

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Каждый вид занимает в биосфере Земли только ему присущее место – нишу, которое определяется совокупностью внешних абиотических условий и наличия необходимых для питания ресурсов. Потенциальная ниша – это такое максимальное пространство (численность), которое мог бы занять (иметь) вид, если бы ареал пригодных для его жизни условий совпадал бы с ареалом распространения доступных для него ресурсов. Поскольку в природе такое явление практически не встречается, мы имеем дело с фундаментальной нишей – пространством (численностью), заселяемым (реализуемой) видом без биотических ограничений. Приведенные в скобках оговорки не случайны. Пространственные характеристики экологической ниши, часто используемые для ее определения, весьма однобоки. Термин «экологическая ниша» был предложен Дж. Гриннелом в 1917 г. Вначале ниша определялась как понятие близкое к местообитанию. Сам Гриннел ввел термин «ниша» для обозначения самой мелкой единицы распространения вида. Определение ниши, данное Элтоном в 1927 г., охватывало главным образом функциональный аспект. Он описывал нишу как место данного организма в биотической среде в смысле пищевых связей и взаимоотношений с врагами, т. е. Элтон придавал нише трофический статус. Объединяя пространственный и трофический подходы к определению ниши, Хатчинсон в 1957 г. предложил рассматривать экологическую нишу как многомерное пространство. Он считал, что нишу следует определять с учетом всего диапазона физических, химических и биотических переменных среды, к которым должен быть адаптирован данный вид и под действием которых видовая популяция живет и возобновляется бесконечно долгое время. Градиент изменения каждого фактора можно рассматривать как некое измерение пространства. Если данная ниша имеет n-значимых измерений, то ее можно описать как n-мерное пространство, или гиперпространство. Это пространство реализуется живыми организмами через определенную условиями обитания численность (биомассу). Развивая свои представления, Хатчинсон (1957,1965) предложил различать фундаментальную нишу – «наиболее абстрактно заселенное гиперпространство», – и реализованную нишу – меньшее гиперпространство, занимаемое видом при биотических ограничениях. Первая отражает возможности вида в их полном объеме; вторая – более узкий спектр условий и ресурсов, допускающий поддержание жизнеспособной видовой популяции даже при наличии конкурентов, паразитов и хищников. Однако многомерное изображение ниши достаточно абстрактно, его трудно использовать для интерпретации конкретного экологического материала и дальнейших теоретических построений. Учитывая тот факт, что отделить значение для организма одних факторов среды от других порой практически невозможно, при описании ниши целесообразно выделить две группы факторов, два измерения: трофическое и пространственное. Иными словами экологическую нишу целесообразно подразделять на пространственную (описанную Гриннелом) и трофическую (описанную Элтоном). Трофическая ниша в теоретическом плане – количество и разнообразие пищевых объектов использованных или могущих быть использованными видом для питания, но в практическом плане – это просто численность (биомасса) данной популяции в данной пространственной нише. Отделить трофическую нишу от пространственной на практике бывает непросто. Для многих видов размеры их трофической ниши не поддаются определению вследствие ее громадных размеров. Об этом говорят многочисленные примеры расселения организмов в новых местах обитания (интродукция), а также обширнейшие ареалы всеядных видов-космополитов (вороны, крысы и др.). Напротив, в ряде случаев распространение видов строго определяется ареалом распространения их пищи. Так, сумчатый медведь – коала Phascolarctos cinereus в Австралии питается листьями определенных видов эвкалиптов, и его ареал ограничен в первую очередь ареалом распространения этих эвкалиптов. Во всех случаях о величине трофической ниши можно адекватно судить только по количеству организмов в данной популяции. Определение экологической ниши при таком подходе будет звучать как пространство (гиперпространство), способное поддерживать жизнеспособную популяцию вида определенной численности (биомассы). Отсюда, динамика сокращения фундаментальной ниши до реализованной при биотических ограничениях может описываться через изменение численности популяции, иными словами, через уже известную кривую толерантности Шелфорда.
Хищничество. В экологии под хищничеством понимают всякое поедание одних организмов другими. Существуют два основных способа классификации хищников. Согласно первой выделяют три основных типа хищников: истинные хищники, хищники с пастбищным типом питания и паразитоиды. Особенности этих типов хищников и их примеры приведены в таблице.
Истинные хищники | Хищники с пастбищным типом питания | Паразитоиды |
Убивают свою жертву более или менее сразу после того, как нападут на нее. | Используют в течение своей жизни большое число жертв, но съедают только ее часть. | Группа насекомых, выделенная на основе сходства в поведении взрослых самок при откладке яиц и типа последующего развития личинки. |
Дневные и ночные хищные птицы, отряды насекомоядные и хищные млекопитающие, зубатые киты, многие рыбы и др. | Травоядные копытные, грызуны и другие млекопитающие, растительноядные рыбы, питающиеся древесиной и листвой насекомые. | Представители отряда перепончатокрылых и двукрылых, например наездники. |
В связи с влиянием хищника на структуру популяции жертв, некоторые экологи условно подразделяют хищников на «расчетливых» и «нерасчетливых». Первые питаются, главным образом, «бесполезными» для популяции особями, вылавливая больных и старых, а также не нашедших себе территории особей низшего ранга, но не трогают особей способных к размножению, которые составляют источник пополнения популяции жертвы. Хищники «нерасчетливые» питаются эффективно особями всех групп и могут серьезно нарушить потенциал роста популяции жертвы.
Паразитизм – такая форма взаимоотношений двух различных организмов, носящая антагонистический характер, когда один из них (паразит) использует другого (хозяина) в качестве среды обитания и источника пищи, возлагая на него регуляцию своих отношений с внешней средой.
Различают периодический и постоянный паразитизм, а также факультативный и облигатный. Паразитизм известен на всех уровнях организации живого, начиная от вирусов, бактерий и грибов, кончая высшими растениями и животными. Среди паразитов различают также эндопаразитов, живущих в теле своего хозяина и питающихся его тканями, соками или содержимым пищеварительного тракта. К ним относятся сосальщики-трематоды, цепни, двуустки, аскариды, трипаносомы и др. Эктопаразиты живут, как правило, на поверхности хозяина и обладают достаточной подвижностью, чтобы переходить от одного хозяина к другому. Это блохи, вши, пухоеды, все ракообразные, паразитирующие на рыбах и т. д. В ряде случает сами паразиты в свою очередь становятся источником получения пищи и среды обитания для более мелких паразитов второго порядка. Таким образом, возникает особое явление называемое гиперпаразитизмом, или сверхпаразитизмом. Довольно много гиперпаразитов среди низших раков. Весьма уникальное явление – внутривидовой паразитизм. Он встречается лишь у специфических групп животных, например у глубоководных удильщиков. Гнездовой паразитизм – специфический способ заботы о потомстве преимущественно у птиц, при котором самка подкладывает оплодотворенные яйца в чужие гнезда; насиживает кладку и кормит птенцов другая птица. Гнездовой паразитизм теоретически можно отнести к периодическому паразитизму. Клептопаразитизм – насильственное присвоение одной особью корма, добытого другой, реже овладение кормом в отсутствии владельца, тайно. Клептопаразитизм широко распространен у птиц, млекопитающих и рыб, встречается также у насекомых.
«Конкуренция – это взаимодействие двух организмов, стремящихся к одному и тому же». Такое определение данному типу биотических взаимоотношений дал Ю. Одум (1975). Взаимодействие организмов путем «отнимания» ресурсов известно в экологической литературе, как косвенная конкуренция. Ее иногда называют еще эксплуатационной конкуренцией. Прямая конкуренция (интерференция) возникает при непосредственном взаимодействии организмов, будь то столкновения в результате опосредованного соперничества за ресурсы, или борьба за обладание особями противоположного пола и «место под солнцем». В крайних случаях прямая конкуренция принимает формы каннибализма. Если конкурирующие организмы принадлежат к одному виду, то взаимоотношения между ними называют внутривидовой конкуренцией; если же они относятся к разным видам, то их взаимоотношения называют межвидовой конкуренцией.
Неоднократно отмечалось в обширной литературе, что близкородственные организмы, ведущие сходный образ жизни, не обитают в одних и тех же местах. Если же они живут в одном месте, то потребляют разную пищу, активны в разное время или обладают еще какими-либо различиями, благодаря которым занимают несколько различающиеся ниши. Известное правило гласит, что не существует двух видов, которые могли бы занимать совершенно одинаковые ниши при совместном проживании. Это явление разобщения экологически близких видов получило название принципа конкурентного исключения, или закона Гаузе. Иногда закон Гаузе отождествляют с одним из его следствий, вытекающим из него. Примером может служить определение Л. Слободкина (1962): «Если два вида сохраняются в одном местообитании, то между ними должны существовать некоторые экологические различия» (цит. по Галлу, 1979). Синтопия – это экологическое разобщение близких видов при совместном проживании. Принцип конкурентного исключения не реализуется, когда время заметных изменений окружающей среды меньше того, какое требуется для вытеснения одного вида другим.
Комменсализм – буквально сотрапезничество – форма симбиоза, при которой один партнер извлекает пользу из сожительства, а другой к его присутствию безразличен. Наиболее простой тип положительных взаимодействий. В современном понимании комменсализма сотрапезничество, или нахлебничество – лишь одна из его форм. Кроме того, довольно известной другой формой комменсализма является квартиранство (синойкия). Комменсализм характерен, в частности, для взаимоотношений между прикрепленными растениями и животными, с одной стороны, и подвижными организмами – с другой. Особый случай комменсализма – зоохория. Это перенос животными семян, спор пыльцы растений. Перенос других более мелких животных называют форезией.
Мутуализмом называют взаимоотношения между парами видов, приносящие обоюдную пользу. В популяциях каждого из таких видов (мутуалистов) особи растут, размножаются и выживают с большим успехом в присутствии особей другого вида. Различают: факультативный, или «необязательный», мутуализм и облигатный, или постоянный (обязательный), мутуализм. Факультативный мутуализм называют также протокооперацией. Примером мутуализма великое множество. Однако, по причине сложности мутуалистических отношений часто бывает непросто отличить протокооперацию от облигатного мутуализма.
Проблемы для коллективного обсуждения
и вопросы для самоконтроля
1. Понятие о биотических факторах. Типы взаимодействия между популяциями. 2. Концепция экологической ниши. История вопроса. Понятие фундаментальной и реализованной ниши. 3. Конкуренция, ее виды. Принцип конкурентного исключения Гаузе. Синтопия, ее примеры. 4. Хищничество. Класификации хищников. Отношения хищник-жертва. 5. Эволюционные последствия хищничества. Активные и пассивные формы защиты от хищников, их примеры. 6. Паразитизм, его виды. 7. Комменсализм. Его основные формы, их примеры. Зоохория. 8. Понятие симбиоза. Облигатный и факультативный мутуализм. Протокооперация. Их многообразие.
Рекомендуемая литература
Биология. В 3-х томах. – М.: Мир, 1990. – Т. 2. – 325 с.
Кашкаров экологии животных. – Л.: Учпедгиз, 1945. – 338 с.
Наумов животных. – М.: Высшая школа, 1963. – 618 с.
, Харламова и среда: основы аутэкологии. – Мурманск: Пазори, 1998. – 274 с.
Основы экологии. – М.: Мир, 1975. – 740 с.
, Былова экология. – М.: Дрофа, 2004. – 416 с.
Шилов . – М.: Высшая школа, 1997. – 512 с.
ТЕМА ЛЕКЦИИ: Понятие популяции, ее динамика
и структура
Популяция – любая, способная к самовоспроизведению совокупность одного вида, более или менее изолированная в пространстве и времени от других аналогичных совокупностей того же вида.
При описании популяции, ее изучении используются обычно две группы показателей: статические, которые характеризуют состояние популяции в какой-то определенный момент времени, и динамические, характеризующие процессы, протекающие в популяции за некоторый промежуток времени. К статическим показателям относят общую численность популяции, плотность популяции, а также возрастную, половую, размерную и другие структуры популяции. К динамическим показателям – рождаемость, смертность, рост популяции и т. д.
Рождаемость – число новых особей, появившихся за единицу времени в результате размножения. Выделяют максимальную, или абсолютную (физиологическую), и экологическую (реализуемую) рождаемость. Максимальная рождаемость по сравнению с экологической является величиной постоянной для популяции.
Максимальная рождаемость | Экологическая рождаемость |
Образование теоретически максимально возможного количества новых особей в идеальных условиях (при отсутствии лимитирующих факторов). | Увеличение популяции при фактических условиях среды. Эта величина изменяется в зависимости от размера, состава популяции, физических условий среды. |
Величина рождаемости зависит от многих причин: соотношения полов, возрастных групп, доли особей, способных к размножению, частоты последовательных генераций и др. По числу периодов размножения в течение жизни у животных различают моноциклические и полициклические виды.
Моноциклические виды | Полициклические виды |
Все или значительная часть особей размножаются один раз в жизни и после этого гибнут. | Размножаются несколько раз в жизни. |
Поденки, майские жуки, морские миноги, тихоокеанские лососи и др. | Большинство видов животных. |
Смертность – количество погибших в популяции особей за определенный отрезок времени. Различают минимальную смертность (в идеальных условиях, отсутствуют лимитирующие факторы) и экологическую, или реализуемую, смертность, которая характеризует гибель в данных условиях среды. Минимальная смертность представляет собой теоретическую константу, используемую обычно для сравнения. Она определяется физиологической (максимальной) продолжительностью жизни. В этом плане наибольший интерес представляет выживание. На графике представлены три основных типа «кривой выживания» (Рис. 1).
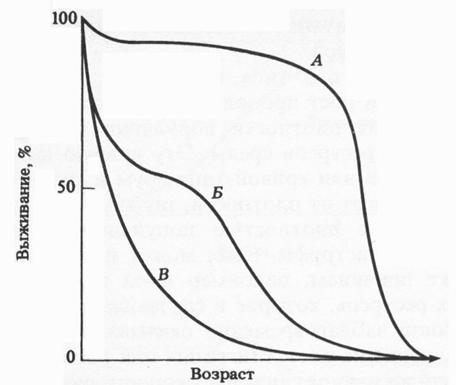
Рис. 1. Три типа кривых выживания (из Грина, Стаута, Тейлора, 1990).
Сильновыпуклая кривая I типа – «кривая дрозофиллы» – иллюстрирует низкую смертность до старости, после чего вся популяция вымирает за короткий промежуток времени; диагональная кривая II типа – постоянную смертность в популяции на протяжении всей ее жизни. Такое встречается среди рыб, птиц и пресмыкающихся. Сильно вогнутая кривая III типа демонстрирует массовую гибель особей в начальный период жизни, а затем низкую смертность оставшихся особей. Данный тип кривой выживания называют типом устрицы, так как этот моллюск ведет прикрепленный образ жизни во взрослом состоянии и имеет низкую смертность, а его планктонные личинки – очень высокую смертность. В кривой выживания обычно суммированы следующие взаимосвязанные показатели: число особей, остающихся в живых к концу данного периода, число смертей в каждой возрастной группе, смертность и ожидаемая продолжительность дальнейшей жизни. Таким образом, кривая выживания выражает зависимость между числом выживших особей (из гипотетической группы в 100 или 1 000 особей) и возрастом.
Плотность популяции – величина популяции по отношению к единице пространства. Плотность популяции определяется числом особей или биомассой популяции на единицу площади или объема. Выделяют среднюю плотность и специфическую, или экологическую.
Средняя плотность | Экологическая плотность |
Число особей или биомасса на единицу всего пространства. | Число особей (биомасса) на единицу заселенного пространства (доступной площади или объема, которые фактически могут быть заняты популяцией). |
Обычно для размеров популяции существуют верхний и нижний пределы. Верхний предел определяется продуктивностью экосистемы, трофическим уровнем данного организма, интенсивностью его метаболизма и т. д., или объемом ниши. Для нижнего предела нет точного определения. Он может определяться минимально необходимой частотой родительских особей для дачи полноценного потомства, исключая близкородственное скрещивание. Для поддержания оптимальной численности (или плотности) популяции в данных условиях действуют обычно гомеостатические механизмы (биотические взаимодействия).
Характер увеличения численности популяции может быть различным, соответственно различают несколько типов роста популяции, например экпоненциальный и логистический (Рис. 2). Первый рост описывается J-образной кривой. В этом случае плотность популяции быстро увеличивается по экспоненте, но затем, когда начинает действовать сопротивление среды или другой лимитирующий фактор, рост популяции внезапно прекращается. Логистический рост описывается S-образной кривой, и при этом типе роста увеличение популяции происходит вначале медленно (фаза положительного роста), затем быстрее (фаза приближения к логистическому росту), и вскоре под влиянием сопротивления среды рост популяции постепенно начинает замедляться (фаза отрицательного ускорения).
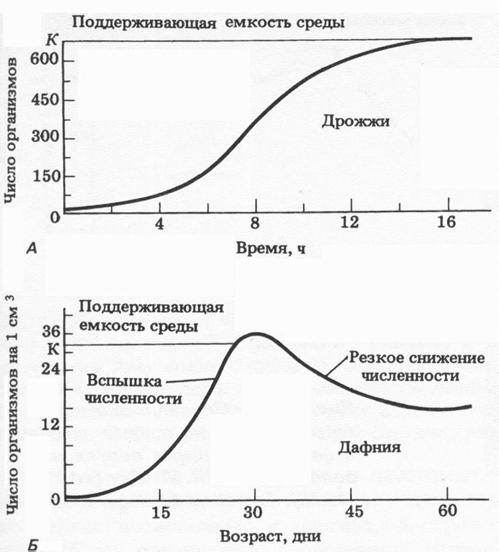
Рис. 2. Два типа кривых роста популяции: А – S-образная кривая роста; Б – J-образная кривая роста (из Грина, Стаута, Тейлора, 1990).
Константы «r» и «K» из логистического уравнения дали название двум типам естественного отбора. Согласно этой концепции среди множества разнообразных экологических стратегий, направленных на повышение вероятности выжить и оставить потомство, можно выделить два крайних типа: r-стратегия и K-стратегия. В случае r-стратегии отбор направлен на повышение скорости роста популяции в начальный период увеличения численности при слабом тормозящем действии конкуренции и малой плотности популяции. При K-стратегии отбор направлен на повышение выживаемости при стабильной численности и сильном действии конкуренции.
Характерные особенности r- и K-видов представлены ниже (Грин, Стаут, Тейлор, 1990).
r-виды | K-виды |
Размножаются быстро (высокая плодовитость, время генерации короткое), поэтому r (врожденная скорость роста популяции) высокое, скорость размножения не зависит от плотности популяции. Размножение идет с относительно большими затратами энергии и вещества. | Размножаются медленно (низкая плодовитость, продолжительное время генерации), поэтому значение r низкое, скорость размножения зависит от плотности популяции и быстро увеличивается, если плотность падает. Размножение идет с относительно малыми затратами энергии и вещества; большая часть энергии и вещества расходуется на нерепродуктивный (вегетативный) рост. |
Энергия и вещество распределяются между многими потомками. | Энергия и вещество концентрируются в немногих потомках; родители заботятся о потомстве. |
Размеры популяции некоторое время могут превышать K (поддерживающую емкость среды). | Размеры популяции близки к равновесному уровню, определяемому K. |
Вид не всегда устойчив на данной территории. Расселяются широко и в больших количествах; у животных может мигрировать каждое поколение. | Вид устойчив на данной территории. Расселяются медленно. |
Малые размеры особей и малая продолжительность их жизни. | Крупные размеры особей. Большая продолжительность их жизни. |
Местообитания сохраняются недолго. | Местообитания устойчивые и сохраняются долго. |
Слабые конкуренты. Не становятся доминантами. Защитные приспособления развиты сравнительно слабо. Лучше приспособлены к изменениям окружающей среды (менее специализированы). | Сильные конкуренты. Могут становиться доминантами. Хорошие защитные механизмы. Менее устойчивы к изменениям условий среды (высокая специализация для жизни в устойчивых местообитаниях). |
Примеры | |
Тли, мучные хрущаки, инфузории-парамеции и др. | Кондоры, альбатросы, крупные тропические бабочки и т. д. |
Пространственная структура популяций выражается характером размещения особей и их группировок по отношению к определенным элементам ландшафта и друг к другу и отражает свойственный виду тип использования территории. Закономерное распределение особей в пространстве имеет важное биологическое значение и, по существу, является основой всех форм нормального функционирования популяции. Пространственная структурированность популяций представляет собой «морфологическую» основу популяционного гомеостаза, определяя снижение уровня конкуренции и поддержания устойчивых внутрипопуляционных контактов как функциональных, так и информационных.
Различают следующие типы пространственного распределения особей в популяциях: равномерный, диффузный, агрегированный. Равномерный, или регулярный, тип распределения характеризуется равным удалением каждой особи от всех соседних; величина расстояния между особями соответствует порогу, за которым начинается взаимное угнетение. В природе равномерное распределение особей встречается достаточно редко. Близкий к этому характер распределения свойствен уплотненным популяциям некоторых сидячих беспозвоночных. Диффузный, или случайный, тип распределения особей встречается в природе значительно чаще, при нем особи распределены в пространстве неравномерно, случайно. Такой тип распределения широко представлен среди многих таксонов животных. Агрегированный (групповой, мозаичный) тип распределения выражается в образовании группировок особей, между которыми остаются достаточно большие незаселенные территории, и характерен для многих позвоночных животных, насекомых и др.
Численное соотношение различных категорий организмов в составе населения рассматривается как демографическая структура популяции. При этом в первую очередь имеется в виду соотношение половых (половая структура) и возрастных (возрастная структура) групп. Изменение этих показателей существенным образом влияют на темпы репродукции, и соответственно на общую численность популяции, ее изменение во времени. Возрастная структура популяции определяется соотношением различных возрастных групп (когорт) организмов в составе популяции. Особенно отчетливо возрастные различия проявляются у видов животных, развитие которых происходит с метаморфозом. В наиболее четкой форме половая структура популяции выражена у членистоногих и позвоночных животных. Половая структура динамична и в своей динамике тесно связана с возрастной структурой популяции. В связи с возрастом различают первичное, вторичное и третичное соотношение полов. Первичное соотношение полов определяется чисто генетическими механизмами, основывающимися на разнокачественности половых хромосом (X- и Y-хромосомы). В любом случае в процессе оплодотворения возможны различные комбинации половых хромосом, полученных от разных родителей, что и определяет пол каждой особи в потомстве. При таком механизме определения пола детерминируется статистически равномерное соотношение полов в потомстве. Это соотношение в момент оплодотворения и принимает за первичное. В результате различного рода воздействий на характер развития, а также неодинакового уровня смертности плодов разного пола соотношение самцов и самок среди новорожденных животных – вторичное соотношение полов – отличается от генетически детерминированного. Третичное соотношение полов характеризует этот показатель среди взрослых животных и складывается в результате дифференцированной смертности самцов и самок в ходе онтогенеза. Этот показатель прямо определяет особенности репродуктивного процесса и отличается у разных таксонов животных.
Проблемы для коллективного обсуждения
и вопросы для самоконтроля
1. Понятие популяции в экологии. Популяционная структура вида. 2. Динамика популяций. Понятие рождаемости, биотического потенциала, выживаемости, смертности, репродуктивной ценности, плотности и др. 3. Кривые выживаемости и роста популяций (логистическая и экспоненциальная модели). 4. Популяционные взрывы, периодические и непериодические колебания численности. Оптимальная эксплуатация популяции. 5. Экологические стратегии популяций, особенности r- и K-отбора.
Рекомендуемая литература
Харпер Дж., Экология. Особи, популяции и сообщества. В 2-х томах. – М.: Мир, 1989. – Т.1. – 667 с.
Гиляров экология. – М.: Изд-во МГУ, 1990. – 184 с.
Биология. В 3-х томах. – М.: Мир, 1990. – Т. 2. – 325 с.
Мантейфель поведения животных. – М.: Наука, 1980. – 220 с.
Наумов животных. – М.: Высшая школа, 1963. – 618 с.
Основы экологии. – М.: Мир, 1975. – 740 с.
, Былова экология. – М.: Дрофа, 2004. – 416 с.
Шилов . – М.: Высшая школа, 1997. – 512 с.
РАЗДЕЛ 4. Словарь терминов (Глоссарий). Приведен в УМК без разделения на темы, так как большинство терминов встречается во многих разделах.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
