
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Вид распределений по уровню ФКТ свидетельствовал о наличии в популяции семян трех фракций (субпопуляций, групп): I, II и III, как, например, показано на рис. 4 для партии семян с 50%-ной всхожестью. В каждой группе семена распределены нормально, семян с промежуточным уровнем ФКТ мало.
При проращивании из семян фракции I вырастали нормальные проростки. Из семян фракции II преимущественно выросли проростки с морфологическими дефектами, которые считаются ненормальными. Семена фракции III не прорастали, т. к., по-видимому, были мертвыми.
Таким образом, ранжируя воздушно-сухие семена по уровню ФКТ, можно выбрать семена, из которых вырастут проростки определенного качества. Отобрав ослабленные семена фракции II, можно было выяснить причину, по которой из этих семян вырастают проростки с морфологическими дефектами, и уменьшается всхожесть партии семян. Определив эту причину, можно было понять, каким образом стимулирующие воздействия ее устраняют.
III.1.2. Набухание семян бобовых
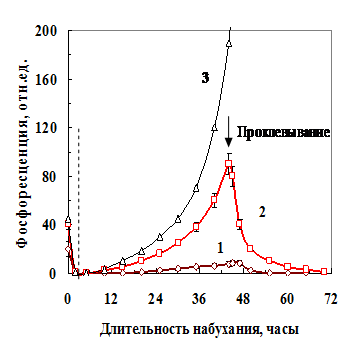
ФКТ воздушно-сухих семян уменьшалось до фона, когда в набухающем семени появлялась свободная вода. Однако, при дальнейшем увлажнении (при содержании воды в семени более 50%) у некоторых семян свечение возникало вновь, и могло в 5-10 раз превышать свечение семян в воздушно-сухом состоянии (рис. 6, кривая 3). Такие семена не прорастали (подвергались лизису).
|
О том, что есть для вида свечения свидетельствуют спектры. Спектр ФКТ сухих семян широкий. В спектре излучения набухших семян (рис. 2, кривая 2) присутствовали четыре характерных максимума. На этом основании и, учитывая спектр возбуждения свечения (рис.2, кривая 3), был сделан вывод, что свечение набухших семян является фосфоресценцией не содержащих металла порфиринов.
Рис. 6. Фосфоресценция проклюнувшихся (1 и 2) и не проклюнувшегося (3) семян гороха во время набухания. Стрелкой показан момент проклевывания. Слева от пунктирной линии показано затухание ФКТ сухих семян при их увлажнении
Когда зародышевый корешок у набухающего семени прорывал семенную оболочку (проклевывание семени), то свечение снижалось (кривые 1 и 2). Если уровень свечение был низкий, то вырастал нормальный проросток. При среднем уровне свечения семя проклевывалось, свечение падало, но из такого семени чаще всего вырастал проросток с морфологическими дефектами.
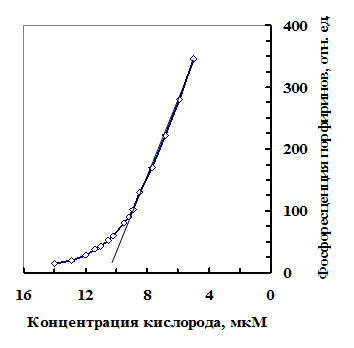
Известно, что наблюдать фосфоресценцию порфиринов можно лишь при очень низком содержании кислорода в среде (меньше 20 мкМ) [Теренин, 1967]. Это означает, что у набухающих семян светятся только те структуры, где содержание кислорода меньше 20 мкМ. Свечение набухших семян исчезало после нарушения целостности семенной оболочки. У “потухших” на воздухе семян свечение частично восстанавливалось в атмосфере азота или при помещении семян в раствор Na2SO3 (с целью удаления из воды кислорода), что доказывает дефицит кислорода под оболочкой семени [Веселова и др., 1985в; Veselova et al., 1988]. Как нами было показано, оболочка плохо проницаема для кислорода, а кислород активно поглощается зародышем набухающего семени в процессе дыхания [Веселова и др., 2003]. При содержании воды в семенах гороха 45-50% завершается митохондриогенез [Obroucheva, 1999]. Как показано на рис. 7, снижение концентрации кислорода в герметичной камере при митохондриальном дыхании зародышевых осей семян гороха приводит к пропорциональному возрастанию фосфоресценции порфиринов. Блокирование митохондриального дыхания цианидом замедляло поглощение кислорода и нарастание фосфоресценции порфи-
|
ринов [Веселова и др., 1985в].
Рис. 7. Соотношение между концентраций кислорода в камере и уровнем фосфоресценции порфири-нов зародышевых осей семян гороха.
Таким образом, уровень фосфоресценции эндогенных порфиринов у набухших семян может служить маркером степени недостатка кислорода под оболочкой.
III.1.3. ТЕРМОХЕМИЛЮМИНЕСЦЕНТНЫЙ МЕТОД НАБЛЮДЕНИЯ АМИНО-КАРБОНИЛЬНОЙ РЕАКЦИИ В ПОРОШКАХ СЕМЯН
Известно, что при хранении в семенах повреждения белков и нуклеиновых кислот вызывают продукты автоокисления мембранных липидов и процесс неферментативного гликозилирования восстанавливающими сахарами (амино-карбонильная реакция) [Smith, Berjak, 1995; Sun, Leopold, 1995]. Исследование роли этих процессов в снижении качества семян затрудняет определение их продуктов, имеющих сходные спектральные характеристики и присутствие в семенах флуоресцирующих соединений полифенольной природы [Feeney, Whitaker, 1982; Ory, St. Angelo, 1982; Baker, Bradford, 1994]. Известно, что хемилюминесценция возникает при автоокислении липидов [Тарусов, Журавлев, 1965] и гликозилировании белков [Castilho et al., 1994]. Обычно ее регистрируют при повышенной температуре. Так, например, термохемилюминесценцию листьев гороха и водорослей в красной области спектра (>650 нм), обусловленную терми-ческих распадом перекисей липидов, наблюдали ранее [Венедиктов и др. 1989; Stallaert et al., 1995].
Для определения продуктов перекисного окисления липидов и неферментативного гликозилирования белков в сухих семенах мы регистрировали температурные зависимости хемилюминесценции порошков семян (в сине-зеленой части спектра, максимум при 450 нм). В модельных опытах наблюдали ТХЛ в порошках аминокислот и белков с разными восстанавливающими сахарами, линоленовой кислотой (18:3) и глютаровым диальдегидом. Температуру объектов от комнатной до 190-200ОС повышали со скоростью 10О/мин.
| |
| Рис. 8. Термограмма хемилюминесценции порошка семян огурцов без добавок (1), и с добавлением 0,5% раствора глютарового диальдегида (2) и 2,5% порошка глюкозы (3). |
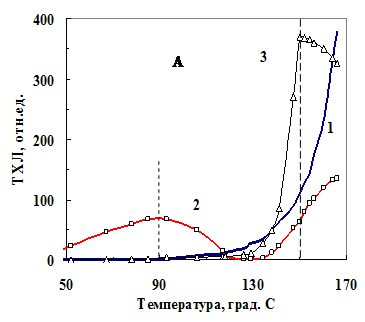
Термограмма порошка семян представляет экспонен-циально нарастающую кривую, начиная от 110ОС (рис. 8-А, кр. 1). ТХЛ смеси порошка семян с глюкозой свидетельствует о том, что при температуре выше 130ОС свечение обусловлено участием в хемилюминес-центной реакции глюкозы (максимум при 150ОС, кривая 3).
В области температур от 50 до 110ОС в ТХЛ, по-видимому, участвуют продук-ты перекисного окисления липидов, как показывает термограмма порошка семян при добавлении глютарового диальдегида (кривая 2).
Свечение порошка из мертвых семян при этих температурах в несколько раз превышало фон. После обработки порошка хлороформ-метанольной смесью свечение пропадало.
Иммобилизованная на кварцевом песке линоленовая кислота (18:3), имела низкий уровень ТХЛ (рис. 9, кривые 1, 2). В присутствии порошка триптофана
|
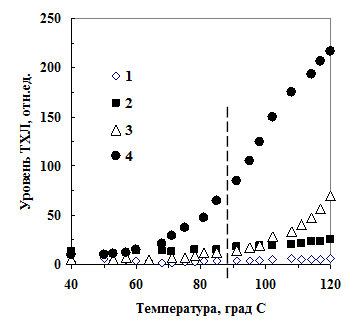 Рис. 9. ТХЛ не окисленной (1, 3) и окисленной (2, 4) линоленовой кислоты, иммобилизованной на кварцевом песке (1, 2) и порошке триптофана (3, 4).
Рис. 9. ТХЛ не окисленной (1, 3) и окисленной (2, 4) линоленовой кислоты, иммобилизованной на кварцевом песке (1, 2) и порошке триптофана (3, 4).ТХЛ окисленной кислоты много-кратно возрастала (кривая 4). Т. е. для возникновения ТХЛ в этой химической системе необходимо наличие аминогруппы. 0,5%-ный раствор глютарового диальдегида активировал хемилюминесценцию различных аминокислот и коллагена (данные не приведены). ТХЛ при 90ОС использовали как свидетельство присутствия в семенах продуктов перекисного окисления ненасыщенных жирных кислот.
|
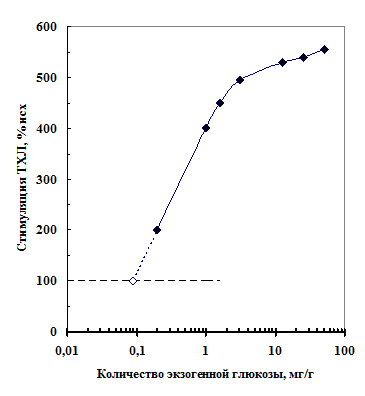 Рис. 10. Зависимость стимуляции уровня ТХЛ при 150ОС от количества глюкозы, добавленной к порошку семян гороха (черные ромбы).
Рис. 10. Зависимость стимуляции уровня ТХЛ при 150ОС от количества глюкозы, добавленной к порошку семян гороха (черные ромбы).В модельных опытах наблюдали ТХЛ в смеси глюкозы с различными аминокислотами и растительными белками. Эти вещества в отдельности не обладали ТХЛ. Термограммы смеси аминокислот и белков с глюкозой в координатах Аррениуса имели такой же наклон (энергию активации), что и термограмма порошка семян. Это позволило предположить, что свечение семян при высокой температуре обусловлено реакцией гликозили-рования.
Пропорционально увеличе-нию количества экзогенной глюкозы в порошке семян (от 0,2 до 5 мг/г) возрастал уровень ТХЛ при 150ОС (рис. 10). На основании этой зависимости можно показать, что количество эндогенной глюкозы в порошке семян гороха не превышает 0,1 мг/г семян. Подобное же значение (0,08-0,11 мг/г) содержания глюкозы в семенах гороха и сои приводятся в литературе [Locher, Bucheli, 1998].
На основании этих результатов уровень ТХЛ порошка сухих семян при 150ОС мы использовали как показатель содержания в них глюкозы.
III.2. ДИНАМИКА КАЧЕСТВА СЕМЯН ПРИ РАЗНЫХ ВОЗДЕЙСТВИЯХ
Поскольку нами был разработан метод оценки индивидуального качества семян, представляло интерес проверить, можно ли анализируя распределение воздушно-сухих семян по уровню ФКТ прогнозировать их качество в зависимости от дозы воздействия. В качестве действующих факторов были выбраны g-облучение и влаго-тепловая обработка семян (40ОС и 85% относительная влажность воздуха). Проникающая радиация, как и тепловая обработка, ускоряют старение семян [Roberts, 1972].
|

Рис. 11. Всхожесть семян гороха (1) и средний уровень ФКТ (2) через неделю после g-облучения. |
III.2.1. Изменение ФКТ воздушно-сухих семян гороха и их всхожести под влиянием g-облучения. С увеличением дозы g-облучения всхожесть семян гороха
и средний уровень ФКТ воздушно сухих семян до проращивания изменялись сложным образом. При малых дозах 190 мГр и 3 Гр всхожесть уменьшалась до 58 и 45%, соответственно (рис. 11, кривая 1), а уровень ФКТ возрастал (кривая 2). Всхожесть семян, облученных в дозах 7 и 10 Гр, была близка к исходной. Уровень ФКТ соответствовал свечению необлученных семян. После больших доз облучения всхожесть падала и при 100 Гр составляла 4%. ФКТ возрастало и превышало исходный уровень почти втрое у семян, облученных в дозе 100 Гр.
|
Рис. 12. Распределение по уровню ФКТ сухих семян гороха через неделю после облучения в разных дозах. Римскими цифрами I и II обозначены номера фракций. |
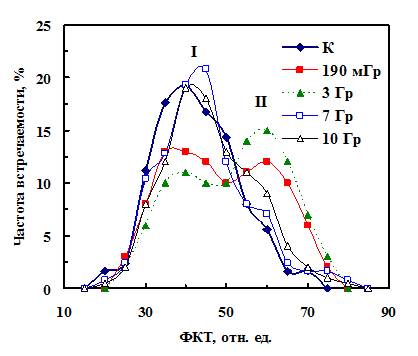
Изменение среднего уровня ФКТ определялось изменением фракционного состава партии (рис. 12). До облучения семян гороха их распределение по уровню ФКТ было унимодальным с максимумом при 40 отн. ед. (фракция I) и небольшим плечом при 60 отн. ед. (кривая К). Увеличение среднего уровня ФКТ после облучения в дозах 190 мГр и 3 Гр вызвано тем, что в распределении семян по уровню ФКТ количество семян фракции I уменьшилось и увеличилось число семян во фракции II (ФКТ 60 отн. ед., кривые 190 мГр и 3 Гр). Увеличение количества живых, но невсхожих семян (фракции II) означало, что в эту фракцию перешли семена из фракции I. После облучения в дозах 7 и 10 Гр распределения семян по уровню ФКТ мало отличались от контрольного. Часть семян из фракции II (невсхожих) перешла во фракцию I (всхожих). Мы назвали такие семена «улучшенными», поскольку они имели такую же всхожесть, как и необлученные семена фракции I. Поэтому и всхожесть семян, облученных в дозе 7 и 10 Гр стала близка исходной.
Таким образом, как и средний уровень ФКТ, анализ распределения сухих семян по уровню ФКТ позволяет прогнозировать изменения всхожести в зависимости от дозы g-облучения.
III.2.2. Изменение всхожести семян гороха после теплового воздействия и ФКТ воздушно-сухих семян. С увеличением продолжительности тепловой обработки (40ОС, 85% относительная влажность воздуха) всхожесть семян изменялась сложным образом (рис. 13, кривая 1). После индукционной фазы всхожесть снижалась, затем возрастала выше исходного уровня, и, наконец, необратимо падала. Эти изменения не связаны с изменением числа живых семян (число мертвых семян не меняется (кривая 2). Еще до проращивания изменения всхожести
Рис. 13. Всхожесть семян гороха (1), доля мертвых (2) и средний уровень ФКТ (3) сухих семян через неделю после тепловой обработки |
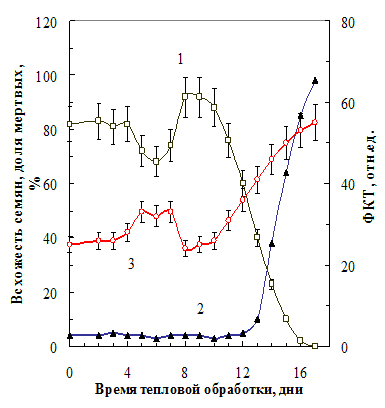
можно было предвидеть, регистрируя средний уровень ФКТ сухих семян (кривая 3). Изменения последнего в зави-симости от времени воздействия являлось зеркальным отражением кривой всхожести.
|
Рис. 14. Распределение воздушно-сухих семян гороха через неделю после 0 дней, 5 дней (1), 9, 14 и 16 дней тепловой обработки (40оС, 80%) по уровню ФКТ. I, II и III – номера фракций. |
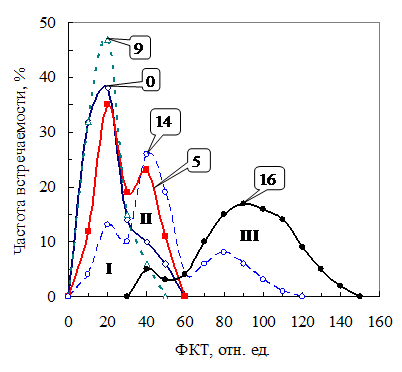
Изменение среднего уровня ФКТ было обуслов-лено варьированием фрак-циионного состава сухих семян. У исходных семян (всхожесть 82%) распреде-ление по уровню ФКТ близко к нормальному с небольшим плечом справа (рис. 14, кривая 0). Оно практически не менялось в течение первых четырех суток тепловой обработки. После 5-7 суток обработки появлялся новый макси-мум II (кривая 5). Это предполагает уменьшение всхожести. Распределение вновь становилось унимо-дальным после 8-9 суток теплового воздействия. Доля семян во фракции I увеличивалась (кривая 9) за счет семян перешедших из фракции II (невсхожих). Это привело к возра-станию всхожести.
После 12-14 дней тепловой обработки в распределении были представлены уже три максимума (кривая 14). Третий максимум свидетельствует о появлении мертвых семян. К 16-м суткам теплового воздействия в ФКТ-распределении оставался, в основном, максимум III, т. е. большинство семян погибло. Иными словами, изменение всхожести семян под влиянием теплового воздействия можно предсказать, анализируя фракционный состав партии воздушно-сухих семян ФКТ-методом.
При исследовании выхода электролитов в дистиллированную воду из отдельных семян, подвергнутых тепловой обработке, тоже было показано наличие фракций в партии семян гороха [Веселова и др., 1999а]. Из хороших семян фракции I выход электролитов был минимальным, При переходе семян во фракцию II выход электролитов возрастал вдвое. Переход семян из фракции II (невсхожих) во фракцию I («улучшенных») выход электролитов становился таким же, как и у необлученных семян. При переходе семян во фракцию III (отмирании) выход электролитов был в 5-5 раз выше, чем у необлученных семян, свидетельствуя о нарушении целостности клеточных мембран.
Переходы семян из одной фракции в другую, зарегистрированные по ФКТ, и выходу электролитов, имели скачкообразный характер. Например, после 4-х суток тепловой обработки большая часть семян находилась во фракции I, но уже через сутки значительная часть семян оказывалась во фракции II. Если после 7 суток экспонирования при повышенной температуре доля семян во фракции II была велика, то после 8-х суток большая часть семян вновь оказывалась вo фракции I.
Т. о. снижение и возрастание всхожести семян как при нарастании дозы g-облучения, так и в случае теплового воздействия, обусловлены изменением фракционного состава воздушно-сухих семян (числа семян во фракциях I и II).
III.2.3. Примеры изменения ФКТ распределений воздушно-сухих семян, подвергнутых действии разных факторов. На рис. 15 показаны распределения семян до (кривые 1) и после (кривые 2) стимулирующего воздействия. Стимулирующие всхожесть дозы были подобраны экспериментально (см. методический раздел). По характеру распределения можно еще до проращивания оценить результат воздействия.
Через 3 дня после светоимпульсной обработки семян овса вид распределения по уровню ФКТ изменялся (рис. 14, а). Доля семян во фракции II уменьшилась, и возросло число семян во фракции I. До обработки всхожесть партии составляла 69%, а после нее возросла до 83% за счет уменьшения числа ненормальных проростков.
|
| |
|
| |
Фосфоресценция при комнатной температуре, отн. ед. |
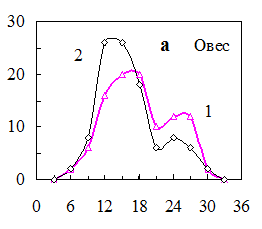
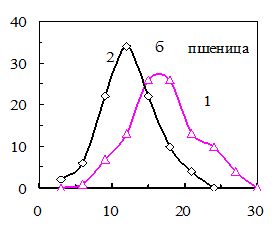
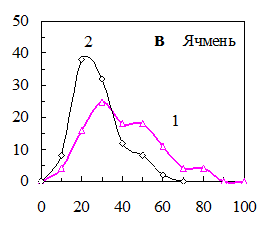
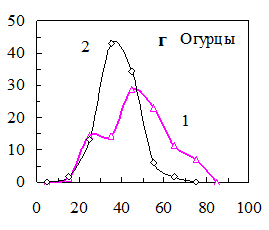
Рис. 15. Распределения семян по уровню ФКТ до (1) и после (2) а) светоимпульсной обработки семян овса (0,5 с, 50 МВт), б) обработки электрическим полем коронного разряда семян пшеницы (0,5 с, 2кВт/см2), в) озвучивания семян ячменя (200 гц, 5 мин, 65 дБ), импульсного облучения гелий-неоновым лазером семян огурцов (632,5 нм, 0,3 мВт/см2, 100 имп. по 50 мкДж/см2).
Через три дня после воздействия на семена пшеницы электрического поля коронного разряда происходило аналогичное изменение распределения семян по уровню ФКТ. Уменьшалась доля семян фракции II и увеличивалась доля семян во фракции I (рис. 14, б). Всхожесть возросла от 47 до 88%.
Озвучивание семян ячменя с частотой 200 Гц увеличивало всхожесть семян от 67 до 85%. При этом доля семян с низким уровнем ФКТ возрастала, и уменьшалось число семян с более высоким уровнем ФКТ (рис. 14, в).
После импульсного облучения семян огурцов гелий-неоновым лазером необходима была двухдневная “отлежка”, после которой можно было наблюдать переход некоторой части семян с высоким уровнем ФКТ во фракцию с более низким свечением (рис. 14, г). Всхожесть семян огурцов возрастала от 82 до 91%.
Иными словами, предпосевная обработка сухих семян различными по природе факторами в стимулирующих их всхожесть дозах всегда сопровождается неспецифическим увеличением доли семян во фракции I.
III.2.4. Стимулирующий эффект зависит от качества семян и уровня воздействия. В опытах использовали семена гороха различной всхожести (98, 82, 80, 72, 56, 54, 50 и 48%). С увеличением длительности теплового воздействия (40ОС, 85% относительная влажность воздуха) всхожесть сначала снижалась, а затем возрастала (стимуляция). Причем эти изменения были сильнее выражены у семян с низкой всхожестью. У семян с 56%-ной всхожестью она сначала снижалась до 18-20%, и затем возрастала до 64%. У семян высокого качества (всхожесть 98%) термическое воздействие таких изменений всхожести не вызывало.
Анализ фракционного состава партии семян по ФКТ показал, что стимуляция была выше в партии, в которой было больше семян фракции II.
У семян гороха 16%-ной влажности при 40ОС эффект хорошо воспроизводился. Если такой же тепловой обработке подвергали семена с влажностью 20%, что увеличивает их термочувствительность, то стимулирующий эффект отсутствовал, а наблюдали только однонаправленное снижение всхожести. Когда вместо увеличения влажности семян, использовали прогрев при 45ОС (увеличивали силу воздействия), то стимулирующий эффект тоже пропадал.
Эти наблюдения подтверждают выводы других авторов о роли состояния объекта и мощности действующего фактора для проявления стимулирующих эффектов при воздействиях, имеющих разную природу [Преображенская, 1971; Александров, 1985; Кузин 1995].
На основании вышеприведенных данных мы пришли к выводу, что разнообразные факторы вызывают у воздушно-сухих семян однотипные изменения фракционного состава партии. С увеличением дозы воздействия возрастает доля семян фракции II и всхожесть снижается. В области доз, стимулирующих всхожесть, число семян фракции II уменьшается, и они переходят во фракцию I. Дальнейшее возрастание дозы воздействия приводит к появлению семян фракции III – мертвых. Эта закономерность подсказывает, что для исследования механизмов стимуляции всхожести семян, необходимо, знать как минимум фракционный состав партии, или лучше использовать отдельные фракции.
III.2.5. Свойства разных фракций на примере семян гороха. Для исследования из партии необлученных отобрали семена фракции I (уровень ФКТ 20-30 отн. ед.). Из партии, облученных в дозе 3 Гр отобрали семена фракции II (50-60 отн. ед.) и мертвые семена фракции III (80-100 отн. ед.). Кроме того были использовали «улучшенные» семена (20-30 отн. ед.) из партий семян, облученных в дозе 10 Гр. Эти семена, судя по уровню ФКТ, «возвратились» из фракции II во фракцию I. Из таких семян тоже вырастали нормальные проростки.
Прежде всего, мы хотели найти отличия у семян, из которых вырастают нормальные и ненормальные проростки, а также выяснить, на каком этапе прорастания семян фракции II у проростков возникают морфологические дефекты.
Семена разных фракций при одной и той же относительной влажности воздуха содержат разное количество воды. Как следует из измерения ФКТ, сухие семена фракций II и III менее гидратированы. Определение влажности стандартным весовым методом показало, что семена гороха из фракции I (исходных и «улучшенных») содержали 9,81 и 9,84 % воды, соответственно, а из фракции II - 8,9 %, а фракции III – 8,2 %. Это может быть обусловлено как разной водоудерживающей способностью биополимеров семян, так и разной проницаемостью мембран клеток для воды.
|
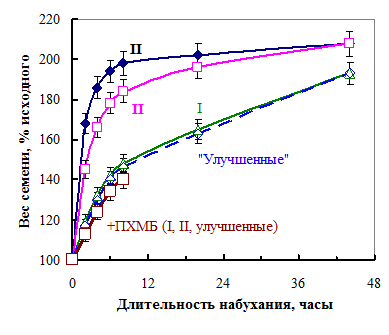 Рис. 16. Кинетика набухания семян разных фракций (I-III) на влажной фильтровальной бумаге и семян фракций I, II и «улучшенных» в присутствии ПХМБ.
Рис. 16. Кинетика набухания семян разных фракций (I-III) на влажной фильтровальной бумаге и семян фракций I, II и «улучшенных» в присутствии ПХМБ.Сухие семена из разных фракций не только удерживали разное количество воды, но и с разной скоростью поглощали ее при набухании (рис. 16). Семена второй фракции вдвое быстрее поглощали воду при набухании, чем семена фракции I и «улучшенные». Семена третьей фракции поглощают воду активнее, чем семена фракции II. Семенная оболочка является главным барьером на пути воды в семя. Как мы показали ранее, семенная оболочка одинаково тормозила поступление воды в семена фракций I и II, т. е. не оболочка была причиной более быстрого поступления воды в семена фракции II по сравнению с семенами фракции I.
Семена гороха разных фракций отличались не только по влажности и скорости поступления воды в них при набухании, но и по выходу электролитов в это время. Скорость выхода электролитов из семян фракции II вдвое, а из фракции III в 3-5 раз выше, чем из семян фракции I и «улучшенных» семян [Веселова и др., 1999].
|
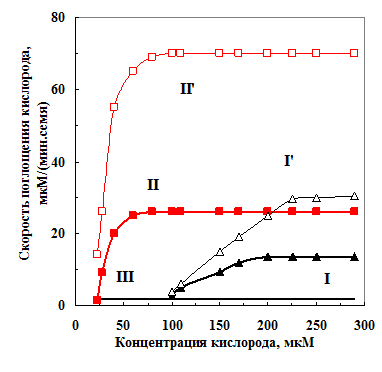 Рис. 17. Зависимости скорости поглощения
Рис. 17. Зависимости скорости поглощениякислорода набухавшими 20 ч семенами гороха I, II и III фракций в оболочке и без нее (I’, II’) от концентрации кислорода в камере |
III.2.6. Взаимосвязь между скоростью набухания и возникновением дефицита кислорода у семян гороха. Появление в семенах при набухании свободной воды активирует поглощение кислорода. При влажности 30-35% скорость поглощения кислорода выходит на стационарный уровень. Когда при влажности семян гороха 45% заканчивается митохонд-риогенез [Obroucheva, 1999], скорость поглощения кисло-рода вновь резко возрастает. Сродство дыхания к кислороду увеличивается. На рис. 17 показаны зависимости скорости поглощения кисло-рода семенами от его концен-трации в среде. После 20 ч набухания семена I имели влажность 43%, а II фракции - 50%, т. е. у семян фракции II уже включилось цитохромное дыхание (рис. 17).
Регистрация поглощения кислорода семенами в оболочке и без нее показала, что оболочка семян фракции I тормозила поглощение кислорода в 2,1 раза (в оболочке 15 мкМ, а без оболочки – 32 мкМ О2/(мин семя), а семенами фракции II в 2,6 раза (29 и е 76 мкМ О2/(мин. семя), соответственно. Очевидно, более высокая скорость поглощения зародышем кислорода в процессе дыхания и медленная диффузия последнего через оболочку была причиной дефицита кислорода у зародыша семян фракции II.
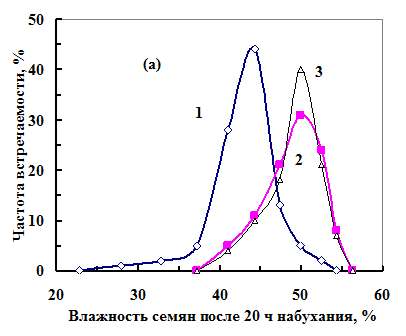
Из-за разной скорости набухания семян после 20 ч набухания семена сформировались две на группы (рис. 18, а) в соответствии с фракциями I и II, выбранными по ФКТ. Из семян фракции I (максимум в распределении по влажности на 44% (кривая 1)) вырастали нормальные проростки. Максимум в распределении семян фракции II приходился на влажность 50%. Некоторые семена этой фракции не прорастали (кривая 3), а из других вырастали ненормальные проростки (кривая 2).
Как показано на рисунке 18,б, семена фракции I или не светились, или имели низкий уровень фосфоресценции порфиринов (кривая 1). Семена фракции II с уровнем фосфоресценции порфиринов 20-40 отн. ед. (соответствует концентрации кислорода под оболочкой 10-12 мкМ, рис. 6), обычно проклевывались, но из них вырастали ненормальные проростки (кривая 2).
|
|
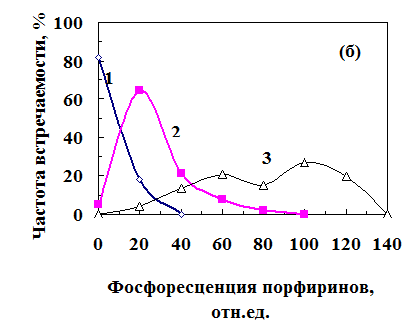
Рис. 18. Распределение семян по влажности (а) и фосфоресценции порфи-ринов (б) в партии семян с 72%-ной всхожестью после 20 ч набухания. 1 – семена фракции I, из которых выросли нормальные проростки; 2 - семена фракции II, из которых выросли ненормальные пророст-ки; 3 – ненаклюнувшиеся семена.
Семена с высоким уровнем свечения - 40 и более отн. ед. (концен-трация кислорода менее 10 мкМ) не проклевы-вались. Поскольку зародышевой корешок не мог проткнуть семенную оболочку, то они задохнулись во время набухания (кривая 3).
У мертвых семян фосфоресценция порфиринов не возникала, т. к. они не дышали (рис. 17, прямая III). Поэтому можно было отличить задохнувшиеся семена от исходно мертвых.
Если у семян с высоким уровнем фосфоресценции порфиринов (значительный дефицит кислорода) удаляли оболочку, то некоторые из них прорастали, но проростки имели морфологические нарушения, т. е. были ненормальными.
Оставалось неясным, почему даже после восстановления нормальной обеспеченности зародышей кислородом из семян фракции II вырастали проростки с морфологическими дефектами.
III.2.7. Синтез общих белков и ДНК на ранних стадиях набухания семян гороха. Для прорастания семян и нормального роста необходимы синтез белков и ДНК. Нарушение синтеза ДНК препятствует делению клеток зародышевых осей. Торможение синтеза белка нарушает процесс растяжения клеток [Гумилевская и др., 1996; Obroucheva, 1999]. Вероятно, нарушение этих процессов могло быть причиной появления проростков с морфологическими дефектами из семян фракции II. Как мы показали ранее, судя по включению меченого [35S]метионина в белки осевых органов в течение первых 3-х часов набухания семян гороха, незначительные различия в скорости этого процесса у семян первой и второй фракций определяются только разной скоростью набухания [Veselova et al., 2004].
Также был изучен процесс репликации ДНК на стадии подготовки клеток зародышевых осей к делению. После созревания семян большинство ядер в меристематических клетках содержат 2С набор ДНК (фазы G1 или G0 клеточного цикла) [Обручева, 1982; Bino et al., 1993; Redfearn et al., 1995; Gornik et al., 1997]. В сухих семенах гороха I II и III фракций около 90% ядер содержали 2С набор ДНК (табл. 1). С увеличением времени набухания увеличивалась число ядер в фазе G2, содержащих 4С набор ДНК. Динамику этого процесса отражает отношение 4С/2С.
У мертвых семян фракции III удвоение ДНК не происходило и количество ядер с 2С набором ДНК не изменялось. В случае семян гороха фракций I и II в течение 48 часов набухания количество ядер с 2С набором ДНК уменьшалось и увеличивалось число ядер с 4С набором ДНК. Причем у семян фракции II, которые набухали быстрее и имели большую влажность, отношение росло сильнее. Рост отношения 4С/2С означал, что повреждения ДНК либо были незначительны, либо репарированы на ранних стадиях гидратации. Известно, что процесс удвоения ДНК может начинаться только после ликвидации ее нарушений [Osbornе, 1983; Smith, Berjak, 1995].
Таблица 1. Соотношение ядер в состояниях 2С и 4С у зародышевых осей семян гороха разных фракций во время набухания (%).
Время набухания, ч. | Фракция I | Фракция II | Фракция III | ||||||
2C | 4C | 4C/2C | 2C | 4C | 4C/2C | 2C | 4C | 4C/2C | |
0 | 88,0 | 12,0 | 0,14 | 88,0 | 12,0 | 0,14 | 91 | 9,0 | 0,10 |
6 | 84,7 | 15,3 | 0,18 | 82,6 | 17,4 | 0,21 | 90 | 9,0 | 0,10 |
24 | 75,4 | 23,6 | 0,30 | 71,4 | 28,8 | 0,40 | 90 | 10,0 | 0,11 |
48 | 56,4 | 39,5 | 0,70 | 54,0 | 46,0 | 0,85 | 90 | 10,0 | 0,11 |
72 | 30,7 | 63,0 | 2.02 | 53,5 | 46,5 | 0,87 | 89 | 11,0 | 0,12 |
Однако после проклевывания на 48 ч набухания увеличение отношения 4С/2С у семян фракции II прекращалось, в то время как у семян фракции I оно продолжало расти. Задержка репликации ДНК происходила накануне или во время проклевывания. Она могла быть вызвана продуктами брожения, образующимися под семенной оболочкой [Al-Ani et al., 1985]. Дефицит кислорода под семенной оболочкой у семян фракции II возникал после 12-14 ч набухания и продолжался до момента проклевывания (40-46 ч набухания). Тем не менее, в это время удвоение ДНК происходило, т. е. не гипоксия в данном случае являлась причиной прекращения репликации ДНК.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
