

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
У 130 детей, проживающих на экспонированной территории в возрасте 5-9 лет, были проведены анализы на содержание фтора в моче. У 75 детей (57,7%) было обнаружено повышенное содержание фтора в моче, уровень которого составил от 1,03 до 3,0 мг/л (норма от 0,4 до 1,0 мг/л.). Из этих детей у 31 ребенка было проведено рентгенологическое исследование костей кисти. Рентгенологическая норма выявлена у 1 ребенка (3%). У 30 детей выявлен ряд рентгенологические симптомы. Наличие псевдоэпифизов во II и в V пястных костях; отсутствие ядер окостенения трапециевидной и полулунной костей; дистального отдела локтевых костей и костей запястья; истончение кортикального слоя; очаговое склерозирование диафизов проксимальных фаланг, а также эпифизов пястных костей; слабая функциональная оформленность костей запястья; асимметрия ядер окостенения локтевой кости и трапециевидных костей; отставание в размерах ядер окостенения костей запястья (разница в числе) и т. д. По данным -Строгановой (1957), (1964) подобные симптомы рентгенологических отклонений от нормы могут наблюдаться при токсическом воздействии соединений фтора. Таким образом, на опытной территории у большинства детей в моче определяется повышенное содержание фтора, а рентгенологические симптомы нарушений развития костей кисти и дистального отдела костей предплечья более разнообразны, чем у детей на контрольной территории, и выявляются значительно чаще.
Таким образом, на основании полученных результатов можно сделать ряд выводов:
- на опытной территории нарушения в развитии и росте костей наиболее выражены в периоды детства у девочек (растущих более активно), а в подростковый период – у мальчиков (активно растущих в 9 и 15 лет);
- периоды активации роста костей совпадают с периодами наибольшей частоты встречаемости нарушений в развитии и росте костей, т. е. костные ткани больше уязвимы при действии токсикантов в периоды активного роста;
- на контрольной территории нарушения в развитии и росте костей наименее выражены и диагностируются, преимущественно, у мальчиков, что, по-видимому, связано с характерными для них двумя ростовыми скачками.
Морфогенез костей у плодов человека на изучаемых территориях
Результаты гистологического исследования бедренных костей у плодов человека демонстрируют значительные отличия процесса остеогенеза на изучаемых территориях. На опытной территории у плодов 15-16 недель гестации все измеряемые параметры в бедренной кости, кроме толщины стенки диафиза и костной ткани, нарастающей на эпифиз, оказались меньше, чем у плодов на контрольной территории (табл.7). При одинаковой толщине стенки диафиза толщина ее костных балок оказалась меньше в 1,5 раза, а метаэпифизарная пластинка роста тоньше в 4 раза. Зоны пузырчатого и столбчатого хряща в пластинке роста вдвое меньше по толщине, а длина энхондральных балок в линии окостенения – в 5 раз меньше. Количество проросших в эпифиз сосудов было меньше в 3 раза.
На стадии 18-20 недель эмбриогенеза в бедренной кости толщина перихондральной костной манжетки, составляющей стенку диафиза, и ее костных балок уменьшается вдвое и становится меньше в 3 и 5 раз, соответственно. Толщина костной ткани, нарастающей на эпифизы, тоже уменьшается вдвое, параллельно уменьшается более чем в 2 раза и толщина зон пузырчатого и столбчатого хряща, хотя общая толщина метаэпифизарной пластинки роста незначительно увеличивается за счет возрастания в 1,7 раза длины энхондральных балок в линии окостенения. Активизируется прорастание сосудов в эпифизы бедренных костей, тем не менее, значительно отстает рост бедренных костей в длину и в толщину. Полученные данные позволяют говорить не только о меньшей активности остеобластов, но и предполагать гиперактивацию процессов резорбции костной ткани у плодов на экспонированной территории.
Таблица 7.
Морфометрические показатели бедренных костей у плодов (г. Шелехов)
Показатели | Возраст | |||
15-16нед. | 18-20нед. | 22-25нед. | 27-29нед. | |
Количество сосудов в эпифизе | 8,3±4,4* | 27,9±3,1* | 30±1,8* | 30,6±3,8* |
Толщина пузырчатого хряща (мкм) | 216±33,2* | 88,3±8,2* | 93,7±15,6* | 77,5±7* |
Толщина столбчатого хряща (мкм) | 330,5±52,3* | 150±13,3* | 172±13,1* | 178,5±15,5* |
Длина энхондральных балок (мкм) | 780±156,9* | 1343±159* | 1554±129* | 1765±143* |
Толщина костной ткани, нарастающей на эпифиз (мкм) | 14,5±1,7 | 6,7±1* | 4,6±0,6* | 4±0,00* |
Толщ. энхонд. балок, сросшихся со стенкой диафиза (мкм) | 38,5±8,5* | 28,3±2,9* | 41,1±8,9* | 41±4,9* |
Толщина стенки диафиза (мкм) | 274,8±25,8 | 135,3±6,4* | 132,6±17* | 102,5±9* |
Толщ. кост. балок стенки диафиза(мкм) | 51,5±7* | 22,3±1,9* | 23,4±2,4* | 21,5±2,4* |
Количество наблюдений | 8 | 12 | 7 | 8 |
Примечание: * - отличие показателя от данных г. Иркутска, p<0,05.
С 22 по 29 недели эмбрионального развития плодов на экспонированной территории изучаемые параметры бедренных костей существенно не изменяются. Толщина стенки диафиза и ее костных балок у плодов меньше, чем у плодов на контрольной территории, в 4 раза на 22-25 неделе развития, в 6-9 раз на 27-29 неделе (р<0,05). Толщина костной ткани, нарастающей на эпифизы, меньше в 3 раза (р<0,05), а энхондральные костные балки, сросшиеся со стенкой диафиза, тоньше в 3,7-5 раз (р<0,05). Из этих данных следует, что аппозиционный рост костей в толщину существенно заторможен, вероятно, по причине снижения синтетической активности остеобластов и повышения резорбционной активности остеокластов. Количество сосудов, проросших в эпифизарный хрящ меньше на 30-35 %, при этом толщина метаэпифизарной пластинки роста увеличивается очень медленно и только за счет удлинения энхондральных балок в линии окостенения, которые, тем не менее, остаются в 4,5 раза короче, чем у плодов г. Иркутска. Хрящевые зоны пластинки роста с 22 по 29 неделю развития практически не изменяются, толщина зоны пузырчатого хряща меньше, чем у плодов г. Иркутска, в 4,6-5,2 раза (р<0,05), а зоны столбчатого хряща – в 4,4-4,6 раза (р<0,05). На опытной территории происходит торможение роста костей в эмбриональный период в толщину и в длину.
Полученные результаты гистологического исследования показали, что в условиях техногенного загрязнения внешней среды фторидами существенно изменяется ход остеогенеза в период эмбрионального развития. Это выражается в повышении резорбирующей функции остеокластов и в торможении синтетической функции остеобластов, а также подавлении размножения столбчатого хряща пластинки роста, что приводит к торможению роста костей и в длину, и в толщину.
Анализ корреляционных связей между измеряемыми параметрами показал, что у плодов на контрольной территории преобладают положительные взаимосвязи (рис.6). Функциональная система, обеспечивающая рост кости, хорошо сбалансирована и устойчива, процессы остеогенеза и остеорезорбции сбалансированы и характеризуются значительным преобладанием активности остеобластов над активностью остеокластов, что обеспечивает интенсивный рост костей. При этом в процессе роста костей в плодный период выделяются два этапа: 1 этап - с 15 по 22 неделю бедренные кости более активно растут в длину, 2 этап - с 22 по 29 неделю рост костей в длину замедляется, а в толщину - активизируется. Ведущим индуктором остеогенеза в г. Иркутске является, по-видимому, кислород, высокая концентрация которого обеспечивается большим количеством прорастающих в эпифиз кровеносных сосудов.
Корреляционные связи между измеряемыми параметрами бедренных костей (рис.6) у плодов на опытной территории значительно отличаются.
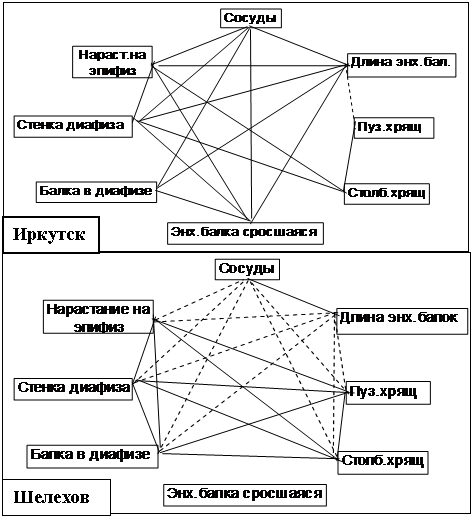 |
Рис. 6. Корреляционные взаимосвязи между морфометрическими показателями бедренной кости в период 15-29 недель эмбрионального развития плодов человека в городах Иркутск и Шелехов. Обозначения: сплошные линии – прямые связи, пунктир – обратные связи.
Обращает внимание изменение направленности взаимосвязей между количеством проросших в эпифиз сосудов и остальными показателями. Бедренные кости растут в длину и толщину тем активнее, чем меньше сосудов в эпифизе, т. е. кровоснабжение и оксигенация тканей растущей кости утрачивает роль индуктора остеогенеза. Аналогичный вывод был сделан нами и по данным экспериментального исследования. Изменение направленности взаимосвязей наблюдается между длиной энхондральных балок в линии окостенения и толщиной стенки диафиза и костных балок в ней. Это означает однонаправленное изменение активности остеокластов (разрушающих энхондральные балки) и остеобластов (надстраивающих стенку диафиза в толщину). При этом толщина стенки диафиза в процессе развития плода не только не увеличивается, но даже уменьшается, следовательно, резорбционная активность остеокластов выше, чем синтетическая активность остеобластов. В пользу этого предположения свидетельствует потеря всех взаимосвязей с толщиной участков срастания энхондральных балок с перихондральными у плодов на опытной территории.
На основании полученных результатов морфогенеза костей у плодов, выделен ряд закономерностей:
- на экспонированной территории у плодов функциональная система роста костей устойчива, но кислород - основной индуктор остеогенеза заменяется другими индукторами (влиянием костного матрикса и клетками микроокружения и т. п.). Более высокая плотность и химическая устойчивость фторапатита, химического соединения, откладывающегося в костной ткани, приводит, вероятно, к компенсаторному повышению активности остеокластов, которое наблюдается у плодов на опытной территории. Возможно, по этой же причине снижается активность размножения клеток зоны столбчатого хряща в пластинке роста, дифференцировка остеобластов и их синтетическая активность, что приводит к торможению энхондрального роста костей в длину и аппозиционного роста в толщину.
- выделены два этапа морфогенеза костей, на первом этапе (до 16 недель), остеосинтез преобладает над остеорезорбцией, бедренные кости более активно растут в толщину, а в длину рост заторможен. По-видимому, в первую половину эмбриогенеза в организме плода концентрация соединений фтора еще незначительна, и его влияние распространяется пока только на процессы метаболизма и пролиферативную активность клеток. На втором этапе (с 18 по 29 неделю) процессы остеорезорбции преобладают над остеосинтезом, толщина костей уменьшается, а рост костей в длину остается малоактивным. Вероятно, на этом этапе в организме плода накапливается значительное количество соединений фтора, которые образуют фторапатит и повышают минеральную насыщенность растущих костей, затрудняя остеорезорбцию в процессе их ремодуляции, и приводят к компенсаторной гиперактивации функций остеокластов
Постэмбриональный морфогенез костей у потомства крыс с хронической фтористой интоксикацией
У потомства, полученного от крыс с фтористой интоксикацией, морфометрические характеристики пястных и бедренных костей отличались от аналогичных характеристик у потомства от интактных животных (табл.8).
Таблица 8
Морфометрические показатели пястных костей у новорожденных
и полуторамесячных крысят
Показатели | Возраст 1сутки | Возраст 1,5 месяца | ||
Интактные (контроль) | Действие фтора (опыт) | Интактные (контроль) | Действие фтора (опыт) | |
Сосуды (V%) | 0,5±0,08 | 0,6±0,14 | 2,2±0,13 | 1,2±0,18* |
Толщина пузыр. хряща (мкм) | 117±16,8 | 117±10,4 | 301±10,6 | 282±13* |
Толщина столб. хряща (мкм) | 114±9 | 134±11,3 | 486±11,6 | 436±27* |
Длина энхондр. балок (мкм) | 1454±80 | 1385±150 | 533±40 | 641±50* |
Толщ. кост. стенки метафиза(мкм) | 4±1 | 6±0,1* | 6±0,3 | 5±0,3* |
Срастание энхондр. кости со стенкой диафиза (мкм) | 280±60 | 370±40 | 150±20 | 230±20* |
Толщина стенки диафиза(мкм) | 181±19 | 218±12* | 114±8 | 82±8* |
Толщина костных балок (мкм) | 34±3 | 41±2* | 20±1 | 25±2* |
Примечание: * - отличие показателя от контрольного, p<0,05, n=8.
У «новорожденных» подопытной группы прослеживалась тенденция к увеличению толщины стенки диафиза и костных балок, а толщина костной ткани в области метафиза и нарастания ее на эпифизы была больше в 1,5 раза. Толщина участков срастания энхондральной костной ткани со стенкой диафиза в опыте тоже была больше. Эти данные косвенно свидетельствуют о снижении функциональной активности остеокластов, разрушающих энхондральную костную ткань и участвующих в перестройке перихондральной костной ткани, что приводит к более активному росту пястных костей в толщину. Пластинка роста исследуемых костей, а также количество сосудов, проросших в нее по хрящевым каналам у новорожденных крысят одинаковы. Из этого следует, что в эмбриональный период в контроле и опыте кровоснабжение метаэпифизарной области и скорость роста костей в длину одинакова.
В течение последующих 1,5 месяцев жизни у крыс-«подростков» контрольной группы существенно усиливается кровоснабжение тканей формирующейся кости, количество сосудов увеличивается, по сравнению с новорожденными крысятами, в 4,3 раза (p<0,05). Вероятно, это связано с особенностями подросткового периода, который характеризуется активацией всех систем, участвующих в метаболизме. При этом толщина стенки диафиза, по сравнению с костями новорожденных, уменьшается в 1,6 раза, что связано, по-видимому, с перестройкой грубоволокнистой костной ткани в тонковолокнистую костную ткань. Общая толщина пластинки роста уменьшается к 1,5 месяцам в 1,3 раза, при этом толщина линии окостенения (длина энхондральных балок) уменьшается в 2,7 раза, а толщина хрящевых зон, наоборот, увеличивается: пузырчатый хрящ в 2,6 раза, столбчатый в 4,3 раза. Таким образом, можно говорить об ускорении роста кости за счет активного деления хондроцитов столбчатого хряща у животных контрольной группы. Существенное уменьшение длины энхондральных балок и участков срастания энхондральной кости с костной стенкой диафиза свидетельствует об активной работе остеокластов.
В опыте у полуторамесячных крыс кровоснабжение растущей кости тоже увеличилось, но оказалось в 2 раза хуже, чем в контроле. Толщина стенки диафиза уменьшилась в 2,6 раза, что на 30% меньше, чем в контроле. Из этого следует, что в процессе перестройки грубоволокнистой костной ткани в компактную костную ткань активность остеобластов существенно снизилась, вероятно, под влиянием кумуляции соединений фтора, полученных от матери в эмбриональный период. В пластинке роста толщина линии энхондрального окостенения уменьшилась в 2,7 раза (в контроле в 3,3 раза), толщина пузырчатого хряща увеличилась, по сравнению с новорожденными, так же как в контроле, а толщина столбчатого хряща возросла в меньшей степени – в 3,3 раза (в контроле – в 4,3 раза). Из этого следует, что рост кости и в толщину, и в длину в условиях опыта идет медленнее, чем в контроле. Необходимо отметить, что в условиях опыта в пястных костях полуторамесячных крыс начинают закладываться точки эпифизарного окостенения. У животных контрольной группы этого не наблюдалось.
Обсуждая представленные данные, следует учесть, что в процессе эмбриогенеза рост костей обеспечивается функциональной физиологической системой, в которой можно выделить три основных структурных компонента, отражающих эффективность работы этой системы: 1 - количество сосудов, обеспечивающих растущую кость кислородом и питанием, 2 – толщина стенки формирующейся кости (перихондральной манжетки), 3 – толщина пластинки роста.
Корреляционный анализ показал, что в контрольной группе у новорожденных крысят все компоненты физиологической системы формирования кости тесно взаимосвязаны (рис.7-А). При этом кровоснабжение играет ведущую роль в функциональной активности пластинки роста, что подтверждается прямыми связями между количеством сосудов, толщиной пузырчатого и столбчатого хряща, а также толщиной костной манжетки, нарастающей на эпифизы. Толщина стенки диафиза и костных балок связаны высокой обратной корреляцией с толщиной зоны пузырчатого хряща и длиной энхондральных балок, что говорит об активной работе остеокластов в пластинке роста. Из этих данных можно сделать заключение о том, что в контрольной группе у новорожденных формируется устойчивая функциональная физиологическая система, обеспечивающая активный рост кости. Она поддерживается десятью корреляционными взаимосвязями, из них шесть положительных и четыре отрицательных.
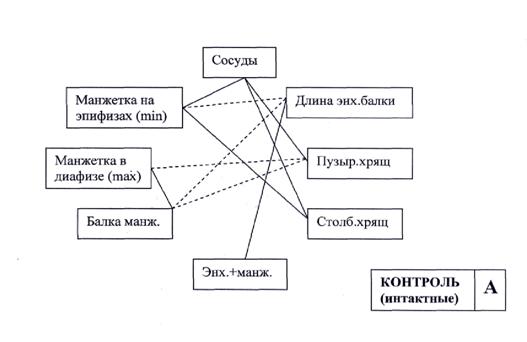
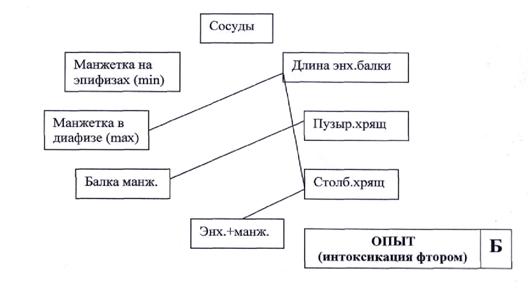
Рис. 7. Схема корреляционных связей между морфометрическими показателями пястной кости у новорожденных крысят контрольной (А) и подопытной (Б) групп.
В условиях опыта (рис.7-Б) количество корреляционных связей между составными компонентами функциональной системы сократилось до четырех. При этом количество сосудов в метаэпифизарной области не имеет взаимосвязей с другими структурными элементами формирующейся кости, следовательно, утрачивается ведущая роль снабжения кислородом тканей пластинки роста. Меняется направленность взаимосвязей между толщиной стенки диафиза и линией окостенения, что является подтверждением ослабления функциональной активности остеокластов у подопытных животных. Об этом же свидетельствует и прямая корреляция между толщиной костных балок и толщиной пузырчатого хряща. Толщина растущего столбчатого хряща обнаруживает прямую зависимость от длины энхондральных балок, в отличие от контрольных животных, у которых этот показатель зависит от количества сосудов. Учитывая, что толщина столбчатого хряща у новорожденных в контроле и опыте одинакова, можно предположить, что в функциональной системе подопытных животных изменяется индуктор роста кости, которым могут выступать биологически активные вещества, вырабатываемые остеобластами или другими клетками костномозгового микроокружения. Это предположение, безусловно, нуждается в тщательной проверке.
На основании представленных данных можно однозначно утверждать о том, что под действием избыточных концентраций соединений фтора на эмбрион у новорожденных формируется функциональная патологическая система, обеспечивающая рост кости. Характерными признаками этой системы являются, во-первых, ухудшение кровоснабжения пластинки роста кости, во-вторых, ослабление функциональной активности остеокластов, обеспечивающих перестройку линии окостенения и формирование стенки диафиза.
У полуторамесячных крысят в функциональной системе, обеспечивающей рост и формирование кости, наблюдаются существенные изменения, по сравнению с новорожденными. Они выражаются в разбалансировке составных компонентов и потере устойчивости системы и более выражены у подопытных животных. У крысят контрольной группы количество сосудов не обнаруживает устойчивых взаимосвязей с другими элементами функциональной системы кости, следовательно, несмотря на увеличение кровоснабжения, повышенная концентрация кислорода в костной ткани утрачивает свое ведущее значение в процессе остеогенеза. Толщина костной стенки диафиза связана прямой корреляцией с длиной энхондральных балок - свободных и сросшихся со стенкой кости. Учитывая, что значение этих показателей существенно уменьшилось по сравнению с «новорожденными», можно считать перестройку кости стенки диафиза и резорбцию энхондральной кости у животных контрольной группы активизированной. Толщина пузырчатого и столбчатого хряща тоже прочно связаны прямой корреляцией, а их значения существенно увеличены. Следовательно, рост кости в длину, по сравнению с новорожденными, ускорен. В целом функциональная система растущей кости у крыс-«подростков» контрольной группы характеризуется всего пятью прямыми взаимосвязями, что говорит о ее низкой устойчивости, которая, вероятно, предшествует последующему замедлению и прекращению роста костей скелета. Ее регуляция осуществляется, преимущественно, влиянием микроокружения.
В опыте у полуторамесячных крыс функциональная система, обеспечивающая рост кости, еще менее устойчива и обнаруживает всего три значимых корреляционных взаимосвязи. Количество сосудов прочно взаимосвязано прямой корреляцией с толщиной костной стенки диафиза, которая в свою очередь положительно связана с толщиной столбчатого хряща. Таким образом, в условиях опыта кровоснабжение определяет рост кости и в толщину, и в длину, играя ведущую индукторную роль (в отличие от контроля) на данном этапе остеогенеза. Учитывая, что в опыте количество сосудов вдвое меньше, чем в контроле, логично предположить, что именно недостаточность кровоснабжения и является основной причиной замедления роста кости.
Аналогичные закономерности выявлены и в развитии бедренной кости, в которой этапы остеогенеза проходят, по сравнению с пястными костями, ускоренно. Уже у новорожденных в бедренной кости в эпифизах обнаруживаются признаки дистрофии хондроцитов и формирование пузырчатого хряща, что указывает на развитие начального этапа эпифизарного окостенения. В возрасте 1,5 месяца у крыс окостенение эпифизов хорошо развито, эпифизарный хрящ почти полностью замещен костной тканью губчатого строения, в которой полости между костными балками заполнены красным костным мозгом. У интактных животных этого возраста метаэпифизарная пластинка роста истончается за счет сужения зоны столбчатого хряща, хотя окружающие его зоны пузырчатого хряща (со стороны диафиза и эпифиза) остаются относительно широкими. У подопытных животных, вероятно, под действием кумулированных в костях фтористых соединений, общая толщина пластинки роста, в том числе зоны столбчатого хряща, существенно больше, чем у интактных животных.
Основываясь на представленных данных, можно сделать следующее заключение.
Высокие концентрации соединений фтора, воздействующие на организм беременной крысы, оказывают негативное действие на формирование и рост скелета плода, путем проникновения через плаценту, накапливаясь в его тканях, вызывая существенные нарушения в формировании и росте костей у новорожденных и у 1,5-месячных крыс. Нарушения остеогенеза в эмбриональный период отражаются в разбалансировке у новорожденных функциональной системы роста кости и в замене индуктора остеогенеза, которым в норме является кровоснабжение и концентрация кислорода в тканях растущей кости, а в условиях опыта – предположительно, влияние микроокружения.
В постнатальном онтогенезе в переходный период, соответствующий предпубертатному периоду человека, закономерно снижается устойчивость функциональной системы роста костей. У интактных животных рост костей в толщину и в длину значительно более активен и обеспечивается преимущественно взаимодействием клеток микроокружения, т. е. локальными механизмами. В условиях интоксикации соединениями фтора локальные механизмы ауторегуляции оказываются недостаточными и на первый план выходят центральные механизмы в частности кровоснабжение.
На основании проведенного комплексного исследования по выявлению закономерностей морфогенеза костей в условиях влияния загрязнения окружающей среды соединениями фтора, составлена концептуальная схема, представленная на (рис. 8). Из которой видно, что у населения, проживающего в зоне размещения предприятий по производству алюминия, фтористые соединения окружающей среды оказывают токсическое действие на процессы морфогенеза костей на всех этапах онтогенеза.
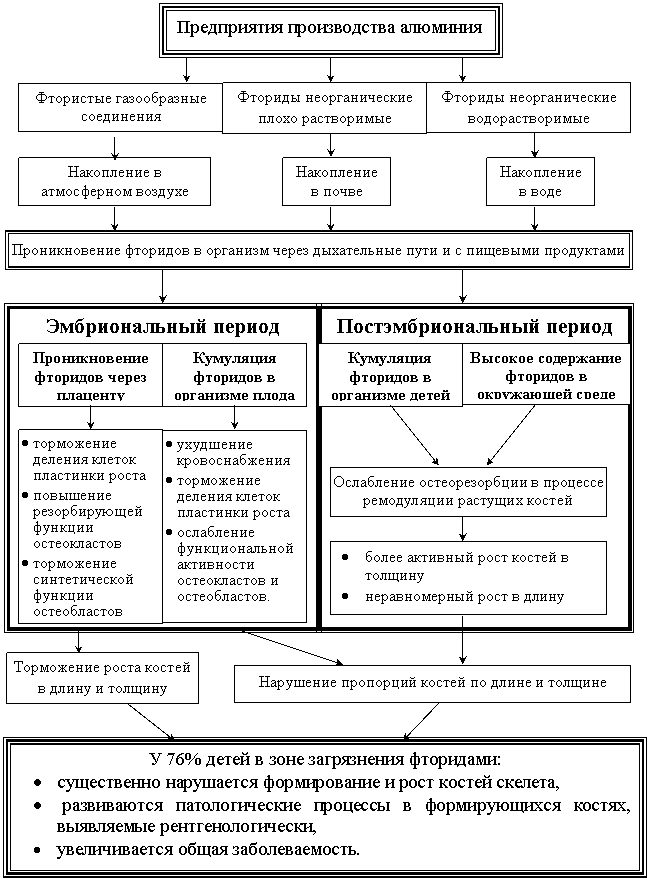
Рис.8. Концептуальная схема токсического действия загрязнения окружающей среды соединениями фтора.
Таким образом:
· В эмбриональный период соединения фтора способны проникать через плаценту, вызывая торможение размножения столбчатого хряща пластинки роста, повышение резорбирующей функции остеокластов и подавление синтетической функции остеобластов, что приводит к торможению роста костей и в длину, и в толщину.
· Соединения фтора способны кумулироваться в организме плода и оказывать токсическое действие в постэмбриональный период, вызывая ухудшение кровоснабжения, торможение деления клеток пластинки роста костей, ослабление функциональной активности остеокластов и остеобластов.
· В постэмбриональный период повышенное содержание фторидов в окружающей среде приводит к ослаблению остеорезорбции в процессе ремодуляции растущих костей, к нарушению пропорций костей кисти и по длине, и по толщине в результате более активного роста костей в толщину и неравномерного, диспропорционального роста в длину.
· В итоге у большинства детей (76%), проживающих на загрязненных фторидами территориях, существенно нарушается формирование и рост костей скелета, развиваются патологические процессы в формирующихся костях, выявляемые рентгенологически, увеличивается общая заболеваемость.
ВЫВОДЫ
1. Приоритетными специфическими загрязняющими веществами на экспонированной территории являются соединения фтора, среднегодовые концентрации которых составляют в атмосферном воздухе 6,2-1,4 ПДК, в почве 5,9-11 ПДК. Уровень общетоксического риска для здоровья жителей составил (IH)=28, риск патологии костно-мышечной системы (IH)=1.9, коэффициент опасности для газообразного фтора составил (QH)=1,1.
2. На территориях размещения предприятий по производству алюминия высокие концентрации соединений фтора оказывают токсическое действие на процессы морфогенеза костей человека в эмбриональный и постэмбриональный периоды онтогенеза, вызывая нарушения формирования и роста костей скелета, что приводит к увеличению общей заболеваемости детского населения.
3. Хроническое влияние соединений фтора на детское население подтверждается величиной коэффициента динамики заболеваемости. Величина коэффициента в экспонированной группе в 2 раза выше по сравнению с контрольной группой. Выявлены значимые различия в структуре заболеваемости детей и подростков изучаемых территорий, в том числе увеличении доли костно-мышечной патологии.
4. Симптомы нарушения морфогенеза костей, выявленные рентгенологически, у детского населения на экспонированной территории встречаются в 3,8 раза чаще (76%), чем на контрольной территории (20%). В условиях повышенных концентраций соединений фтора в окружающей среде рост костей кисти у детей происходит неравномерно и диспропорционально вследствие поражения зон роста и дисбаланса процессов остеогенеза и остеорезорбции в периоды активации роста – у девочек в периоды детства, у мальчиков – в подростковый период.
5. В условиях загрязнения внешней среды фторидами, у плодов сроком 29 недель в бедренной кости толщина столбчатого хряща пластинки роста в 3 а толщина стенки диафиза в 9 раз меньше, чем в контрольной группе, что обусловлено постепенно нарастающими на протяжении эмбриогенеза ухудшением кровоснабжения костей, активацией остеорезорбции, ослаблением остеогенеза и подавлением размножения клеток пластинки роста,.
6. В эксперименте подтверждено, что соединения фтора проникают через плаценту, накапливаются в организме плода и оказывают токсическое действие в постэмбриональный период, вызывая ухудшение кровоснабжения, торможение деления клеток пластинки роста костей, ослабление функциональной активности остеокластов и остеобластов, результатом чего является замедление роста костей и в длину, и в толщину.
ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ
Учитывая высокий уровень заболеваемости детского и подросткового населения на экспонированной территории в условиях воздействия фтористых соединений необходимо внедрить систему биомониторинга - определение фтора в моче 1 раз в месяц. Детям, (при концентрации фтора в моче выше 1,0 мг/л), проводить оздоровительные мероприятия, включая: детоксикационную терапию с применением энтеросорбентов, диетотерапию, витаминотерапию, иммунопротекторы.
Детям с повышенной концентрацией фтора в моче и предъявляющим жалобы со стороны опорно-двигательного аппарата, на боли в костях и суставах необходим ежегодный осмотр врачом ортопедом. С целью раннего выявления костной патологии диспластического и дистрофического генеза проводить 1раз в год рентгенологическое обследование костей кисти как наиболее традиционного объекта оценки возрастных изменений скелета.
Ключевым фактором морфогенеза костей является фактор - механической нагрузки. В случае выявления признаков костной патологии диспластического и дистрофического генеза костей кисти, необходимо назначать детям ЛФК с комплексом упражнений для укрепления опорно-двигательного аппарата с учетом возрастных особенной.
СПИСОК ОПУБЛИКОВАННЫХ РАБОТ ПО ТЕМЕ ДИССЕРТАЦИИ
1. Шалина строения костей верхней конечности у детей Иркутского региона с учетом экологических факторов внешней среды / , // Морфология. – 1993. – Т.105. – №9-10. С.173.
2. Шалина нарушений в развитии костной ткани у детей, обусловленные техногенным воздействием / , // В Сб. научных трудов посвященных 100 - летию Городской детской больницы. Актуальные вопросы педиатрии и детской хирургии. Иркутск, 1995. - С.107-108.
3. Шалина развития костной ткани у детей, обусловленные техногенным воздействием / , //Тез. докл. III Конгресса Международной Ассоциации морфологов. Морфология. Санкт-Петербург, 1996. – Т.109. – № 2. С.104.
4. Шалина изменения костной ткани у детей в экологически неблагоприятных районах Иркутской области / , // В сб. научных трудов ИГМУ « Актуальные вопросы современной клинической медицины». Иркутск, 1997. – С. 85-86.
5. Шалина остеогенеза у детей в условиях техногенных воздействий фтористых соединений / , , // Тезисы доклада IV Съезда Российских морфологов с международным участием. Российские морфологические ведомости. Москва, 1999г. – № 1-2. С.163.
6. Шалина эпифизы в коротких трубчатых костях кисти / , , // Тезисы доклада IV Съезда Российских морфологов с международным участием. Российские морфологические ведомости. Москва, 1999. – № 1-2. С.164.
7. Корнев фтористых соединений на формирование костной ткани / , , // Всероссийская научно-практическая конференция хирургов. Пятигорск, 1999. – С.263.
8. Шалина характеристика костей кисти у детей / , // В Сб. ИГМУ «Актуальные вопросы современной клинической медицины в условиях ОМС с международным участием. Иркутск, 2000. – Вып.3. С.324.
9. Шалина показатели у детей в некоторых городах Иркутской области / // Тез. V Конгресса Международной Ассоциации морфологов. Морфология. Санкт-Петербург, 2000. – Том.117. – № 3 .С.134.
10. Шалина турецкого седла у детей / , , // Тез. VI Конгресса Международной Ассоциации морфологов. Морфология. Санкт-Петербург, 2002. – Т. – 121. – №2-3. С.175.
11. Шалина костей и типологические особенности черепа / , , // Тез. VI Конгресса Международной Ассоциации морфологов. Морфология. Санкт-Петербург, 2002. – Т. – 121. – № 2-3. С. 176.
12. Шалина развития костной системы у детей в некоторых регионах Восточной Сибири / , , // Сибирский медицинский журнал. – 2002. – №4. – С.70-72.
13. Шалина рентгенологических изменений костей кисти у детей / // V Общероссийский съезд анатомов, гистологов и эмбриологов. Морфологические ведомости Москва, 2004. – №1-2. – С.119.
14. Шалина показатели воздействия соединений фтора на организм / // V Общероссийский съезд анатомов, гистологов и эмбриологов. Морфологические ведомости Москва, 2004. – №1-2. – С.119.
15. , , . Влияние фторидов на эмбриональное развитие бедренной кости человека / , , // В сб: Актуальные проблемы морфологии. Красноярск, 2006. – С.189–190.
16. Шалина костей кисти у детей г. Шелехова / // Морфология. – 2008. – Т.133. – №4. – С.102.
17. Шалина соединений фтора на морфогенез костей белых крыс / // Морфология. – 2008. – Т.133. – №4. – С.103.
18. Шалина костей кисти у детей в зонах алюминиевого производства / , // Сибирский медицинский журнал. – 2006. – №9. – С.78-81.
19. Шалина соединений фтора на рентгеноанатомические параметры и активность роста костей кисти у детей / , // Сибирский медицинский журнал. – 2008. – №6. – С.48-52.
20. Шалина соединений фтора на морфогенез костей кисти у детей / , // Сибирский медицинский журнал. – 2008. – №8. – С.35-38.
21. Шалина соединений фтора на морфогенез бедренных костей плодов человека / , // Сибирский медицинский журнал. – 2009. – №1. – С.42-46.
22. Шалина общей заболеваемости детей и подростков по классам болезней в промышленных городах / , // Сибирский медицинский журнал. – 2009. – №2. – С.66-68.
23. Шалина роста костей кисти как критерий определения биологического возраста детей в различных экологических условиях проживания / , , // Судебно-медицинская экспертиза. – 2009. – № 4. – С.20-24.
24. Шалина костей кисти у детей промышленных городов / , , // Иркутск: «Оттиск», 2009. – 152 с.
25. Шалина вопросы токсического действия фтора // , // Сибирский медицинский журнал. – 2009. – №5. – С. – С. 5-8.
26. Шалина определения биологического возраста и прогноза морфогенеза костей кисти по рентгенологическим показателям/ , . – Иркутск. – 26 с.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
