спектрам глютенина. //Сб. научн. тр., посвященных 10-летию независимости Казахстана и 45-летию Актюбинской СХОС. «Использование достижений аграрной науки в стабилизации сельскохозяйственного производства Казахстана.». - Актобе.- 2003г.- С.84-89.
21 Булатова показатели в селекции ячменя на засу-хоустойчивость.//Мат. 6-й межд.. Научн.-практ. конф. «Научное обеспечение устойчивого развития АПК Казахстана, Кыргызстана, Монголии, России, Таджикистана и Узбекистана.- Павлодар.- 2003.- С.52-53.
22 , Булатова научная школа по биохимическому обеспечению селекционных и семеноводческих исследований // КазНИИЗ – 70 лет: Сб. науч. тр. – Алматы.- 2004.-С. 90-105.
23 Б, Булатова 1В/1R транслокации в селекции озимой пшеницы на устойчивость к желтой ржавчине // Материалы межд. научн. конф. «Стратегия научного обеспечения АПК РК в отраслях земледелия, рас-тениеводства и садоводства. Реальность и перспективы».- Алматы.- 2004. - С.213-214.
24 , Аширбаева и оценка генетического разнообразия коллекционных образцов твердой пшеницы по спектрам глютенина // Вестник с.-х. науки Казахстана.- 2004.- №6. - С.50-52.
25 , , Қ., Қожабаев қ арпа сорттарының шаруашылыққа құнды және маркерлік белгілері бойынша сипаттамасы //Жаршы.- 2004.- № б.
26 , , Моргунов яровой пшеницы КАСИП как источник доноров высокого качества зерна и урожайности // Мат. межд. конф. «Развитие ключевых направлений с.-х. науки в Казахстане: селек., биотехн., генет. ресурсы. Растениеводство.» - Астана.- 2004.- С.
27 Ш, О первичном семеноводстве полиморфных сортов // Мат. межд. научн. конф. «Дост. аграр. науки в обл. землед., селекц. и растениев.». – Алматы.- 2004.-С. 104-107.
28 Булатова разнообразие озимой пшеницы регионального питомника ЦАЗ по составу запасных белков зерна //Вестник Павлодарского ГУ им. С. Торайгырова, серия Химико-биологическая.-2005.- №4.-С.14-24.
29 , , Булатова сорта пшеницы и ячменя в семеноводстве // Матер. межд. науч. конф. «Биологические основы селекции и генофонда растений». -2005 . – Алматы.- 2005.- С.10-12.
30 Булатова особенности скороспелых форм ячменя // Вестник Павлодарского ГУ им. С. Торайгырова.- 2006.- Серия Химико-биологическая, №2.-С. 50-58.
31 М, Жамбакин электрофоретического спектра высокомолекулярных субъединиц глютенина у дигаплоидной линии пшеницы // Ж. Биотехнология. Теория и практика.- 2006.- №3.-С.41-46.
32 , , Агрономическая и биохимическая характеристики линий ячменя ИКАРДА, выделившихся в условиях Юго-Востока Казахстана // Вестник с.-х. науки Казахстана.- 2006.- №6.-С.8-11.
33 , , Мукин популяций Аegilops cylindrica host. Алматинской области по составу высокомолекулярных глютенинов //Вестник науки Казахского государственного агротехнического университета им. С. Сейфуллина.- 2007.- №1 (44).- С.131-136.
34 , , Ж,
Л Генетическое разнообразие Аegilops Сylindrica host. по составу проламинов – запасных белков зерна //Вестник КазНУ, серия биологическая.- 2007.-№2 (32).-С.81-86.
35 , Каталог генофонда ячменя по спектру гордеина // Алматы:АО «Баспалар Үйі».-2007. – 48 с.
36 , Сариев. Б.С. Генетическое разнообразие генофонда ячменя НПЦЗиР по составу гордеинов // Мат.10-ой межд. конф. по научному обеспечению азиатских территорий. - Улан-Батор.- 2007. -С.130-131.
37 И. А Нурпеисов., М. А Есимбекова., Генетическое разнообразие коллекционных образцов генофонда твердой пшеницы НПЦЗиР по составу высокомолекулярных и низкомоле-кулярных глютенинов // Исследования и результаты.- 2007. - №3.-С.32-36.
38 Булатова генетических ресурсов ячменя НПЦЗиР по составу гордеинов //Вестник КазНУ.-2007.- №5 (35), серия биологическая.-С. 36-40.
39 Булатова разнообразие коллекционных образцов гено-фонда мягкой пшеницы НПЦЗиР по составу высокомолекулярных глютенинов //Вестник Павлодарского ГУ им. С Торайгырова, серия Химико-биологическая.- 2007.- №4.–С.37-47.
40 Булатова аллелей локусов, контролирующих биосинтез субъединицы глютенина 7 мягкой пшеницы и сопутствующих ей компонентов //«Вестник Карагандинского университета», серия «Биология. География. Медицина».- 2007.- №4 (48).-С.10-15.
41 , , Аширбаева генофонда пшеницы по составу высокомолекулярных и низкомолекулярных субъединиц глютенина // Алматы, «CopyLand», 2008.-38с.
42 Булатова сорта зерновых культур как объекты генофонда, селекции и семеноводства// Сб. статей III межд. научн. практ. конф. «Аграрная наука – сельскому хозяйству.- Барнаул.-2008.-С.214-218.
43 , , Баймуратов гордеинов в идентификации ценных селекционных линий ячменя конкурсного сортоиспытания // Сибирский вестник сельскохозяйственной науки.- 2008.- №12.- С.107-113.
44 , , Джиенбаева гена устойчивости к желтой ржавчине Yr 15 сортам озимой пшеницы Казахстана на основе использования белковых маркеров //Вестник государственного университета имени Шакарима, 2008.- №2.-С.220-228.
45 , , Джиенбаева маркеры в создании исходных форм озимой пшеницы с Yr9 геном устойчивости к ржавчине. // Труды XI Международной научно-практической конференции «Развитие АПК азиатских территорий» - Кемерово.-2008.-Т 1.-С.245-250.
46 , , Сарбаев рекомендации по использованию биохимических маркеров в оценке генотипов пшеницы, устойчивых к желтой ржавчине//Изд. компания «Асыл кiтап»с.
БОЛАТОВА КҮЛПАШ МАНСҰРҚЫЗЫ
Масақты дәнді дақылдар селекциясының биологиялық негіздері
06.01.05 - Селекция және тұқымшаруашылығы.
Тұжырым
Зерттеу обьектілері ретінде гендікқордың үлгілері, бидай және арпаның регенеранттық, изогенді, селекциялық линиялары алынды. (3 мың үлгіден аса).
Жұмыстың мақсаты – Бидай және арпа селекциясындағы өсімдіктерді биологиялық талдау элементтерін жете зерттеу және жетілдіру.
Зерттеу нәтижелері: Республикада шығарылған бидай сорттары мен коллекциялық үлгілерінің дән белоктарын электрофоретикалық талдау нәтижесінде D геномымен кодталған арнайы «y» типті суббірліктің 4 варианты және жаңа «х» варианты анықталды.
Гордеин блоктары варианттарының каталогы жасалынды, яғни блоктардың сандық идентификациясынан басқа оларда кездесетін компоненттердің қатынасты электрофоретикалық жылжымалылықтары бойынша сипаттамасы берілді. Каталогта Hrd А локусының гордеин компоненттері блогының 30 варианттарының, Hrd B локусы гордеин компоненттері блогының 36 варианттарының, Hrd F локусы бойынша гордеин компонент блогының 13 варианттарының тасымалдаушыларының атаулары мен баяндаулары енгізілген. Каталогты қолдану арқылы арпа гендікқорының 757 үлгісінің гордеиндік формулалары жасалды. Гендікқордағы анықталған гордеинкодтаушы локустар аллелдерінің кездесу жиілігі айтарлықтай ауытқыды және Hrd А локусы бойынша 36,9%-дан 0,1%-ға дейін, Hrd В локусы бойынша 22,2%-дан 0,1%-ға дейін және Hrd F локусы бойынша 20,9%-дан 0,1%-ға дейін өзгереді. Арпа гендікқорының коллекциясы үлгілерінің едәуір бөлігі (24,1%) гордеинкодтаушы локустары жағдайына байланысты, яғни оның ішінде 12,9%-ы барлық 3 гордеинкодтаушы локустары бойынша, 7,0% - 2-еу және 4,2% - 1 локус бойынша полиморфты болып табылды. ДДС-Na қатысуымен жүргізілген диск –электрофорез әдісі арқылы арпаның ГЖМС-нің Д1, Д2, Д3 деп белгіленген 3 варианты анықталды. Оның ішінде біршама кең таралғаны Д1, Д суббірліктері, ал Д3 суббірлігі небары жекелеген үлгілерде табылды. Бидайдың коллекциялық үлгілеріндегі глютеннің жоғарымолекулалы суббірліктері құрамының талдауы бойынша Glu A1 глютенинкодтаушы локусы 1,2* (a, b) және «null» аллель (с) суббірліктерін кодтаушы 3 аллельді болып табылды. Коллекциялық жинақтың көп бөлігінде сапалылыққа жоғары үлес қосатын 2*суббірлігінің биосинтезін бақылаушы b аллелін тасымалдаушылар саны көп. Зерттелген коллекцияда кездесетін Glu 1В локусы бойынша аллелдер саны біршама әркелкілікті көрсетеді. Берілген локус бойынша 9 аллель (a, b,c, d,e, f,g, h,i) тасымалдаушылары анықталды, оның ішінде «c», және «b» аллельдері едәуір кең тараған және гендікқорда 55,7% және 26,9% (сәйкес) жиілігімен кездеседі. Glu D1 локусы зерттелген материалдарда 4 аллельмен көрсетілді, оның ішінде “d”және “e” аллельдері едәуір кең тараған және (35,5% және 30,5% жиілігімен кездеседі сәйкес). Зерттелген коллекциялардың генетикалық әртүрлілігі (Н) 3 глютенинкодтаушы локустарда кездескен аллелдері саны мен жиілігі жағынан 0,28-ден 0,64 ке дейін ауытқыды. Тұтасымен алғанда, Қаз ЕӨШҒЗИ жұмсақ бидай гендікқорындағы зерттелген үлгілердің 70%-н астамы Glu А1 және Glu В1 локусы аллелдері және 50 %-н астамы Glu Д1 локусы аллельдері бойынша сапа жағынан бағалы тасымалдаушы болып табылады. Қатты бидай үлгілерінің ішінде Glu А1 локусының 4 аллельді варианттарын тасымалдаушылар табылды, “null” аллелі (ең жай-жылдам қозғалатын суббірліктердің болмауы) және ГЖМС 1 биосинтезін бақылаушы аллель, біршама кең тарағаны (59,8% және 23,5% сәйкес). Қатты бидай үлгілерінің ішінде Glu В1 локусы бойынша 6+8; 7+8; 20;13+16; 14+15 (сәйкес) суббірліктері биосинтезіне жауап беруші 5 аллельді варианттарының (d, b,e, f, h) тасымалдаушылары анықталды. Орталықтың қатты бидай гендікқорында 7+8 суббірлікті генотиптер едәуір кең тараған және 59,1%-ды құрайды, ал макаронның жоғары сапалылығын көрсететін 6+8 суббірлікті жұбын тасымалдаушы үлесіне үлгілердің 14,8%-ы жатады екен. Тұтасымен алғанда, қатты бидай гендікқоры үлгілерінің 80%-ы Glu В1 локусы аллелдердің сапасы қатысында бағалы тасымалдаушы болып табылады. Қор белоктар спектрларының ұқсастығы негізінде жаздық арпа сорттарының 9 гордеин-горденинді биотиптерінің 34 линиялары гендікқорда ұзақ сақтау мүмкіншілігі, ыстыққа төзімділігі, даму кезеңінің ұзақтығы, өнімділік элементтерінің көрінуі бойынша зерттелінді және көбейтілді, осылардың есебінен ерте пісушілік және құрғақшылыққа төзімді селекция үшін бағалы формалар бөлініп алынды. Жаздық арпаның ыстыққа төзімді түрлері жоғары температура стресс кезінде 12 күндік өскін кезеңіндегі ұлпаларында бос пролин деңгейі біршама қарқынды түрде жинақталады және ең жылдам қозғалатын катодты изопероксидазаның белсенділік қатынасы бойынша тұрақты (С100). Жаздық арпаның 12 күндік өскін кезеңіндегі бос пролин концентрациясы мен жекелеген катодты изопероксидазаның қатынасты белсенділігі қоршаған ортаның жоғары температураға төзімді түрлерін зертханалық жағдайда сұрыптап алу үшін пайдаланылды, «Дәнді дақылдардың ыстыққа төзімділігін екісатылы бағалау тәсілі» жаңалығына алдын-ала патент және ҚР патентін алу үшін мәлімдеме жасалды, тіркелген мемлекеттік № 000/0593.1. Дәндегі бос пролин деңгейі арпаның сорттары мен линияларында айтарлықтай ауытқиды: оның құрамы тез пісетін үлгілерде кеш пісетін түрлеріне қарағанда едәуір жоғары. Гордеиннің тұқым өнуі кезінде тез ажырауы кеш пісетін түрлерінде 3 сөткелік кезеңде, ал тез пісетін түрлерде – 2-5 сөткеге дейін кеш болады. Бидай және арпа селекциясындағы өсімдіктерді биологиялық талдау элементтерін жете зерттеу, жетілдіру және енгізу негізінде бірлесіп шығарған бидайдың 2сорты және жаздық қатты бидайдың 1 сорты МСС –қа берілді, қазіргі таңда селекциялық процестердің аға питомниктерінде суармалы және суарылмайтын егістіктерде арпаның 4 перспективті линиялары мен күздік жұмсақ бидайдың сары татқа төзімді 3 линияларына сынақтар жүргізіліп жатыр, ыстық-құрғақшылыққа төзімділігі бойынша арпаның перспективті 233 будандық линиялары мен сары татқа арнайы төзімді генді (Yr9 және Yr15) сары тат қоздырғыштарына төзімді және иммунды жұмсақ бидайдың 52 будандық линиялары, бидайдың глютенинкодтаушы локустарының сирек кездесетін аллельді F3-F4 ұрпақтарының 45 линиялары селекционерлерге берілді.
BULATOVA KULPASH MANSUROVNA
Biological bases of cereals breeding
Abstract of a dissertation for the degree of Doctor of Biological Sciences on specialty 06.01.05 – Breeding and Seedgrowing.
Summary
Objects of investigation were samples of genefund, regenerated, isogenical and breeding lines of wheat and barley (more then 3 th. samples).
Aim of work: to develop and improve the elements of plant’s biological analyses in wheat and barley breeding process.
Results of investigation: In the results of electrophoretic analyses of seeds protein of wheat collection samples and varieties, cultivated in the Republic 4 variants of specific high molecular weight glutenin “y” type and new variant of “x” type subunits under the control of D genome were revealed. The catalog of hordein component blocks, in which besides number of blocs are given their characteristics on relative electrophoretic mobility and position of cooperated bands in a-, b - and w- fractions was revealed. Catalog includes descriptions of 30 blocks variants of hordein components of Hrd A locus, 36 of ones of Hrd B locus, 13 blocks of Hrd F locus and names of standart samples with definite block. According to this catalog hordein formulas of 757 genefund samples were constructed. Frequency of hordein coding loci alleles in genefund considerably varied and ranges from 36.9%to 0.1% for Hrd A locus, from22.2% to 0.1% for Hrd B locus and from 20.9% to 0.1% for the Hrd F locus.
Considerable part of barley genefund collections samples (24.1%) is polymorphic on the state of hordein coding loci, 12.9% - on all 3 loci, 7.0% - on two and 4.2% -on 1 the disc-electrophoresis method with DDS-Na 3 variants of HMWGS of barley, termed as D1,D2 and D3 were revealed. Most widespread subunits are D1 and D2, subunit D3 was detected only for individuals.
Analyses of HMWGS of wheat collections samples show, that glutenin coding locus Glu A1 is represented by the 3 alleles, controlling subunits 1, 2* (a, b) and silencing allele (c). Most part of collection sites has allelic variant “b”, responsible for the biosynthesis of subunit 2*, high ranged on grain quality. Locus GluB1 more variable on alleles number in the studied collections. In this locus were revealed 9 alleles (a, b,c, d,e, f,g, h,i) from which alleles “c” and “b” more distributed with frequency 55.7% and 26.9% accordingly. Genetic diversity (H) of investigated collections on the number and frequency of 3 glutenin coding alleles varied from 0.28 to 0.64. In a whole, more than 70% of bread wheat genefund material are value on quality according to the alleles of Glu A1 and Glu B1 loci, more than 50% - according to locus Glu D1 alleles.
Among the samples of durum wheat were revealed carriers of 4 allelic variants of locus Glu A1, from them “null” allele (absence of low mobility subunits) and allele, controlled biosynthesis of HMWGS 1 are most widespread (59.8% and 23.8%, accordingly).
For loci Glu В1 among the samples of durum wheat carriers of 5 allelic variants (d, b, e, f, h), responsible for the biosynthesis of subunits 6+8; 7+8; 20; 13+16; 14+15, accordingly, are revealed. In the durum wheat genefund of KazSRIA&PG genotypes with subunits 7+8 more distributed and form 59,1%, the part of carriers subunit pair 6+8, coupled with high pasta quality consist 14,8% samples. In a whole, more then 80% of durum wheat genefund samples are the carriers of value according to quality alleles of Glu В1 locus.
On the base of storage protein spectrum similarity were selected, multiplicated and studied on the productivity elements, length of vegetative period, heat resistance and ability to the long-term conservation in genefund 34 lines of 9 spring barley hordein biotypes, including value forms for the early maturing and drought resistance breeding.
Heat resistant spring barley forms more intensive accumulate in the 12 days old seedlings tissue free amino-acid praline and are stable on the relative activity of fast moving catod isoperoxidase (С100).
Concentration of free proline and relative activity of some catod isoperoxidazes of 12 day old spring barley were used as a criterium of laboratory selection of forms resistant to high temperature of environment, the application for reception of KR patent « the way of two-level estimation of cereals heat resistance» is submitted, № of state registration 2007/0593.1. The level of free praline in grain of varieties and lines considarebly varied: it’s content is higher for early maturing samples comparing with late ripening forms. Active degradation of hordein during late ripening form seeds germination come on the third day, for early maturing – on 2-5 days later.
On the base of development, improving and introducing the elements of plant biological analyses in to the breeding process of wheat and barley to the state variety testing (SVT) passed in co-authors 2 varieties of winter bread wheat and 1 variety of spring durum wheat, in upper chains of breeding processes at present there are tested 4 perspective for cultivation on bogar lands and irrigated conditions barley lines, 3 resistant to yellow rust lines of winter wheat, to breeders passed 233 hybrid lines of barley, perspective on heat-drought resistance, 52 resistant and immune to the pathogen of yellow rust hybrid lines with genes of specific resistance Yr9 и Yr15, 45 wheat lines F3-F4 generations with rare alleles of glutenin coding loci.
