
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Рис. 3. Соотношение величины коэффициентов вертикальной и горизонтальной деформации листовой ткани при дегидратации
Согласно проведенным расчетам, вертикальная деформация, связанная с изменениями тургора, у влаголюбивых видов составляет 84-94 %, у мезофитов - 70-87%, у ксеромезофитов - 94-96%, у ястребинки волосистой данный показатель составляет 72% от общего значения деформации листовой ткани при обезвоживании. Горизонтальная деформация обусловленная изменением тургора, у сныти обыкновенной составляет -70%, у гравилата речного - 63%, у герани - 59%, у чисто%, у земляниники - 83%, у коровяка -64%, у короставника -50%, у ястребинки волосистой -11%.
Эластичность листовой ткани в горизонтальном направлении также является неоднородной. Для всех исследованных растений наблюдается большая величина коэффициентов деформации в направлении поперек сосудов. Это свидетельствует о том, что эластичность ассимиляционной ткани в направлении вдоль сосудов ниже таковой в поперечном направлении. Уменьшение эластичности ассимиляционной ткани в направлении вдоль сосудов объясняется, вероятно, тем, что величина уменьшения размеров поверхности помимо эластичности живых клеток паренхимы и сосудов флоэмы зависит от степени жесткости клеток, составляющих сосуды ксилемы и механическую ткань.
Особенности изменения тургора у исследованных нами растений характеризуют следующий данные. При изменении влажности ткани листа на 1% наибольшие изменения (толщины) тургора наблюдаются у сныти - 3,31%, у гравилата -3,63%, у герани - 2,71%. У мезоморфных растений эта величина составляет: у василька лугового - 2,12%, у розги - 2,08%, у чисто,75%. Для ксеромезоморфной группы растений отмечаются меньшие изменения тургора ткани при изменении ее влажности на 1%: у короставника лугового эта величина составляет 1,92%, у коровяка - 1,57%, у мезоксероморфного вида (ястребинки волосистой) - 1,25% от диапазона изменения толщины листа при изменении состояния растительной ткани от максимального тургора до состояния плазмолиза. Эти данные показывают, что величина изменения тургора у влаголюбивых видов выше по сравнению с ксеромезофитами.
Исходя из представленных данных, во-первых, было бы логично ожидать, увеличения эластичности листовой ткани в ряду гигрофиты-мезофиты-ксерофиты. Однако, как показано выше, однозначной зависимости “показателя жесткости” в ряду исследованных растений не установлено. Данное несоответствие может быть связано с наличием различного характера зависимости тургора от объема клеток, находящихся в составе ткани. Последнее, по нашему предположению, обусловлено особенностями морфоструктуры листьев исследуемых растений. На схемах, часто используемых различными авторами [3,5,16] для выяснения соотношения между сосущей силой клетки, осмотическим и тургорным давлением, зависимость тургора от объема, как правило, считается линейной. Однако такое имеет место только при идеальной эластичности, что практически не наблюдается. В действительности зависимость тургора от объема клетки выражается кривыми, изображенными на рис 2.
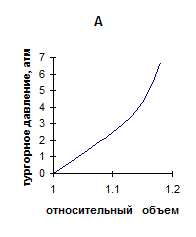
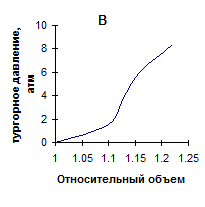
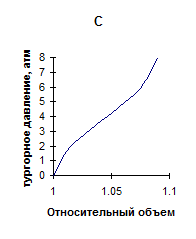
Рис. 2. Кривые, выражающие зависимость тургорного давления от объема
Кривая А характеризует ткань, имеющую высокие возвращающие силы, которые нарастают по мере увеличения деформации. Она характерна, например, для клеток окруженных тканью, обладающей большим сопротивлением эластическому растяжению (специальные клетки эпидермиса листа, или в коре, окружающей стебель). Кривая В характеризует клетки, стенки которых могут при натяжении истончаться по мере увеличения объема. И может наступить момент, когда клетка потеряв способность сопротивляться растяжению разрывается. Кривая С характеризует клетки, на динамике эластического растяжения которых кроме изменения тургора сказываются другие физиологические процессы. Например, если завядшие клетки поместить в воду, то резко уменьшится сосущая сила. Клетка быстро увеличится в объеме. Но за столь короткое время не успеет наступить пластическое растяжение клеточной оболочки, и по мере приближения к состоянию полного тургора будет наблюдаться уменьшение растяжения [5].
Как установлено нами ранее [17], у исследованных растений чаще всего наблюдается зависимость тургора от объема типа А и С. Растения, относящиеся к различным экологическим группам, могут иметь подобное морфологическое строение и близкие значения “показателя жесткости”. Они характеризуются одинаковым наклоном сигмоидальных кривых, отражающих зависимость влагосодержания от толщины (объема) листа. У таких растений наблюдается сходство кинетики тургора листовой ткани
Во-вторых, было бы логичным ожидать, что в ряду гигрофиты-мезофиты-ксерофиты должна наблюдаться подобная закономерность для количества воды, теряемой тканью при изменении ее толщины на единицу. Однако, максимальное количество воды, теряемое тканью при изменении толщины листовой пластинки на единицу (один микрон), у двукисточника составляет ) 0.244 мг/мкм, у сныти - 0.704, у гравилата - 0.430, у герани - 0.380, у чисто, у одуванчика - 0.260, у розги - 0.271, и у василька - 0.214, у земляники 0.490, у коровяка - 0.360, у короставника 0.411, у ястребинки - 0.600 мг/мкм. Эти данные отражают потери влаги при одинаковых изменениях тургора. Количество воды, теряемое тканью при изменении тургора на одну и туже величину, приблизительно одинаково у растений, имеющих близкие величины “показателя жесткости” (двукисточник, чистотел, розга, одуванчик), (герань, коровяк), (гравилат, земляника, короставник), (сныть, ястребинка).
Таким образом, растения, имеющие близкие значения “показателя жесткости”, при изменении тургора на одну и туже величину теряют одинаковое количество влаги. Однако изменения толщины (тургора) у них происходят с различной скоростью.
Так, при температуре 250 С, относительной влажности 70 %, на рассеянном свету начальное завядание (частичная потеря тургора) наступает у гигромезофитов и мезофитов через 35-45 минут, а у ксеромезофитов через 50-70 минут после начала дегидратации. Устойчивое завядание наблюдается у гигрофитов через 45-60, у мезофитов - через 55-80, у ксерофитов - через 80-100 минут после начала обезвоживания. Аналогично у гигроморфных растений плазмолиз наступает через 80-120 минут, у мезоморфных - через 115-150 минут, у ксеромезоморфных - через 130-185 минут (у ястребинки волосистой - через 155-220 мин.).
Вопрос о способах расчета скорости водоотдачи растительной тканью до настоящего времени также остается открытым. Дело в том, что в начале сушки изменения толщины и массы листа подчиняются линейному закону, а на втором этапе кривая сушки и толщины имеют нелинейный характер. Различные авторы для расчета скорости водоотдачи на нелинейном участке предлагают использовать эмпирические формулы. Точность получаемых по ним результатов неоднозначна для растений различных видов и условий обезвоживания. Основываясь на теории расчета скорости водоотдачи, изложенной в работах [20-22], мы рассчитывали скорость выхода воды в диапазоне бесстресовых (линейный участок кривой сушки) и стрессиндуцированных изменений тургора (нелинейный участок). На этапе бесстрессовых изменений тургора скорость выхода воды значительно выше таковой при завядании растений. Скорость выхода воды из клеток мезофилла на этапе стресиндуцированных изменений тургора зависит от вида растения и отрицательно коррелирует с величиной коэффициента жесткости ассимиляционной ткани. В диапазоне бесстресовых изменений тургора скорость выхода воды в большей степени определяется условиями дегидратации, чем видом дегидратируемой ткани.
Исследованные нами поля рассеяния корреляции между толщиной и влагосодержанием различных листьев одного и того же растения позволяют констатировать отсутствие прямой зависимости между толщиной листьев и их влагосодержанием. В тоже время, для каждого отдельного листа между толщиной и содержанием в нем воды существует определенная зависимость, графически выражающаяся сигмоидальной кривой. Данный факт указывает на то, что толщина листа (тургор) определяется комплексом показателей, зависящих не только от содержания воды, но и от степени эластичности и параметров морфоструктуры листьев (объема и формы межклетников, числа слоев столбчатой паренхимы и др.). Последнее детерминируется рядом факторов, среди которых следует выделить стадию онтогенеза листа, месторасположение его на стебле, взаимоотношением растений в ценозе.
На рис 1. приведены данные о величине и диапазоне вариации максимальной толщины листьев различных растений. Толщина листа в состоянии максимального тургора варьирует у двукисточника в диапазоне мкм; у чистомкм; у сныти обыкновенной - мкм; у земляники - мкм; у герани - мкм; у гравилата - мкм; у розги - мкм; у одуванчика - мкм; у коровяка - мкм; у короставника - мкм; у василька - мкм; у ястребинки - мкм.
Как видно из рисунка, по мере ксерофитизации видов увеличивается максимальная толщина листа, а также возрастает диапазон изменения этого показателя. Его средние значения отображены в виде линии.
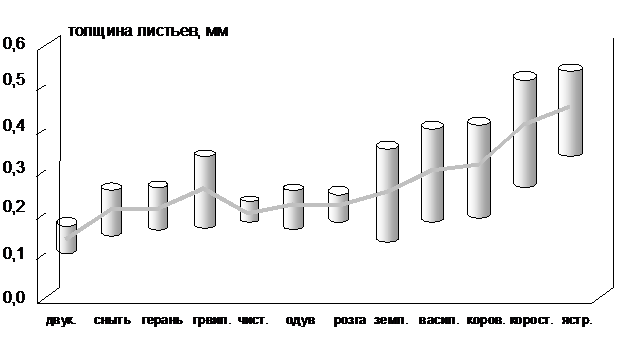
Рис. 1. Вариация максимальной толщины листьев растений.
Полученные данные позволяют констатировать, что в одинаковых условиях среды, чем влаголюбивее растение, тем значительнее деформация его ткани в ответ на изменение влажности. У ксеромезоморфных видов, с одной стороны - большая абсолютная величина толщины листа, а с другой - в ответ на экзогенную нагрузку: амплитуда колебания толщины у них значительнее, чем у влаголюбивых видов.
Таблица 4
Влагосодержание и эластические свойства дикорастущих растений. (Vmt - вариация влагосодержания в состоянии максимального тургора, Vkt - вариация объема воды, связанного с изменениями тургора, Vnkt - вариация объема воды, не связанного с изменениями тургора в % от объема листа при максимальном тургоре, a - Показатель жесткости, b - отношение объемов воды связанного и не связанного с изменениями тургора, dD - изменение толщины листа при дегидратации)
Вид растения | Год исследо-вания | Vmt | Vkt | Vnkt | dD | a | b |
Сныть обыкновенная | III | 46-74 | 20-43 | 14-45 | 19-65 | 0.87-4.2 | 0.33-3.0 |
среднее значение | 62 | 31 | 31 | 35 | 2.24 | 1.27 | |
диапазон изменения | II | 67-75 | 32-54 | 16-37 | 35-54 | 1.97-3.56 | 0.87-3.3 |
среднее значение | 72 | 47 | 25 | 41 | 2.50 | 2.1 | |
диапазон изменения | I | 40-75 | 17-58 | 14-39 | 33-51 | 0.88-3.29 | 0.71-3.3 |
среднее значение | 61 | 30 | 31 | 42 | 2.66 | 1.3 | |
Герань лесная | III | 39-69 | 32-61 | 6-18 | 29-74 | 1.54-6.11 | 1.65-5.93 |
среднее значение | 52 | 41 | 10 | 53 | 3.79 | 3.5 | |
диапазон изменения | II | 49-79 | 28-58 | 10-34 | 22-61 | 1.16-4.62 | 0.80-6.33 |
среднее значение | 62 | 40 | 22 | 46 | 2.92 | 2.16 | |
диапазон изменения | I | 52-67 | 10-29 | 7-39 | 40-70 | 2.37-5.52 | 0.77-1.47 |
среднее значение | 50 | 24 | 26 | 56 | 3.88 | 1.03 | |
Коровяк медвежье ухо | III | 42-88 | 40-78 | 10-23 | 20-77 | 1.03-9.67 | 1.78-7.67 |
среднее значение | 70 | 55 | 18 | 43 | 3.58 | 4.58 | |
диапазон изменения | II | 52-78 | 32-72 | 5-26 | 44-55 | 2.66-3.69 | 1.72-14.0 |
среднее значение | 68 | 49 | 19 | 49 | 3.09 | 4.02 | |
диапазон изменения | I | 33-53 | 21-42 | 17-37 | 52-56 | 3.38-3.84 | 1.04-2.19 |
среднее значение | 43 | 31 | 27 | 54 | 3.60 | 1.61 | |
Короставник полевой | III | 53-77 | 21-50 | 17-37 | 25-63 | 1.31-4.57 | 0.68-3.84 |
среднее значение | 71 | 44 | 27 | 41 | 2.50 | 1.91 | |
диапазон изменения | II | 65-76 | 45-67 | 6-23 | 24-64 | 1.24-4.65 | 1.9-10.3 |
среднее значение | 70 | 56 | 13 | 48 | 3.15 | 5.15 | |
диапазон изменения | I | 43-65 | 28-42 | 15-46 | 28-67 | 1.52-5.16 | 0.80-2.30 |
среднее значение | 62 | 34 | 29 | 53 | 3.64 | 1.36 | |
Ястребинка | III | 29-82 | 23-73 | 5-27 | 27-76 | 1.44-6.56 | 1.26-7.49 |
среднее значение | 67 | 47 | 16 | 46 | 3.13 | 4.67 | |
диапазон изменения | II | 52-73 | 41-59 | 5-11 | 11-64 | 0.59-5.79 | 3.59-9.50 |
среднее значение | 60 | 51 | 9 | 34 | 2.33 | 6.43 | |
диапазон изменения | I | 45-77 | 24-44 | 21-47 | 23-46 | 1.18-4.83 | 0.68-2.19 |
среднее значение | 63 | 31 | 32 | 41 | 2.52 | 1.08 |
Таким образом, тургор листа является показателем, отражающим степень адаптированности растений к конкретным экологическим условиям существования. В ряду гигрофиты - мезофиты - ксерофиты наблюдается тенденция увеличения толщины листьев и уменьшения диапазона изменения толщины в ответ на экзогенную нагрузку. Для растительной ткани, так же как и для клетки установлена анизатропность эластических свойств. Способность листа к сокращению в продольном направлении (вдоль градиента толщины) значительно превышает таковую в горизонтальном направлении (вдоль градиента поверхности). Коэффициенты продольной и поперечной деформации листовой ткани при обезвоживании отражают неравномерность эластических свойств листовой ткани в горизонтальной плоскости. Величина деформации ткани листа при дегидратации зависит от величины коэффициента ее жесткости. Особенности эластических свойств листовой ткани тесно сопряжены с различным характером зависимости тургорного давления от объема и скорости изменения тургора. Эластичность ассимиляционной ткани, определяющая способность клеток к сжатию и расширению в горизонтальном направлении и особенно в направлении, перпендикулярном к испаряющей поверхности, в ответ на изменяющееся влагосодержание листовой ткани, является существенным элементом регуляции растениями их водного гомеостаза.
Резюме
Вводная составляющая растений тесным образом связана с колебаниями толщины (объема) листовой пластинки и параметрами, характеризующими упругие свойства растительной ткани. Колебания объема ткани листа являются одним из необходимых адаптивных элементов, обеспечивающих устойчивость растения к меняющимся факторам среды Скорость реакции ассимиляционной ткани на обезвоживание зависит от “показателя ее жесткости”.
Литература
1. Zimmerman U., Steudle E. Physical aspects of water relation of plant cells. // Adv. Bot. Res. (London.v. 6.- P. 45-117.
2. Parkhurst D. F. The adaptive significance of stomatal occurrence on one or both surfaces of leaves. // J. Ecol.- 1978.- v. 66, N 2.- P. 367-383.
3. Физиология растительной клетки. М., 197с.
4. , Справочник по физике. М. 1979. С.304
5. Вода и ее значение в жизни растений. М., 195с.
6. Steudle E., Zimmermann V., Luttge U. Effect of turgor pressure and cell size on the wall elasticity of plant cell. // Plant Physiol.- 1977.- v. 59.- P. 285-289.
7. Современные аспекты изучения водного режима растений. // Физиолог. и биохим. культ. растен.- 1976.- т.8, в. 6.- С. 563-573.
8. Carrier H. B. Water relations of root cell of Beta vulgaris, Amer. Jour. Bot., 1944., 31, - P. 378-387.
9. Neumann P. M. The role of cell wall adjustment in plant resistance to water deficits. Crop Sc., 1995, vol. 35., N 5. P. .
10. Heyn R. P. The physiology of cell elongation Bot. Rev. 1940. N 6, P. 515-574
11. Steudle E., Wieneke J. Ñhanges in Wwter Relations and Elastic Propeties of Appele Fruit Cells During Growth and Development // J. Amer. Soc. Hort. Sci.- 1985.- v. 110.- P. 284.
12. Zimmermann U., Steudle E. The hydraulic cjnductivity and volumetric elastic modulus of cells and isolated cеll walls of Nittella and Chara spp. Pressure and volume effects. // Aust. J. Plant Physiol.- 1975.- v. 2.- P. 1-12.
13. Мелещенко водного транспорта высшего растения и ее элементы. 2. Метод определения упругих свойств стенок одиночных и находящихся в составе тканей клеток растений. // Физиол. растен.- 1997.- т. 44, N 2.- С. 256-263.
14. , , Родионов мониторинга водообмена листьев растений. // Весцi АН Беларусi. Сер. бiял. навук.- 1996, № 4.- С. 38-42.
15. Шенников растений. М., 195с.
16. Полевой физиология растений М. 1989. С.26
17. Телюк параметров морфоструктуры и водного режима листьев растений различных экологических групп.// Диссертационная работа на соискание ученой степени кандидата биологических наук. 19с.
18. Heyes J. A., Sealey D. F. Textural changes during nectarine (Prunus persica) development and ripening. //Sc. hortic., 1996, Vol. 65, N1, P. 49-58.
19. Tazawa M., Kamiya N. Water permeability of a Characefn internodal cell with special reference to its polarity. // Aust. J. Biol. Sci.- 1966.- v. 19.- P. 399-410.
20. Лыков сушки. М., 196с.
21. О формах связи влаги с материалами в процессе сушки. // Всесоюзное научно-техническое совещание по сушке. Пленарные заседания. М., 1958.- С. 20-33.
22. , Коссек сушки кукурузы и других зерновых и измельченных материалов. // сб.: Тепло - массоперенос. Минск, 1966.- т.5.- С. 217-221.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 |
