


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
4.4 Списки видов планктона
В таблице видов фитопланктона приведены названия таксонов и указана синонимия согласно требованиям современной ботанической номенклатуры. Фитопланктон разделен на 8 групп, характеризующиеся таксономическим родством (Bacillariophyta, Chlorophycota, Chrysophyta, Cryptophycophyta, Pyrrophycophyta, Euglenophycota, Haptophyta, Prasinophyta). С учетов сино-нимов общее количество таксонов достигает 527. Для каждого вида микроводорослей дана ве-личина массы клетки, вычисленная методом геометрического подобия фигур (Кольцова, 1970; Кожова и др., 1978; Plinski et al., 1984). Приведена видовая, экологическая и фитогеогра-фическая характеристика (PG – фитогеографическая характеристика: A – аркто-бореальные ви-ды, B – бореальные виды, C – космополитные виды; EG – экологическая хара-ктеристика: O – океанические формы, N – неритические формы, P – панталассные формы, M – микрофито-бентос, F - пресноводные формы). Таксоны снабжены кодами международного кодификатора (NODC Taxonomic Code). Пример таблицы видов фитопланктона дан в приложении B1. Списки видов фитопланктона помещены в файл DATA\TAXA\TAXPHYTO. XLS.
Список таксонов зоопланктона (Приложение В2) Баренцева и Карского морей включает ~200 названий животных (вместе с синонимами этот список достигает 282 названия). Таблица, пред-ставляющая этот список, построена по следующему принципу. Весь зоопланктон разделен на группы, характеризующиеся таксономическим родством. Выделена большая группа одноклеточ-ного зоопланктона. Многоклеточный зоопланктон, представлен голопланктоном (Coelenterata, Crustacea, Gastropoda, Chaetognatha, Appendicularia, Ctenophora, Rotatoria,) и меропланктоном (пелагические личинки донных животных).
Пример таблицы видов зоопланктона дан в приложении B2. На CD-ROM списки видов зооплан-ктона помещены в файл DATA\TAXA\TAX_ZOO. XLS.
______________________________
5. КОНТРОЛЬ КАЧЕСТВА ГИДРОБИОЛОГИЧЕСКИХ ДАННЫХ
5.1 Физические и гидрохимические данные
Контроль физических, гидрохимических данных проводился согласно схеме обработки океаногра-фической информации, принятой в работах Conkright et al., 1998; Matishov et al., 1998.
При обработке информации за годы возникла необходимость объединения биологичес-ких и физических данных. Первичная информация представляли собой два массива с разной структурой организации данных. Физические данные были сгруппированы по рейсам. Для каждо-го рейса указано название судна и определены координаты каждой станции. Второй массив содержал весовые характеристики проб фитопланктона. Для каждой пробы был указан номер станции, на которой она отобрана и дата. Объединение этих двух массивов проводилось на осно-ве таблицы, устанавливающей соответствие между номерами и координатами станций.
5.2 Биологические данные
Необходимый этап контроля данных состоит в проверки того, что значение параметра находится в допустимых пределах. Существует литература, содержащая диапазоны изменения океанографи-ческих характеристик для различных районов Баренцева моря (Matishov et al., 1998). Нам неиз-вестны работы, содержащие информацию о допустимых диапазонах изменения планктона для Баренцева морях. В настоящем разделе рассмотрены некоторые закономерности развития планктона и формулируются критерии контроля биологических данных.
Фитопланктон
В работах (Дружков, Макаревич, 1991; Дружков и др., 1997; Роухияйнен, 1967; Рыжов, 1985; Druzhkov, Makarevich, 1999) обсуждается схема функционирования фитопланктонного сообщест-ва (сукцессионная система) южной части Баренцева моря. Структура сукцессионных систем других районов Баренцева моря в целом тождественна той, которая рассмотрена в настоящем разделе. Отличия состоят во времени наступления вспышки цветения фитопланктона и её продолжительности.
Весна. Середина марта - начало июня
Начало весенней активности фитопланктона связано с появлением в прибрежной пелагиали во второй половине марта ранневесенних форм диатомовых: Thalassiosira hyalina (Grun.) Gran, T. cf. gravida Cl., Navicula pelagica Cl., N. septentrionalis (Grun.) Gran, Nitzschia grunowii Hasle, Amphora hyperborea (Grun.). Численность клеток в этот период невелика и может колебаться в зависи-мости от видового состава от нескольких десятков до нескольких сотен кл./л..
Биомасса фитопланктона достигает максимума во второй половине апреля. Максимальный уровень биомассы сохраняется в течение нескольких дней. Численность фитопланктона в период раннего цветения колеблется от нескольких сот тысяч до 2 млн кл./л (по неопубликованным дан-ным до 12 млн кл./л), а биомасса от 1 до 3 мг/л. В вертикальном распределении фитопланктона в этот период наблюдается концентрация основной массы в слое 0-10 м. Видами, формирующими первый максимум цветения фитопланктона являются: Thalassiosira cf. gravida Cl, T. nordenskioeldii Cl., Chaetoceros socialis Laud., C. furcellatus Bail., Navicula vanhoeffenii Gran. Кроме того, для этого периода в отдельные годы характерно интенсивное развитие золотистой водорос-ли Phaeocystis pouchetii (Hariot) Lagerh., которая может достигать значительных величин числен-ности и биомассы и активно участвовать в формировании весеннего максимума (наибольшие отмеченые численность и биомасса – 8 млн кл./л и 1.7 мг/л) (Дружков, Макаревич, 1989).
Лето. Конец июня – конец августа
В фитопланктонном сообществе летнего периода происходят значительные изменения. Идет процесс исчезновения весенних форм диатомовых. Отмечается повышение роли динофитовых микроводорослей, хотя их присутствие в пелагиали спорадическое. Наблюдается заметное за-мещение арктобореальных форм космополитными, а неритических форм – панталассными и океаническими. Основу доминирующего комплекса в этот сезон составляют диатомовые водо-росли Skeletonema costatum (Grev.) Cl., Leptocylindrus danicus Cl., L. minimus Gran, Chaetoceros decipiens Cl., C. laciniosus Schütt, а также динофитовые рода Protoperidinium. Максимальная численность клеток пелагических водорослей не превышает в этот сезон 20 тыс. в литре.
Осень. Середина сентября – начало ноября
В слое 0-25 м наблюдается максимальная плотность клеток. Доминирующие виды: динофитовые родов Ceratium, Dinophysis, Protoperidinium и диатомовые рода Chaetoceros. Численность клеток не превышает 2 тысяч в 1 литре. К началу декабря численность клеток не превышает тысячи
в 1 литре, биомасса менее - 5 мкг/л. В пелагиали наблюдается полное доминирование динофито-вых водорослей, а в качестве единственной активной группы фотосинтезирующих организмов остаются нанопланктонные флагелляты.
Зима. Середина ноября - середина марта
Фитопланктонное сообщество весь зимний период находится в стадии покоя. В пелагиали фитопланктон представлен в основном крупными океаническими динофитовыми водорослями космополитного и арктобореального происхождения. Концентрация колеблется от нескольких клеток до нескольких десятков клеток на 1 литр. Основу доминирующего комплекса составляют Ceratium longipes (Bail.) Gran, C. tripos (O. Müll.) Nitzsch, Dinophysis norvegica Clap. Et Lachm., Protoperidinium depressum (Bail.) Balech.
В таблице 2 приведены характеристики годового цикла развития фитопланктона Баренцева моря. Эта таблица определяет диапазон допустимых значений доминирующих видов фитопланктона для южной части Баренцева моря.
Таблица 2. Характеристики годового цикла развития фитопланктона Баренцева моря
Район: 74oN - Кольский полуостров
Время | Глубинаобитания (м) | Структура таксонов | Преобладающие виды | Численность (клеток/л) |
Весна | ||||
Середина марта - начало июня | 0-70 | N > O+P+M+F A > B+C | Phaeocystis pouchetii Thalassiosira gravida T. nordenskioeldii Nitzschia grunowii Chaetoceros socialis Navicula | 100 тысяч - 12 миллионов |
Лето | ||||
– конец августа | 0-50 | C ³ A+B N ³ P+O | Leptocylindrus danicus L. minimus Chaetoceros decipiens C. laciniosus Protoperidinium Skeletonema costatum | >100 тысяч |
Осень | ||||
Середина сентября – начало ноября | 0-25 | C > A; C > B O ³ P; O ³ N | Chaetoceros Ceratium Dinophysis Protoperidinium | > 2000 |
Зима | ||||
Начало ноября - середина марта | 0–дно | O > N O > P; C+A > B | Ceratium Protoperidinium | 1 |
Зоопланктон
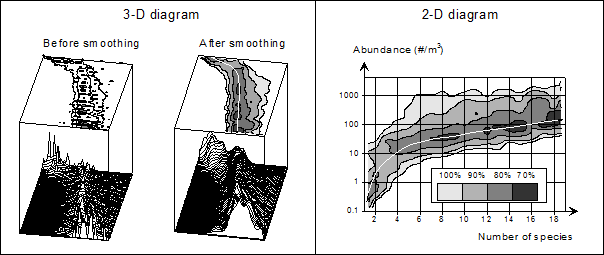
Наличие в базе данных более 9,000 проб позволяет рассмотреть зависимость между числом видов и количеством зоопланктона в пробе.
На рисунке 5 приведен график зависимости число видов в м3 – обилие зоопланктона в м3 для Баренцева и Карского морей (только голопланктон). Характер полученной зависимости хорошо согласуется с теоретическими кривыми (Magurran, 1988).
| |
|
| |
 | |
Рис. 5. Зоопланктон Баренцева и Карского морей: обилие - число видов
График (рис. 5), представлен в табличном виде для алгоритмизации контроля качества данных.
Количество видов (экз./м3) | 1 | 2-3 | 4-5 | 6-10 | 11-15 | 16-20 | >20 |
Минимальное обилие (экз./м3) | 0.1 | 1 | 3 | 10 | 12 | 14 | >15 |
Среднее обилие (экз./м3) | 1-75 | 76-200 | 201-260 | 261-350 | 351-400 | 401-450 | >450 |
Максимальное обилие (экз./м3) | 150 | 350 | 1500 | 2500 | 2550 | 2600 | >2600 |
______________________________
ВИЗУАЛИЗАЦИЯ ДАННЫХ
6.1 Физические характеристики
Процессы таяния льда, вертикальная структура вод и термические характеристики морской сре-ды определяют динамику развития планктона Карского и Баренцева морей. Настоящий разделе содержит информацию о положении кромки льда, вертикальной структуре вод, полях темпера-туры и солёности.
Лёд
На CD-ROM в разделе WWW\MAP\ICE приведены, карты характеризующие среднее положе-ние кромки льда для середины каждого месяца (Eastern-Western., 1984).
Температура и солёность
Процедура объективного анализа, используемая в настоящей работе для построения полей океанографических характеристик, в основном соответствует схеме, предложенной Barns (1973) и методам, рассмотренных в работе Levitus S., Boyed T. P. (1994). Дополнения алгоритма каса-лись учёта анизотропности океанографических полей Баренцева и Карского морей.
При расчетах полей распределения температуры на поверхности Баренцева и Карского морей летом принят радиус корреляции 250 км, зимой этот радиус уменьшается до 180 км. На глубине 100 м радиус на 35-40 % меньше, чем на поверхности. Значения температуры и солёности рассчитывались для сеточной области с шагом 20х20 км за три интервала времени: годы, годы, годы. Выбор таких интервалов обусловлен наличием данных по планктону, температуре и солености воды в пределах указанных лет. Для каждого интервала времени построены следующие карты:
Баренцево море – температура и солёность, поверхность и 100м, зима и лето.
Карское море - температура и солёность, поверхность и 100м, лето.
Зима = {январь, февраль, март, апрель}, лето = {июль, август, сентябрь}.
Карты представлены в Приложении D и на CD-ROM в формате HTML.
Океанографические данные, используемые для построения карт температуры и солёности, получены из баз данных WDC, Silver Spring и ММБИ.
Вертикальная структура вод Баренцева моря
Вопросам исследования вертикальной структуры вод Баренцева моря посвящено большое число работ. Установлено, что зимой температура (ТоС) и плотность (σ) воды незначительно изменя-ются с глубиной. Резкий градиент Т и σ наблюдается в слое 30-80 метров летом из-за повыше-ния температуры верхнего слоя воды. Наличие месячных климатических полей по температуре и солёности для Баренцева моря (Матишов и др., 1998) позволяют документировать годовой цикл изменения Т и σ в вертикальной плоскости. Алгоритм расчёта вертикального градиента Т и σ состоял из следующих этапов. а) На основе месячных климатических полей температуры и солёности рассчитаны климатические поля плотности для января, ... , декабря. Шаг сеточной области 10'x30'. в) Для каждого месяца построены поля, характеризующие разность значений температуры (ΔТ) и плотности (Δσ) на горизонтах 0 и 100 метров:
ΔТ = Т0м - Т100м; Δσ = σ0м - σ100м
с) Метод объективного анализа использовался для картирования значений ΔТ и Δσ.
На CD-ROM через систему HTML доступны график и карты, характеризующие годовой цикл из-менения значений ΔТ и Δσ. Полученные результаты свидетельствуют о наличие двух периодов времени, в течение которых сохраняется устойчивый режим вертикального градиента темпе-ратуры и плотности: летний и зимний р6ежимы. Протяженность зимнего режима с январь по апрель. В этот период значения ΔТ и Δσ достигают годового минимума. Протяженность летнего режима с июля по сентябрь. В этот период значения ΔТ и Δσ достигают годового максимума.
6.2 Биологические характеристики
Для описания состояния планктонных сообществ, обычно, используются поля (карты) распре-деления численности, биомассы, и числа видов планктона. В практике гидробиологических ис-следований также используются коэффициенты биоразнообразия. Причина повышения уровня разнообразия жизни - поступление в экосистему дополнительной энергии (Legendre, Demers, 1985). Источник дополнительной энергии определяется региональными особенностями иссле -
дуемого района океана. Например, в Карском море это может быть приток атлантических вод с севера или сток рек Оби и Енисея. В Баренцевом море - приток атлантических вод или пресная вода, поступающая при таянии льда (Тимофеев, 1988). Таким образом, поля распределения характеристик планктона (Приложение Е, F), не только документируют состояние планктонного сообщества, но и выступают как инструмент исследования водных масс морей Арктики.
В качестве коэффициента биоразнообразия использовался коэффициент Глиссона (Kgl):
Kgl=(Nt-1)/log(Ni)
где: Ni - количество экземпляров,
Nt - количество видов в пробе.
В базе данных содержится информация по зоопланктону Карского моря, собранная в 1936 году на судне Нерпа и в 1981 году на судне Дальние Зеленцы. Численность зоопланктона в 1936 го-ду определена в терминах редко, обычно, обильно и очень обильно. Численность зоопланктона в 1981 году определена в экз./м3. Для сравнения данных этих рейсов численность зоопланктона 1981 года представлены в тех же терминах, что и в 1936 году с использованием следующей шкалы (Дробышева и др., 1986):
редко = 1-10 экземпляров/м3
обычно = 11-100 экземпляров/м3
обильно = экземпляров/м3
очень обильно >1000 экземпляров/м3
В Приложениях Е и F приведены поля распределения характеристик планктона в вертикальной и горизонтальной плоскостях. В приложении E4 приведены графики изменения характеристик фитопланктона по трассе движения атомных ледоколов зимой из Баренцева в Карское море и обратно. Эти графики документируют состояние фитопланктона в районах, ранее недоступных для гидробиологических исследований в зимнее время. Весь графический материал доступен на CD-ROM через информационную систему HTML.
______________________________
7. ИЗМЕНЕНИЯ ПЛАНКТОННОГО СООБЩЕСТВА
Задача настоящего раздела – иллюстрировать возможности базы данных для документирования изменений планктонных сообществ Баренцева и Карского морей. Для этого выбраны две группы данных. Первая – относится к периоду потепления Арктики– годы (Fu et al., 1999). Вторая–относится к периоду с более суровыми условиями – после 1950 года (Fu et al., 1999). Все рисунки, на которые сделаны ссылки в настоящем разделе, находятся в Приложении G.
Фитопланктон. Баренцево море. Кольский разрез: 1921 vs. 1997
Данные: a) рейс судна Соколица, май 1921, 5 станций 16 проб по Кольскому разрезу; b) рейс судна Помор, май 1997, 7 станций 35 проб по Кольскому разрезу.
Характеристики: Численность фитопланктона, коэффициент биоразнообразия (коэффициент Глиссона), доли видов с различной биогеографической (число арктических видов) и экологичес-кой (число океанических видов) характеристиками. Графики изменения перечисленных показа-телей вдоль Кольского разреза приведены на рисунках G1, G5. Эти графики свидетельствуют о наличии существенных различий в структуре сообществ фитопланктона в 1921 и 1997 годах.
Вывод: Каждая из анализируемых характеристик указывает на то, что в мае 1921 года условия для развития фитопланктона были более благоприятные, чем в мае 1997 года.
Фитопланктон. Баренцево море:
Данные: Для анализа взяты данные за апрель-май 1921, 1957, 1985 и 1997 годов из района с центром в точке 71oN 33o30'E и радиусом 15 миль. Всего выбрано 8 станций, 37 проб.
Характеристики: Рассчитаны средние для апреля-мая значения численности клеток фи-топланктона под м2 и коэффициента биоразнообразия (коэффициент Глиссона) для 1921, 1957, 1985 и 1997 годов (рис. G2). Значения указанных параметров в 1921 году значительно большие по сравнению с остальными годами.
Вывод: Условия для развития фитопланктона в апрель-май 1921 года были более благопри-ятные, чем в аналогичные периоды 1957, 1985 и 1997 годов.
Зоопланктон. Карское море: 1936 vs. 1981
Данные: a) рейс судна Нерпа, август 1936 г., 38 станций 143 пробы в Карском море; б) рейс судна Дальние Зеленцы, август 1981 г., 24 станции 109 проб в Карском море.
Характеристики: Рассчитана встречаемость видов (количество видов в % от общего числа) зоопланктона, являющихся индикаторами арктических вод (рис. G3). Встречаемость видов-индикаторов арктических вод в 1981 заметно выше, чем в 1936.
Вывод: Климатические условия в Карском море в 1981 были более суровыми, чем в 1936 году.
Зоопланктон. Южная часть Баренцева моря:
Данные: Материалы 1630 станций, 7137 проб, собранные в 84 рейсах, в годы.
Характеристики: Построены графики, характеризующие изменения биомассы, численности, индекса биоразнообразия (коэффициент Глиссона) и аномалий температуры в период (рис. G6). Прослеживается тенденция снижения значений этих параметров от 1952 к 1959 году.
Вывод: годы характеризуются более благоприятными условиями для развития зо-опланктона, по сравнению с годами. Одна из возможных причин, обусловливающих это явление – положительные аномалии температуры в годы и отрицательные анома-лии температуры в годы (рис. G4).
Резюме
Приведенные примеры свидетельствуют о том, что в рассматриваемом районе Арктики в е годы наблюдались более благоприятные условия для развития планктона, по сравнению в ми годами. Этот вывод согласуется с существующими наблюдениями о резком потеплении Арктики в период х годов (Fu et al., 1999).
______________________________
8. СОДЕРЖАНИЕ CD-ROM
На CD-ROM помещены первичные данные, вспомогательные файлы, а также настоящий Атлас в формате HTML и doc (Microsoft WORD). HTML версия Атласа состоит из 8 разделов:
Документация. Раздел содержит текст Биологического атласа морей Арктики 2000: планктон Баренцева и Карского морей на русском и английском языках.
История. Представлен список работ по исследованию планктона Баренцева моря и карты распределения бентосных станций за периоды и годы.
Виды планктона. Приведены списки видов планктона морей Арктики. Предусмотрен поиск видов по алфавиту и по таксономическим группам. Для каждого вида фитопланктона даны его геогра-фическая и экологическая характеристика.
Фотогаллерея. В фотоальбом включены фотографии живых клеток и рисунки 50 доминантных видов фитопланктона морей Арктики, а также фотографии процесса сбора проб фитопланктона с борта атомного ледокола Советский Союз.
База данных. Приведены карты распределения данных. Предусмотрены средства для просмотра данных по каждому рейсу. На CD-ROM в разделе DATA\PRIMARY приведены данные 158 рейсов за период годы.
Морская среда. В раздел включены карты и графики, описывающие распределение различных характеристик фито - и зоопланктона, а также карты распределения температуры и солёности, положения кромки льда, а также характеристики вертикальной структуры вод Баренцева моря.
Изменения планктонного сообщества. Сравнивается структуры планктона 1930-х, 1950-х, 1990-х годов. Выявленные различия сопостовляются с изменением климата Арктики.
Авторы. Приводится список авторов с указанием их адресов, телефонов и электронной почты.
_____________________________
9. ЗАКЛЮЧЕНИЕ И БУДУЩИЕ РАБОТЫ
На примере зообентоса показано, что предложенный формат описания данных по блокам может быть использован для формализации широкого круга параметров.
Результаты сравнения структурных характеристик планктона показывают, что 20-30-е годы были более благоприятны для развития планктона по сравнению с 50-ми, 80-ми и 90-ми годами.
Показано, что внутригодовая изменчивость характеристик зоопланктона, собранного в период годов в прибрежной зоне Кольского полуострова, совпадала по фазе с флуктуациями аномалий температуры.
Пополнение базы данных и документирование изменений гидробиологических характеристик морей Арктики будут приоритетными направлениями наших будущих работ. Мы предполагаем усовершенствовать контроль качества гидробиологических данных и описание методов изме-рений.
______________________________
10. БИБЛИОГРАФИЯ
, , 1988. Гидролого-гидрохимические исследования. – В книге: Контроль экологической ситуации в районе опытно-промышленной плантации водорослей в губе Дальнезеленецкой. Апатиты, изд. Кольского филиала АН СССР, с. 7-23.
Баренцево море, 1990. Гидрометеорология и гидрохимия морей СССР. Том 1, вып. 1. Л., Гидрометеоиздат, 280 с.
, 1927. К методике обработки планктона (Новая камера для обработки зооплан-ктона). - Русский гидробиологический журнал, т. 6, вып. 8-10, с. 193-198.
, 1934. Инструкция по сбору и обработке материала по исследованию питания планктоноядных рыб. М., изд. ВНИРО, 15 с.
, 1938. К методике исследования планктона. - Зоологический журнал, т. 17, вып. 2, с. 373-380.
, 1940. К методике исследования планктона в море: Некоторые новые приборы для сбора планктона. - Зоологический журнал, т. 19, вып. 1, с. 172-182.
, , 1978. Потенциметрическое определение рН. – В книге: Методы гидрохимических исследований океана. М., Наука, с. 106-110.
, , 1989. Об объективном анализе океанографических полей по данным судовых съемок в Баренцевом море. - Труды ААНИИ, т.415, с.117-126.
, , 1992. Комплекс методов статистической обработки и объективного анализа данных натурных океанографических экспериментов. - Труды ААНИИ, т.426, с.7-19.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 |
