
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
ВЕСЕЛОВА Татьяна Владимировна
ИЗМЕНЕНИЕ СОСТОЯНИЯ СЕМЯН ПРИ ИХ ХРАНЕНИИ, ПРОРАЩИВАНИИ И ПОД ДЕЙСТВИЕМ ВНЕШНИХ ФАКТОРОВ (ИОНИЗИРУЮЩЕГО ИЗЛУЧЕНИЯ В МАЛЫХ ДОЗАХ И ДРУГИХ СЛАБЫХ ВОЗДЕЙСТВИЙ), ОПРЕДЕЛЯЕМОЕ МЕТОДОМ ЗАМЕДЛЕННОЙ ЛЮМИНЕСЦЕНЦИИ
03.00.01-03 - радиобиология
03.00.02-03 - биофизика
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
доктора биологических наук
Москва, 2008 г.
Работа выполнена на кафедре биофизики Биологического факультета Московского Государственного университета им.
Официальные оппоненты:
доктор биологических наук, профессор
доктор биологических наук, профессор
доктор биологических наук, акад. РАЕН
Ведущая организация – ВНИИ сельскохозяйственной радиологии и агроэкологии, Обнинск
Защита состоится «__» _______ 200_ г. в 15.30 часов на заседании диссертационного совета Д.501.00.65 при Московском Государственном Университете им. Москва, Ленинские горы, дом 1, МГУ, корп. 12, Биологический факультет, ауд. ___.
С диссертацией можно ознакомиться в библиотеке Биологического факультета МГУ им. . Отзывы просим присылать Москва, Ленинские горы, МГУ им. , биологический факультет. Факс (4
Автореферат разослан «__»__________2008 г.
Ученый секретарь диссертационного совета
доктор биологических наук, профессор
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования. Существует проблема разнонаправленного действия больших и малых доз ионизирующей радиации. Если причины повреждающего действия больших доз достаточно хорошо изучены, то вопрос о стимулирующем действии малых доз до сих пор остается открытым, несмотря на многочисленные исследования в этой области [Бреславец, 1946; Кузин, 1995; Преображенская, 1971; , Гудков, 1973; Ижик, 1976; Савин, 1981; , 1989; Райнхарт, 1998; Miller, Miller, 1987; Sheppard, Regitnig, 1987; Mokobia et al., 2006]. Интерес к проблеме вызван перспективой использования явления радиационной стимуляции растений в сельском хозяйстве с целью увеличения продуктивности растений и получения более высокого урожая.
Большинство работ по радиостимуляции растений выполнено путем облучения элитных семян, результат наблюдают по урожаю. Однако урожай в первую очередь зависит от всхожести семян. Ее уменьшение даже на 10-20% приводит к двух–трех-кратному снижению урожая [Реймерс, 1987].
При хранении семена старятся, качество и всхожесть семян снижаются, поэтому в партии семян, хранившейся несколько лет, присутствуют сильные семена, слабые (живые, но не прорастающие) и мертвые.
Известны приемы предпосевной обработки семян, с помощью которых можно увеличить всхожесть семян, утраченную при хранении. Ионизирующая радиация в малых дозах, озвучивание [Данько и др. 2000], кратковременная тепловая [Priestley, 1986; Реймерс, 1987] и ударно-волновая обработки [Игнатьев, 1976], экспонирование в электрическом и магнитных полях [Аносова и др., 1992; Бондаренко и др., 1998], лазерное облучение [Инюшин, 1978; Числова, 1988], предпосевное замачивание в растворах биологически активных веществ и др. могут увеличить всхожесть семян и урожай на 15-25% [Бреславец, 1946; Преображенская, 1971; Ижик, 1976; Кузин, 1995; Савин, 1981; Гудков, 1985; ,1989].
Возникает естественный вопрос, каким образом воздушно-сухие семена, которые годами старели, накапливали повреждения и теряли всхожесть, в результате кратковременной предпосевной обработки приобретают способность прорастать. Для того чтобы ответить на этот вопрос, надо, прежде всего, иметь представление, какие изменения в стареющих семенах приводят к снижению всхожести – появлению не прорастающих, но живых семян. Всхожесть может быть увеличена только за счет этих семян.
Старение и гибель семян обусловлены нарушением целостности клеточных мембран и повреждением ДНК [обзоры, Priestley, 1986; Bewley, Black, 1994; Smith, Berjak, 1995]. Предполагают, что это вызвано продуктами свободнорадикального перекисного окисления мембранных липидов. Процесс старения семян уподабливают окислительному стрессу [Stewart, Bewley, 1980; Barber, 1984; Thompson et al., 1987; Senaratna, et al,. 1988; Hendry, 19975; Simontacchi, Puntarulo, 1994; Smith, Berjak, 1995]. В последние годы обратили внимание на то, что старение семян, как и животных и человека, сопряжено с процессом неферментативного гликозилирования белков и нуклеиновых кислот (реакция Амадори-Майларда) [Серами и др., 1987; Растинг, 1993; Sun, Leopold, 1995].
Существует гипотезы, объясняющие стимуляцию жизнедеятельности растительного организма, слабыми воздействиями. Одна из них предполагает, что уровень метаболизма возрастает из-за ослабления контроля со стороны регуляторных механизмов, которые в норме ограничивают функциональную активность клетки [Александров, 1985], то есть имеет место проявления правила Арндта-Шульца. Согласно другой распространенной гипотезе стимуляция роста и развития растения трактуется как следствие гиперфункции репарационных процессов в ответ на первоначальное повреждение и появление малых количеств клеточных токсинов [Бреславец, 1946; Тимофеев-Ресовский, 1956; , Гудков, 1973; Савин, 1981; , 1989; Кузин, 1995].
У организмов в состоянии метаболического покоя (воздушно-сухие семена, пыльца, споры и др.) считают, что облучение в малых дозах и другие физические воздействия оставляют в клетках скрытые (потенциальные) повреждения, которые реализуются во время перехода клеток в жизнедеятельное состояние [Кузин, 1995]. Естественно, что предполагаемые механизмы стимуляции могут включаться у семян только во время их прорастания. Но еще до набухания в облученном семени развиваются пострадиационные физико-химические процессы. Так, после больших доз облучения состояние семян в процессе хранения ухудшается, они теряют всхожесть («эффект хранения»). Эффект стимуляции растений из семян, облученных в малых дозах, при затягивании сроков высева пропадает.
Для стимуляции всхожести воздействию обычно подвергают неоднородные партии хранящихся семян, состоящие из сильных, слабых и мертвых. Поскольку увеличить всхожести партии семян можно лишь за счет живых семян, не прораста-ющих при данных условиях, то для исследования механизма стимулирующего действия g-радиации и других факторов необходимо иметь воздушно-сухие семена однородные по качеству. Но задача деления партии таких семян на фракции разного качества до проращивания до сих пор практически не была решена.
Цель и задачи исследования
Целью работы было выяснить механизм изменения всхожести семян при их хранении и под действие внешних факторов (ионизирующего излучения в малых дозах и других слабых воздействий).
В задачи работы входило:
1) разработка люминесцентного метода анализа качественного состава партий воздушно-сухих семян и прогнозирования их всхожести без проращивания;
2) установление взаимосвязи между люминесцентными характеристиками индивидуальных сухих семян и качеством вырастающих из них проростков;
3) исследование влияния ионизирующей радиации и других факторов, в обычно используемых для стимуляции всхожести, на качественный состав партии сухих семян;
4) выяснение причины, препятствующей прорастанию ослабленных семян и роли окислительного стресса в образовании проростков с морфологическими дефектами;
5) исследование динамики всхожести семян в период после действия на семена ионизирующего излучения и тепловой обработки.
На защиту выносятся следующие положения:
1. Изменение всхожести семян под влиянием ионизирующего излучения, тепловой обработки, и других физических факторов (озвучивание, механические воздействия, лазерное облучение, электрические и магнитные поля) является неспецифическим ответом, который определяется, в основном, процессами, происходящими в сухих семенах до проращивания.
2. Кратковременная стимуляция всхожести семян под влиянием физических факторов разной природы обусловлена ускорением процесса естественного старения (накопления повреждений).
Научная новизна работы. Впервые показано, что при помощи регистрации фосфоресценции при комнатной температуре (ФКТ) у воздушно-сухих семян до проращивания можно прогнозировать изменение всхожести после предпосевной обработки факторами различной природы в малых дозах (тепловое, ионизирующее излучение, акустическое воздействие, электрическое поле коронного разряда, комбинированное магнитное поле, лазерное излучение).
Измерение уровня ФКТ индивидуальных сухих семян (злаковых, бобовых, огурцов, сосны) и анализ распределения семян по уровню ФКТ впервые позволило разделять партии воздушно-сухих семян пониженной всхожести на три дискретных фракции, отличающихся по качеству.
Повышение всхожести партии можно прогнозировать по распределению сухих семян по ФКТ, если воздействие вызывает увеличение доли семян во фракции I (всхожих) за счет их перехода из фракции II (живых, но невсхожих).
Причиной стимуляции всхожести семян бобовых низкого качества после g-облучения и других воздействий в малых дозах является модификация клеточных мембран, которая сопровождается замедлением поступления воды в клетки при набухании.
Впервые показано, что, регистрируя фосфоресценцию эндогенных порфиринов у набухающих семян можно оценивать уровень дефицита кислорода под семенной оболочкой.
Нарушение процесса деления у ненормальных проростков было следствием пост-гипоксического окислительного стресса после проклевывания семян. При доступе воздуха к зародышу после гипоксии наблюдали образование активных форм кислорода, в основном, Н2О2, при аэрации семян после гипоксии наблюдали хемилюминесцентным методом в присутствии люминола. Отсутствие гипоксии у семян, из которых вырастают нормальные проростки, обусловлено их более медленным набуханием, при котором потребление кислорода при дыхании зародыша компенсируется его диффузией через оболочку семена.
Анализ уровня ядерной ДНК при подготовке клеток зародышевой оси к делению (переход 2С![]() 4С) показал, что у семян фракции II торможение репликации ДНК совпадает во времени с возрастанием количества Н2О2.
4С) показал, что у семян фракции II торможение репликации ДНК совпадает во времени с возрастанием количества Н2О2.
Практическое значение работы.
Разработан экспрессный метод, основанный на явлении фосфоресценции при комнатной температуре (ФКТ), для определения такого важного показателя качества семян как влажность (А. с. № 1 1981). Метод позволяет оценить разницу во влажности образцов в 0,1-0,2% у семян и биопрепаратов (муки, крупы, конидий и др.) при содержании в них воды от 4 до 20% от сырого веса.
Регистрация ФКТ индивидуальных воздушно-сухих семян позволяет характеризовать гетерогенность партии семян по влажности. Метод может быть рекомендован для выделения из партии ослабленных и мертвых семян.
С помощью метода ФКТ можно наблюдать последействие факторов разной природы на воздушно-сухие семена до их проращивания и контролировать жизнеспособность семян (А. с. № 1 1982).
Разработан метод выделения из партии элитных семян огурцов семян, имеющих высокую потенциальную продуктивность (максимальный урожай) (А. с. № 1 1988).
Разработан метод контроля гипоксии у прорастающих семян, основанный на регистрации фосфоресценции эндогенных порфиринов. Показано, что набухание семян в присутствии антиоксидантов (например, пропилгаллата) уменьшает повреждение зародыша при пост-гипоксическом окислительном стрессе и увеличивает всхожесть семян.
Апробация работы
Материалы диссертации доложены на: Межфакультетской научно-практической конференции “МГУ – сельскому хозяйству”, 1982 (Москва); отчете по программе сотрудничества стран-членов СЭВ и СФРЮ по проблеме исследования в области биологической физики, 1984 (Пущино), Первой республиканской конференции по биофизике Молдавии, 1984 (Кишинев); Всесоюзном научном совещании “Люминесцентные методы исследования в сельском хозяйстве и перерабатывающей промышленности”, 1985 (Минск); Всесоюзном симпозиуме “Биохемилюминесценция в медицине и сельском хозяйстве”, 1986 (Ташкент); Всесоюзной конференции по биотехнологии злаковых культур, 1988 (Алма-Ата); Всесоюзном симпозиуме “Физиология семян”, 1988 (Душанбе); Всесоюзной конференции “Измерительная и вычислительная техника в управлении производственными процессами в АПК”, 1988 (Ленинград); Симпозиуме Интернациональной ассоциации по тестированию семян “Технологический прогресс и исследования семян”, 1994 (Вагенинген, Голландия); III съезде общества физиологов России “Физико-химические проблемы физиологии растений”, 1996 (Пенза); II Международном научно-практическом симпозиуме по селекции и семеноводству, 1997 (Аранжелович, Югославия); Международной школе “Проблемы теоретической биофизики”, 1999 (Москва); II Съезде Биофизиков России, Москва, мгу, 1999; IV съезде общества физиологов растений России “Физиология растений – наука III тысячелетия”, 1999 (Москва); Международной научно-практической конференции «Семя», 1999 (Москва); Школе-конференции “Горизонты физико-химической биологии”, 2000 (Пущино-на-Оке); Международной конференции “Растения под стрессом окружающей среды”, 2001 (Москва); V конференции “Кислород, свободные радикалы и окислительный стресс у растений”, 2001 (Ницца, Франция); VII международном рабочем совещании по семенам, 2002 (Саламанка, Испания); Международной конференции “Семена древесных”, 2002 (Ханья, Греция); Международном рабочем совещании “Новые достижения в улучшении качества семян”, 2003 (Лодзь, Польша).
Публикации. По материалам диссертации опубликовано в 46 печатных работах, в том числе: 2 монографии, 17 статей в журналах из списка ВАК, 7 работ в рецензируемых журналах, 4 авторских свидетельства, 7 статей в сборниках, и 10 тезисов.
Структура и объем диссертации
Диссертация изложена на ___ стр. машинописного текста. Состоит из введения, обзора литературы (глава I), описания объектов и методов исследования (как стандартных, так и разработка новых) (глава II), изложения полученных результатов (глава III), их обсуждения (глава - IV) выводов, списка литературы и приложения. Текст иллюстрирован 18 таблицами и 75 рисунками. Список литературы включает 159 отечественных и 271 зарубежных работ.
СОДЕРЖАНИЕ РАБОТЫ
Во введении дано обоснование актуальности выбранной темы в связи с имеющимися на сегодняшний день исследованиями по стимуляции жизнедеятельности растений после действия ионизирующего излучения в малых дозах и действия других факторов. Сформулированы цели и задачи исследования.
В обзоре литературы (глава I), состоящем из четырех разделов, представлен обзор данных о радиостимуляции, предполагаемых механизмах повышения всхожести семян под влиянием различных факторов, возможных механизмах потери всхожести семенами при старении. Обсуждается взаимосвязь между состоянием мембран семян и их всхожестью. Дан обзор методов, которые можно использовать для индивидуальной оценки качества семян.
II. Объекты и методы исследования
II.1. Объекты. Большая часть исследования проведена на семенах гороха (‘Немчиновский-85 и др.) и сои (‘Букурия’ и др.). Кроме того, в работе использовали семена пшеницы, ржи, ячменя (‘Рядовой’), огурцов (‘Московский тепличный’), перца, подсолнечника, кукурузы, фасоли и сосны.
II.2. Всхожесть семян определяли стандартным способом (правила ISTA – Международной ассоциации оценки качества семян, 1996). Под всхожестью партии понимают процент семян, из которых вырастают нормальные проростки. Не прорастающие семена и семена, из которых вырастают проростки с морфологическими дефектами, считаются невсхожими.
II.3. Выход электролитов из семян измеряли после 1,5 ч экспонирования индивидуального семени в 2 мл дистиллированной воды. Электропроводность среды определяли бесконтактным методом при помощи Oscillotitrator OK-302.
II.3. Действующие факторы:
II.3.1. Семена гороха подвергали воздействию g-излучения в Объединенном институте ядерных исследований (Дубна) на установке ROKUS-M при мощности дозы 0,913 Гр/мин на расстоянии 75 см от источника. Дозу подбирали, варьируя время облучения (1 Гр – 65,78 с). Для облучения семян в дозе 190 мГр использовали источник g-квантов 60Со (Е37) P0=0.362 Р/час/м (мощность дозы облучения – 5,7 мГр/час, расстояние 80 см, время облучения – 1/3 часа). Были выбраны 3 уровня доз: сверхмалая доза 190 мГр (использована для выявления ранних цитогенетических эффектов [Корогодина и др., 2004]), стимулирующие дозы для наблюдения радиационного гормезиса 3-10 Гр, и летальная доза 100 Гр.
II.3.2. Тепловую обработку семян проводили двумя способами. 1. Прогревание при 400С и 85% относительной влажности воздуха (так называемое «ускоренное старение»). 2. Прогревание при 40ОС герметично упакованных семян, влажность которых предварительно была увеличена до фиксированного значения («контролируемое повреждение»).
II.3.3. Лазерное облучение элитных семян огурцов проводили на вращающемся зеркальном диске в сухом затемненном помещении при температуре 20-250С. Использовали гелий-неоновый лазер ЛГ-75 (l=632,5 плотностью мощ-ности 0,3 мВт/см2) дозами 100-150 импульсов (1 импульс 50 мкДж/см2). (Облучен-ные семена были предоставлены ).
II.3.4. Звуковую обработку выборки семян ячменя (от 100 до 500 шт) про-водили в чашке Петри в течение 5 минут с помощью источника акустических волн (Звуковой генератор) мощностью 65 дБел с регулируемой частотой (с точностью 1 Гц). (Озвученные семена были предоставлены ).
II.3.5. Семена овса, подвергнутые светоимпульсному облучению (0,5 с, 50 МВт), и семена пшеницы после обработки электрическим полем коронного раз-ряда (0,5 с, 2кВт/см2) с целью повышения их всхожести были получены из Всесо-юзного НИИ экспериментальной физики (г. Саров).
II.4. Влажность семян определяли весовым способом по правилам ISTA [1996]. Сухие семена размалывали, быстро формировали из муки семян три навески по ~5 г и помещали в сушильный шкаф с 1050С на 3-5 часов до достижения постоянного веса. В дальнейшем закрытые бюксы с семенами охлаждали при комнатной температуре. После остывания семена взвешивали и рассчитывали влажность, как изменение веса после подсушивания, отнесенное к исходному весу.
II.5. Поглощение кислорода индивидуальными семенами или выделенными зародышевыми осями определяли полярографически при 20ОС электродом типа Кларка Электрод E5047, фирмы Radiometer A/S Denmark. Диаметр платинового электрода 20 мкм. Зародышевые оси инкубировали в камере с водой объемом 60 мкл. После того, как при дыхании зародыша или зародышевых соей концентрация кислорода в воде снижалась до нуля, камеру вновь заполняли водой и регистрировали дыхание. Процедуру повторяли 4-4 раза.
II.6. Анализ содержания ядерной ДНК проводили стандартным методом [Redfearn et al., 1995] в нашей модификации. Для анализа 2-мм отрезки апикальной части зародышевой оси растирали в буфере для выделения (0,2 М маннит, 10 мМ Мес-буфер, 10 мМ NaCl, 10 мM спермин-тетрагидрохлорид, 2,5 мМ Na2-ЭДТА, 0,05 об/об Тритон Х-100, рН 5,8). В суспензию добавляли 10 мкл 5 мг/мл этидиум бромида – специфического флуоресцентного (ФЛ) красителя ДНК, уровень ФЛ которого пропорционален количеству ДНК в клетке (2С, 4С, 8С). ФЛ регистрировали на микрофлуорометрическом анализаторе. 10 мкл суспензии помещали на предметное стекло люминесцентного микроскопа (ЛЮМАМ 13) с видеокамерой (QX3, Intel, США). Сигнал анализировали с помощью компьютерной программы, разработанной и . Интенсивность ФЛ измеряли у 600-800 ядер (25-80 ядер на каждом стекле) для каждой экспериментальной точки.
II.7. Определение состояния ДНК. Пятнадцать пятимиллиметровых кончиков зародышевых осей размалывали в жидком азоте с лизирующим раствором (50 мМ Трис-HCl буфер (рН 7,5), 25 мМ ЭДТА, 1% СДС). Смесь инкубировали 30 минут при комнатной температуре. После добавления NaCl до 1 М концентрации смесь была депротеинизирована встряхиванием с хлороформ/спиртовым раствором (10/1 об/об). После центрифугирования в течение 10 минут при 5000 g, ДНК была выделена из жидкой фазы добавлением тройного объема этанола (96%) и растворена в 50 мМ Трис-HCl-буфере (рН 7,5), содержащем 25 мМ ЭДТА. Образцы ДНК были обработаны рибонуклеазой А, не содержащей ДНКазы (50 мкМ/мл) в течение 20 мин при 370С и затем ДНК снова осаждена добавлением тройного объема этанола (96%). Одинаковые объемы изолированной и очищенной ДНК были электрофоретически разделены в течение 2 часов в 1,2% агарозном геле при напряжении 2,3 В/см в 0,09 М Трис-боратном буфере (рН 8,3), содержащем 0,5 мкМ/мл этидиум бромида (эту работу проводили совместно с лабораторией чл-корр. , НИИ физико-химической биологии им. , МГУ).
II.8. Хемилюминесценцию эмбриональных осей семян гороха измеряли в присутствии 5.10-5М хемилюминесцентного индикатора люминола. Свечение регистрировали на хемилюминометре с одноэлектронным счетом фотонов. Сигнал от фотоумножителя (ФЭУ-85), чувствительного в видимой области спектра, поступал на усилитель, а затем на самописец. Образцы во время измерения находились в термостатируемой камере. Растворы каталазы (2000 U/мг, Сигма, США) и антиоксидантов (пропилгаллат [10-4М] и b-меркаптоэтиламина [10-3М] использовали в качестве ингибиторов активных форм кислорода.
II.9. Термохемилюминесценцию регистрировали на этом же хемилюминометре, сигнал с которого поступал на компьютер.
II.10. Фосфоресценцию при комнатной температуре воздушно-сухих и набухающих семян регистрировали на установке с двухдисковым фосфороскопом. Объект освещали импульсами видимого света (6 мс свет, 24 мс темнота) от галогенной лампы КГМ-150. Послесвечение регистрировали в интервале 3-18 мс после прекращения освещения в видимой части спектра. Сигнал с фотоумножителя поступал сначала на усилитель (рН-340), а затем на самописец. Кинетику затухания свечения регистрировали в миллисекундной временной области.
II.11. Содержание глюкозы в семенах гороха оценивали двумя независимыми методами: глюкометром (Gluco care, Венгрия) и по уровню термохемилюминесценции при 150ОС (метод разработан нами).
II.12. Интенсивность синтеза белка определяли по включению меченого [35S]метионина в белки осевых органов семян гороха в течение 3-х часов набухания согласно рекомендации [Гумилевская и др., 1996].
Повторность опытов 3-5-кратная. Статистическую обработку результатов при оценке средних значений по выборке проводили с помощью программ статистической обработки данных: определяли средние значения, среднее квадратичное отклонение и дисперсию; использовали корреляционный анализ.
III. Результаты и обсуждение
Качество семян при хранении ухудшается. Из состарившихся семян наряду с нормальными проростками вырастают проростки с морфологическими дефектами (ненормальные), а часть живых семян, как и мертвые, не наклевываются вовсе [Isoly, 1957]. В период прорастания семена особенно чувствительны к внешним факторам, и потому их всхожесть в большой степени зависит от условий набухания, в частности от скорости поступления воды в семена и температуры [Priestley, 1986; Vertucci, 1989]. Старые семена больше подвержены повреждениям по сравнению с молодыми Vertucci, Farant, 1995; Obroucheva, 1999]. Быстрое поступление воды в клетки сухого зародыша может механически разрушить клетки [Larsson, 1968; Hoekstra et al., 2001].
При набухании у семян возникает кратковременная гипоксия, которую семена обычно благополучно переживают [Джеймс, 1956]. Причиной гипоксия у крупных семян бобовых может быть высокая скорость поглощения кислорода зародышем и медленная диффузия газа внутрь семени через семенную оболочку, поскольку семенная кожура препятствует поступлению к зародышу достаточного количества атмосферного кислорода, особенно в избыточном количестве воды [Crawford, 1977; Duke, Kakefuda, 1981; Rolletschek et al., 2002].
Вода поступает в клетки семян путем диффузии через липидный бислой и по специальным каналам, образуемым белками аквапоринами [Maurel et al., 1995, 1997; Schuurmans et al., 2003]. Они формируются на поздних стадиях созревания и регулируют поступление воды в клетки семян при набухании [Johansson et al., 1998; Chrispeels et al., 1999]. Скорость водного транспорта через каналы, образуемые аквапоринами, зависит от их количества и состояния (открыты или закрыты). В фосфорилированном состоянии каналы «открыты» и закрываются при дефосфорилировании, которая осуществляет фосфатаза. Инактивация фосфатазы не дает каналам закрываться [Maurel et. al., 1995; Johansson et. al., 1998]. Препараты ртути (парахлормеркурий бензоат и HgCl2) являются специфическими ингибиторами аквапоринов. Они связываются с цистеином-187 белка аквапорина, стерически закрывают канал, и тормозят поступление воды в клетки. Состояние каналов восстанавливается при отмывании семян восстановителем тиоловых групп дитиотрейэтолом [Maurel, 1997; Javot, Maurel, 2002]. На участие аквапоринов в набухании семян указывает малая зависимость этого процесса от температуры (энергия активации прохождения воды через аквапорины меньше 5 ккал/М, а диффузии через липидный бислой - 12-14 ккал/M) [Maurel, 1997].
Однако изучение взаимодействия этих факторов, определяющих в совокупности состояние семян при их хранении и при разных воздействиях требует разработки специального метода, который мог бы давать интегральную оценку состояния семени на всех этапах хранения и при прорастании в режиме реального времени.
III.1. Разработка методов оценки качества
индивидуальных семян
Популяция хранящихся семян содержит семена, из которых вырастают нормальные проростки (они определяют всхожесть партии), проростки с морфологическими дефектами (ненормальные проростки, которые по ГОСТу не считаются всхожими), и мертвые.
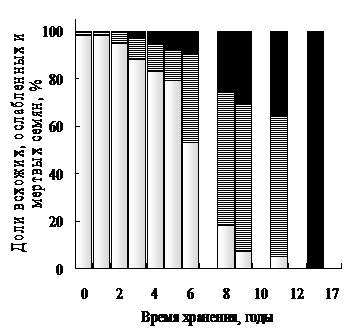
Во время хранения семена стареют, и качественный состав партии меняется (рис. 1). Вначале уменьшение числа всхожих семян обусловлено увеличением числа ослабленных, из которых вырастают проростки с морфологическими дефектами. Очевидно, что повысить всхожесть партии семян можно только за счет воздействия на ослабленные семена. Для изучения механизма «улучшения» такие семена необходимо отобрать из партии еще до проращивания. К началу нашей работы не существовало методов, которые позволили ли бы это делать.
|
О качестве семян обычно судят по качеству вырастающих из них проростков, а жизнеспособность определяют окрашиванием набухших семян витальными красителями и по выходу электролитов в дистиллированную воду. Каждую из процедур можно проводить только один раз, поэтому для выяснения изменений качества семян при хранении используют популяционно-статистический подход. Мы разработали метод неповреждающего контроля качества индиивидуальных сухих семян.
III.1.1. Качество воздушно-сухих семян. Воздушно-сухие семена после освещения видимым светом обладают длительным послесвечением, которое затухает в течение многих минут [Веселова и др., 1985а, б]. Кинетика затухания свечения многокомпонентная. При регистрации свечения с помощью фосфороскопа в миллисекундной области, оно, в основном, представлено двумя компонентами со временами жизни 1-3 и 12-20 мс.
Известно, что длительное свечение различных органических веществ, характеризуется свойствами, включающее линейную зависимость от интенсивности возбуждающего света, снижение при повышении температуры, в присутствии кислорода и повышении влажности, активируемое в присутствии ионов тяжелых металлов. Оно является фосфоресценцией с триплетного уровня при комнатной температуре [Parker et al., 1980, a, b]. Полученные нами характеристики свечения воздушно-сухих семян оказались сходными с таковыми для фосфоресценции органических соединений при комнатной температуре. На основании этого мы пришли к выводу, что свечение семян также является фосфоресценцией при комнатной температуре (ФКТ). Измеренный нами спектр свечения ФКТ воздушно-сухих семян широкий, неструктурированный (рис. 2, кривая 1) и, скорее всего, представляет собой сумму спектров свечения различных веществ: продуктов распада хлорофилла –
|
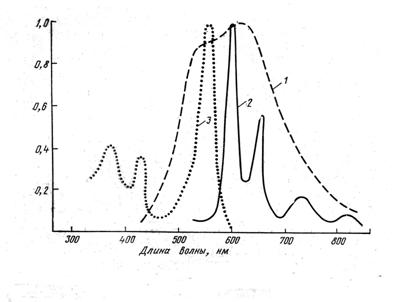 Рис. 2. Спектр ФКТ воздушно-сухих семян (1) и спектры излучения (2) и возбуждения (3) свечения набухающих семян гороха.
Рис. 2. Спектр ФКТ воздушно-сухих семян (1) и спектры излучения (2) и возбуждения (3) свечения набухающих семян гороха.порфиринов, целлюлозы, и флавинов, которых много в семенах. Оказалось, что характер спектра свечения меняется в зависимости от состояния семян. В отличие от широкого неструктурирован-ного спектра ФКТ сухих семян, спектр свечения набухающих семян имеет четыре четко выраженных максимума, характерных для спектры фосфоресценции не содержа-щих металла порфирина. То есть свечение набухающих семян обусловлено присутствием в них порфирина не содержащего метала.
|
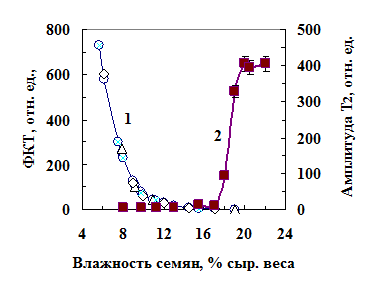 Рис. 3. Уровень ФКТ семян фасоли (кружки), пшеницы (треугольники), гороха (ромбы) (1) и амплитуды Т2 сигнала ЯМР (2) при разном содержании воды в семенах.
Рис. 3. Уровень ФКТ семян фасоли (кружки), пшеницы (треугольники), гороха (ромбы) (1) и амплитуды Т2 сигнала ЯМР (2) при разном содержании воды в семенах.Фосфоресценцию биополимеров наблюдают при криогенных температурах. При температуре выше 170 К фосфоресценция быстро снижается. Тушителем фосфорес-ценции является кислород [Гиллет, 1988]. Фосфоресценцию сухих семян при комнатной температуре можно наблюдать, потому что в них практически отсутствует кислород. При увлажнении семян диффузия кислорода в семя возрастает и фосфоресценция снижается. На рис. 3 кривой 1 показано, как с увеличением содержания влаги в семенах снижается ФКТ (коэффициент корреляции –0,96- -0,98). Увлажнение семян до 18-20% приводит к полному исчезновению ФКТ. При влажности семян 18-20%, судя по резкому в возрастанию амплитуды Т2 сигнала ЯМР, в семенах появляется свободная вода (рис. 3,кривая 2).
На основании зависимости ФКТ биопрепаратов и семян от влажности нами был предложен чувствительный метод оценки влажности этих объектов [Авт. свид. № 1 1981]. Фосфоресцентным методом можно определить разницу в содержании воды до 0,1-0,2% при общей влажности от 6 до 20%. Другие инструментальные методы имеют близкую чувствительность, но при влажности объектов выше 30-40%.
|
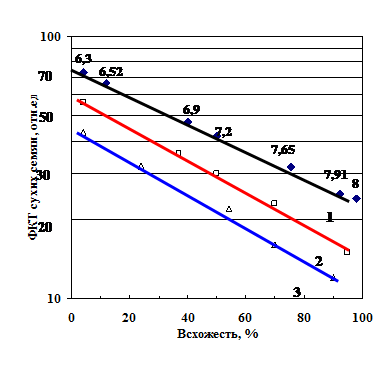
При хранении (старении) сухих семян разных видов параллельно со снижением всхожести возраста-ет уровень их ФКТ (рис. 4). Коэффициент корреляции между всхожестью и уровнем ФКТ составляет -0,94 - -0,98.
Рис. 4. Соотношение между всхожестью семян сои (1), ржи (2) и пшеницы (3) и их уровнем ФКТ. Числами около верхней линии указана влажность семян сои соответствующей всхожести.
Поэтому, было предложе-но по уровню фосфоресцен-ции семян судить о всхожести партии [Авт. свид. № 1 1982].
Известно, что обезвоживание характерно для биополимеров и жизнедеятельных организмов при их старении [Воюцкий, 1960; Серами и др., 1987; Растинг, 1993]. В процессе старения семян и их гибели содержание них воды также снижается (числа около прямой 1 на рис. 4). Вода в воздушно-сухих семенах является, в основном, связанной [Аксенов, 2006]. А в процессе гибели содержание воды уменьшается на 1,5-2%, (т. е. теряется пятая часть связанной воды). Известно, что такая потеря воды отражает необратимые перестройки макромолекул, сопровождающиеся уменьшением их водоудерживающей способности [Библь, 1963; Levitt, 1972; Голдовский, 1986].
Распределение сухих семян по уровню ФКТ (фракции). Методом ФКТ можно проводить измерения без нарушения целостности семян, что дает возможность периодически контролировать их влажность в процессе хранения. Вследствие высокой чувствительности ФКТ метода можно регистрировать сигнал от отдельных семян и анализировать состав популяции. На рисунке 5 показаны распределения по уровню ФКТ семян из партий разной всхожести. Распределение семян гороха в партии с 98-%-ной всхожестью выглядит близким к нормальному. В партии со всхожестью 72% распределение имеет два максимума, а у семян с 50%-ной всхожестью - три. Тесная взаимосвязь между уровнем свечения и влажностью семян, позволила считать, что их распределение в партии по уровню ФКТ отражает распределение семян по влажности. Эта закономерность легла в основу анализа гетерогенности партии семян по влажности.
Рис. 5. Распределение семян гороха в партиях с разной всхожестью по уровню ФКТ. Римскими цифрами обозначены номера фракций. |
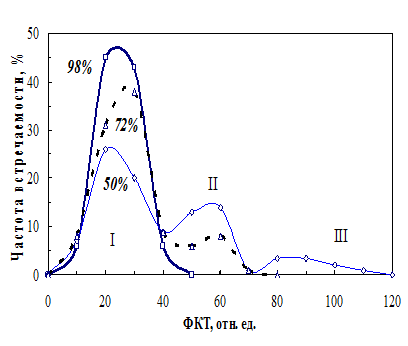
Средняя влажность семян в партии 72%-ной всхожести – 9,52%. Однако определе-ние влажности семян фракции I, отобранных из этой партии, как и влажность семян партии 98%-ной всхожести состав-ляла 9,84%. Влажность семян фракции II (уровень ФКТ 50-60 отн. ед.) – 8,90%, а фракции III, отобранной из партии 50%-ной всхожести (уровень 80-110 отн. ед.) - 8,2%. Т. е. в партиях семян пониженной всхожести семена могут значительно отличаться по содержанию воды, т. е. увеличивается гетерогенность семян в партии. Усреднение влажности партии семян 50%-ной всхожести по всем обнаруженным фракциям с учетом их долевого вклада дает значение среднее влажности при обычном определении у нефракционированной партии 9,2%.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
