

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Высокогорный комплекс играет более заметную роль в сложении флоры Верховий, тогда как в Низовьях высокогорные виды составляют лишь 5,3 % (рис. 4). Из 62 дифференциальных видов Верховий 75,8 % относится к этому комплексу. Для него характерна самая высокая доля южносибирских видов – 18,3 %, что связано с наиболее четкими чертами автохтонности горной флоры Байкальской Сибири (Малышев, Пешкова, 1984).
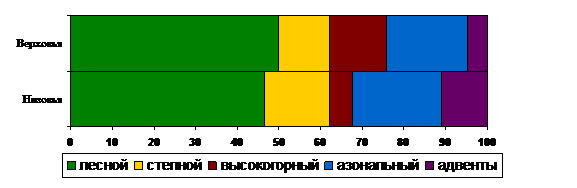 Рис. 4. Долевое участие флористических комплексов аборигенной фракции и адвентивных видов в сложении ЛФ бассейна Тойсука.
Рис. 4. Долевое участие флористических комплексов аборигенной фракции и адвентивных видов в сложении ЛФ бассейна Тойсука.
Наименее специфичным является азональный комплекс, охватывающий 20,2 % всего разнообразия бассейна. 85,5 % азонального комплекса – это широкоареальные растения, только 21 вид ограничен в своем распространении пределами Азии.
Разница участия флористических комплексов в аборигенных фракциях ЛФ колеблется от 2,3 до 8,5 % (см. рис. 4). Наименьшая амплитуда характерна для азонального комплекса (2,3 %), а наибольшая – для высокогорного (8,5 %).
Таким образом, флора бассейна Тойсука сформирована на основе лесных и азональных видов при заметном участии горных элементов в Верховьях и степных – в Низовьях. Особенности поясно-зональной структуры ЛФ определяются различиями в рельефе, следовательно, разным набором экотопов, а также неравномерной антропогенной нагрузкой на эти территории.
3.4. Эколого-биологическая структура флоры
3.4.1. Биоморфологический анализ флоры
При биоморфологическом анализе флоры Тойсука использованы две классические системы: К. Раункиера (Raunkiaer, 1905; 1934) и (1962, 1964).
По системе Раункиера, флора бассейна Тойсука представлена 5 типами жизненных форм (табл. 5). Криптофиты разделены на три подтипа. Спектры жизненных форм ЛФ различаются незначительно. Для флоры в целом характерно преобладание растений, переживающих неблагоприятный период либо в приземном слое (гемикриптофиты – 54,9 %), либо полностью уйдя из воздушной среды (криптофиты – 17,3 %), что обусловлено такими свойствами умеренно-холодного климата территории исследования, как достаточно суровые, продолжительные зимы и сравнительно короткий вегетационный период. Существенное участие терофитов во флоре (более 15 %) косвенно свидетельствует о нарушении растительного покрова. К примеру, группа адвентов, лучше всего представленная в Низовьях, более чем на 60 % сложена терофитами.
Таблица 5
Спектр жизненных форм флоры бассейна Тойсука по системе Раункиера
Жизненная форма | Флора в целом | Верховья | Низовья | |||
Число видов | % от общ. числа видов | Число видов | % от общ. числа видов | Число видов | % от общ. числа видов | |
Фанерофиты | 65 | 9,0 | 50 | 9,3 | 60 | 9,1 |
Хамефиты | 27 | 3,7 | 25 | 4,7 | 22 | 3,3 |
Гемикриптофиты | 397 | 54,9 | 310 | 57,8 | 354 | 53,6 |
Криптофиты: | 125 | 17,3 | 91 | 17,0 | 117 | 17,7 |
геофиты | 92 | 12,7 | 68 | 12,7 | 84 | 12,7 |
гелофиты | 24 | 3,6 | 18 | 3,4 | 24 | 3,3 |
гидрофиты | 9 | 1,2 | 5 | 0,9 | 9 | 1,4 |
Терофиты | 109 | 15,1 | 60 | 11,2 | 108 | 16,3 |
Всего | 723 | 100,0 | 536 | 100,0 | 661 | 100,0 |
Спектр жизненных форм по системе (табл. 6) типичен для бореальных флор (Юрцев, 1968; Толмачев, 1974) и показывает преобладание травянистых (88,8 %) над деревянистыми, а среди травянистых – многолетников над малолетниками. Полудревесные растения (Artemisia frigida, A. gmelinii и др.) представлены, в основном, в горно-степной группе и приурочены к скально-щебнистым местообитаниям и осыпям, что вызвано сходством условий этих экотопов с условиями аридных районов, для которых полукустарники характерны, и где наиболее ярко проявляется их адаптивный потенциал (Горшкова, 1966).
Группа травянистых поликарпиков, наиболее ярко отражающая почвенно-климатические условия района исследования, лидирует в степном (96 %), высокогорном (88 %) и лесном (80 %) комплексах.
Таблица 6
Биоморфологическая структура флоры бассейна р. Тойсук и его ЛФ
Биоморфологические группы | Флора в целом | Верховья | Низовья | |||
число видов | % от общ. числа видов | число видов | % от общ. числа видов | число видов | % от общ. числа видов | |
1. ДРЕВЕСНЫЕ РАСТЕНИЯ | 71 | 9,8 | 57 | 10,6 | 64 | 9,7 |
Деревья | 12 | 1,7 | 11 | 2,1 | 12 | 1,8 |
Кустарники | 50 | 6,9 | 37 | 6,9 | 45 | 6,8 |
Кустарнички | 9 | 1,2 | 9 | 1,7 | 7 | 1,1 |
2. ПОЛУДРЕВЕСНЫЕ РАСТЕНИЯ | 10 | 1,4 | 8 | 1,5 | 9 | 1,4 |
Полукустарники | 3 | 0,4 | 2 | 0,4 | 3 | 0,5 |
Полукустарнички | 7 | 1,0 | 6 | 1,1 | 6 | 0,9 |
3. НАЗЕМНЫЕ ТРАВЫ | 628 | 86,9 | 462 | 86,2 | 574 | 86,8 |
Поликарпические травы | 523 | 72,3 | 405 | 75,6 | 470 | 71,1 |
стержнекорневые | 66 | 9,1 | 43 | 8,0 | 59 | 8,9 |
короткокорневищные | 150 | 20,7 | 126 | 23,5 | 135 | 20,4 |
длиннокорневищные | 196 | 27,1 | 156 | 29,1 | 178 | 26,9 |
плотнокустовые дерновинные | 21 | 2,9 | 14 | 2,6 | 19 | 2,9 |
рыхлокустовые дерновинные | 42 | 5,8 | 35 | 6,5 | 35 | 5,3 |
клубнеобразующие | 17 | 2,4 | 7 | 1,3 | 16 | 2,4 |
кистекорневые | 12 | 1,7 | 10 | 1,9 | 12 | 1,8 |
корнеотпрысковые | 5 | 0,7 | 2 | 0,4 | 4 | 0,6 |
луковичные | 8 | 1,1 | 6 | 1,1 | 8 | 1,2 |
наземно-ползучие | 6 | 0,8 | 6 | 1,1 | 4 | 0,6 |
Монокарпические травы | 105 | 14,5 | 57 | 10,6 | 104 | 15,7 |
4. ВОДНЫЕ ТРАВЫ | 14 | 1,9 | 9 | 1,7 | 14 | 2,1 |
Итого травянистых | 642 | 88,8 | 471 | 87,9 | 588 | 89,0 |
Всего | 723 | 100,0 | 536 | 100,0 | 661 | 100,0 |
Среди травянистых многолетников длиннокорневищные растения наиболее многочисленная группа во всех флористических комплексах, кроме степного (рис. 5). Возможность произрастания значительного числа длиннокорневищных растений, предпочитающих местообитания с достаточно влажными рыхлыми и аэрируемыми почвами (Шенников, 1950), обеспечивается господством таежных лесов, в условиях которых формируются почвы с подобными характеристиками. Это подтверждается тем фактом, что среди длиннокорневищных растений только 8,7 % принадлежат ксерофитному ряду, тогда как среди короткокорневищных уже 14,6 % растений предпочитают более сухие местообитания.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
