

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Институт прикладной математики им. РАН, Москва
*****@***ru
БИОЛОГИЧЕСКИ ПРАВДОПОДОБНЫЙ
НЕЙРОСЕТЕВОЙ ДЕТЕКТОР КРАЯ
Аннотация
На основе физиологических данных об обработке зрительной информации в сетчатке глаза, латеральном коленчатом теле и первичной зрительной коре млекопитающих предлагается нейросетевой детектор края.
В системах машинного зрения важную роль играет выделение краев. Однако ни один из известных детекторов края не является совершенным. Принята следующая классификация ошибок: потеря настоящих краев, появление ложных краев, искажение формы и положения краев. Широко распространен подход, при котором края находят путем сглаживания и последующего дифференцирования изображения. Из-за чувствительности к шумам дифференцирование является некорректно поставленной задачей, а сглаживание изображения (регуляризация) приводит к потере информации [1].
Описываемый детектор края опирается не столько на математический аппарат, сколько на данные о функционировании мозга. Такой подход может привести к алгоритмам с очень хорошими характеристиками.
Свет воспринимается специальными фоторецепторами, колбочками и палочками, содержащими зрительные пигменты. Сигналы от фоторецепторов, посредством биполярных, горизонтальных и амакриновых клеток достигают ганглиозных клеток. Аксоны ганглиозных клеток проходят в составе зрительного нерва и являются единственными нервными волокнами, несущими зрительную информацию от глаза в мозг. Волокна зрительного нерва заканчиваются в области латерального коленчатого тела, где расположены вторые нейроны цепочки зрительного тракта. Нейроны коленчатого тела, в свою очередь, проецируются в первичную зрительную кору V1 [2].
Все эти связи имеют ретинотопическую организацию: окружающий нас мир (его оптическое изображение на сетчатке) упорядоченно отображается в различных областях зрительной системы в виде пространственного распределения возбужденных нейронов [3].
Рецептивным полем ганглиозной клетки называют область сетчатки, световая стимуляция которой может повлиять на импульсацию данной клетки. По устройству рецептивных полей можно выделить клетки с on-центром и клетки с off-центром. В обоих случаях рецептивные поля представляют собой круг и прилегающее к нему кольцо. Круг называют центром, а кольцо - периферией рецептивного поля.
Клетки с on-центром в темноте или при равномерном освещении их рецептивных полей проявляют спонтанную активность. При освещении небольшого пятна в центре рецептивного поля on-клетка разряжается с повышенной частотой. Наиболее сильный ответ получается в том случае, когда светлое пятно занимает весь центр. При освещении периферии рецептивного поля активность on-клетки, напротив, подавляется.
Поведение клетки с off-центром противоположно: освещение периферии увеличивает ее активность, а освещение центра – подавляет. Клетки обоих типов перемешаны и встречаются одинаково часто. Рецептивные поля соседних клеток частично перекрываются.
Ответы клеток латерального коленчатого тела не имеют разительных отличий от сигналов ганглиозных клеток. Они также имеют концентрически организованные рецептивные поля on - или off-типа, но механизм контраста отрегулирован тоньше, за счет большего соответствия между тормозными и возбуждающими зонами [2].
В первичной зрительной коре V1 содержатся (среди многих других) клетки, для которых наилучшим стимулом является граница между светлым и темным, проходящая в фиксированном направлении. Такая клетка совсем не реагирует на границы, перпендикулярные оптимальному направлению, и на рассеянный свет. Она дает несколько более слабый ответ на границы, проходящие под небольшим углом к оптимальному направлению.
Клетки с ориентационной чувствительностью сгруппированы в микроколонки. В микроколонке представлены все возможные ориентации на данном участке зрительного поля. Рецептивные поля соседних микроколонок частично перекрываются.
Равномерной решеткой на прямоугольнике
![]()
называется множество точек ![]() , где
, где ![]() ,
, ![]() ,
, ![]() ,
, 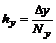 .
.
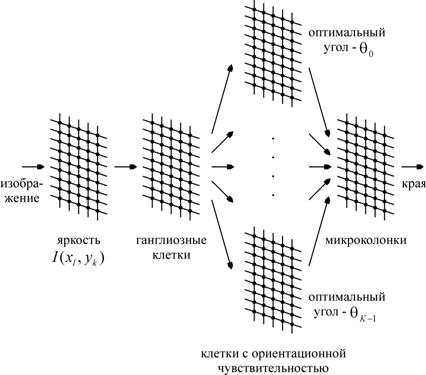
Рис. 1. Общая схема биологически правдоподобного детектора края
Общая схема нейросетевого детектора края представлена на рис. 1. Исходное изображение задано в виде значений яркости ![]() в узлах
в узлах ![]() равномерной решетки. Яркость может принимать значения от 0 (черный) до 1 (белый). Искусственные нейроны, имитирующие активность ганглиозных клеток, клеток с ориентационной чувствительностью и микроколонок, расположены в узлах точно таких же решеток.
равномерной решетки. Яркость может принимать значения от 0 (черный) до 1 (белый). Искусственные нейроны, имитирующие активность ганглиозных клеток, клеток с ориентационной чувствительностью и микроколонок, расположены в узлах точно таких же решеток.
Построим аналог ганглиозной on-клетки. Скалярный выход ![]() искусственного нейрона, расположенного в узле
искусственного нейрона, расположенного в узле ![]() , определим как свертку весовой функции
, определим как свертку весовой функции ![]() со значениями яркости:
со значениями яркости: ![]() . Положим весовую функцию кусочно-постоянной, в соответствии с устройством рецептивного поля on-клетки:
. Положим весовую функцию кусочно-постоянной, в соответствии с устройством рецептивного поля on-клетки:
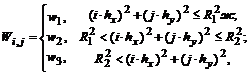
где ![]() – радиус центра рецептивного поля,
– радиус центра рецептивного поля, ![]() – внешний радиус периферии рецептивного поля,
– внешний радиус периферии рецептивного поля, ![]() ,
, ![]() и
и ![]() – постоянные веса.
– постоянные веса.
Поскольку световая стимуляция за пределами рецептивного поля не может влиять на ответ on-клетки, имеем ![]() . Потребуем, чтобы спонтанной активности on-клетки соответствовало
. Потребуем, чтобы спонтанной активности on-клетки соответствовало ![]() . Это означает, что при
. Это означает, что при ![]() (яркость одинакова во всех узлах)
(яркость одинакова во всех узлах)
![]() ,
,
где ![]() – число узлов, попавших в центр рецептивного поля (удовлетворяющих неравенству
– число узлов, попавших в центр рецептивного поля (удовлетворяющих неравенству ![]() ), а
), а ![]() – число узлов, попавших в периферию рецептивного поля (удовлетворяющих неравенству
– число узлов, попавших в периферию рецептивного поля (удовлетворяющих неравенству ![]() ). Отсюда
). Отсюда ![]() .
.
Световая стимуляция в центре увеличивает активность on-клетки, следовательно ![]() . Световая стимуляция на периферии уменьшает активность on-клетки, следовательно
. Световая стимуляция на периферии уменьшает активность on-клетки, следовательно ![]() . Тогда максимально возможный ответ
. Тогда максимально возможный ответ ![]() модели on-клетки достигается при
модели on-клетки достигается при
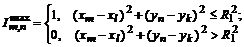
(пятно максимальной яркости полностью занимает центр рецептивного поля): ![]() . Минимально возможный ответ
. Минимально возможный ответ ![]() модели on-клетки достигается при
модели on-клетки достигается при
![]()
(пятно максимальной яркости полностью занимает периферию рецептивного поля): ![]() . Введем условие нормировки:
. Введем условие нормировки: ![]() . Отсюда
. Отсюда ![]() .
.
С учетом того, что 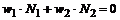 имеем:
имеем:
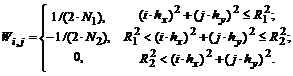
Свертка яркости с этой весовой функцией может быть найдена как половина разности средних значений яркости по центру и по периферии рецептивного поля.
Аналогичным образом построенная модель off-клетки имеет весовую функцию
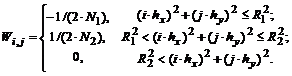
Скалярный выход ![]() искусственного аналога off-клетки отличается от выхода on-клетки только знаком:
искусственного аналога off-клетки отличается от выхода on-клетки только знаком: ![]() .
.
Поскольку особое внимание в этой работе уделяется обнаружению краев, введем следующее понятие. Идеальным краем называется такая прямая ![]() , что яркость
, что яркость
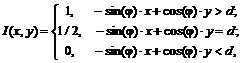
где ![]() – угол наклона прямой,
– угол наклона прямой, ![]() – расстояние от прямой до начала координат. На рис. 2 применительно к краям сравниваются ответы ганглиозных клеток сетчатки и описанной выше модели on-клетки. Из соображений подобия графиков (и, следовательно, соответствия физиологическим данным) выбрано
– расстояние от прямой до начала координат. На рис. 2 применительно к краям сравниваются ответы ганглиозных клеток сетчатки и описанной выше модели on-клетки. Из соображений подобия графиков (и, следовательно, соответствия физиологическим данным) выбрано ![]() .
.
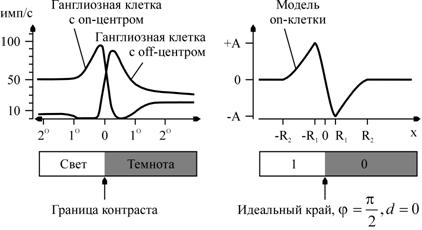
Рис. 2. Слева – ответы on - и off - клеток, построенные по экспериментальным данным [3]; справа – ответы модели on-клетки. По оси абсцисс отложено смещение центра рецептивного поля клетки относительно границы светлого и темного
Построим аналог клетки из первичной зрительной коры V1, настроенной на край, проходящий под углом ![]() . По аналогии с работой мозга, входной информацией для модельной клетки является активность
. По аналогии с работой мозга, входной информацией для модельной клетки является активность ![]() on-клеток и
on-клеток и ![]() off-клеток.
off-клеток.
Распределение активности on-клеток на идеальном крае представлено на рис. 2. Реальный край может быть размытым, иметь меньший контраст, быть зашумлен. Размытость приводит к увеличению расстояния между максимальным и минимальным ответами и к уменьшению их амплитуды ![]() . Уменьшение контраста – к пропорциональному уменьшению амплитуды. При малых значениях
. Уменьшение контраста – к пропорциональному уменьшению амплитуды. При малых значениях ![]() подавление шума не происходит. В общем случае можно лишь предположить, что в среднем слева от края положительными будут
подавление шума не происходит. В общем случае можно лишь предположить, что в среднем слева от края положительными будут ![]() , а справа –
, а справа – ![]() . Иначе говоря, для узла
. Иначе говоря, для узла ![]() через который проходит край с параметром
через который проходит край с параметром ![]() будет выполнено
будет выполнено ![]() ,
, ![]() , где
, где ![]() и
и ![]() определяют прямоугольники, по узлам внутри которых производится суммирование, а
определяют прямоугольники, по узлам внутри которых производится суммирование, а ![]() – число слагаемых в сумме.
– число слагаемых в сумме.
Необходимо, чтобы по мере отклонения угла наклона края ![]() от угла
от угла ![]() , на который настроена клетка, ответ
, на который настроена клетка, ответ ![]() модельной клетки убывал. Необходимо, чтобы по мере удаления края от узла
модельной клетки убывал. Необходимо, чтобы по мере удаления края от узла ![]() ответ
ответ ![]() быстро убывал. Этим требованиям удовлетворяет функция
быстро убывал. Этим требованиям удовлетворяет функция 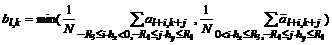 . Внутри однородных областей и когда край перпендикулярен оптимальному направлению имеем
. Внутри однородных областей и когда край перпендикулярен оптимальному направлению имеем ![]() .
.
Клетка в узле ![]() , настроенная на угол
, настроенная на угол ![]() отличается от описанной выше поворотом прямоугольников, по узлам внутри которых производится суммирование, вокруг точки
отличается от описанной выше поворотом прямоугольников, по узлам внутри которых производится суммирование, вокруг точки ![]() на угол
на угол ![]() .
.
Клетки с ориентационной чувствительностью, расположенные в одном и том же узле ![]() , но настроенные на разные ориентации края
, но настроенные на разные ориентации края ![]() , сгруппированы в микроколонки. За ответ целой микроколонки примем максимальный из ответов входящих в нее клеток:
, сгруппированы в микроколонки. За ответ целой микроколонки примем максимальный из ответов входящих в нее клеток: 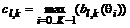 . Тогда высокая активность микроколонки означает наличие края, проходящего близко к центру ее рецептивного поля.
. Тогда высокая активность микроколонки означает наличие края, проходящего близко к центру ее рецептивного поля.
Работа описанного детектора края продемонстрирована на рис. 3.


Рис. 3. Слева – матрица яркости, 192x168 узлов. Справа – результат выделения краев с параметрами ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() . Узел закрашивался в черный, если активность микроколонки в нем превосходила
. Узел закрашивался в черный, если активность микроколонки в нем превосходила 
Список литературы
1. Ziou D. and Tabbone S. Edge Detection Techniques - An Overview // Pattern Recognition and Image Analysis. 1998. Vol 8. № 4. P. 537–559. http://www. loria. fr/~tabbone/PAPERS/EDsurvey. pdf
2. Николлс Дж. Г., , Дж., От нейрона к мозгу. М.: Едиториал УРСС, 2003.
3. Физиология человека / Под ред. Р. Шмидта и Г. Тевса. 2-е изд., перераб. и доп. М.: МИР, 1996.
