
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
увеличения температуры. У мышей и крыс величина п близка к таковой у дрозофил (п = 4—5), а у людей несколько больше (примерно 5 у мужчин и 7.6 у женщин) (Rosenberg et al., 1973).
Результаты наших анализов смертности разных видов животных с помощью степенной MMC приведены в табл. 8, из которой видно, что по мере увеличения ВПЖ величина константы Л снижается, а п несколько растет при достаточно высоких значениях коэффициента корреляции между InR t и /.
В приведенных выше исследованиях на степенной MMC указывалось, что преимущество таких моделей по сравнению с экспоненциальными становится особенно отчетливым при обработке данных смертности только от биологических причин (без учета влияния несчастных случаев). Это позволяет допустить, что степенная MMC более чувствительна к «помехам» случайной природы, которые могут быть частично «устранены» введением в модель дополнительных членов, например учитывающих линейно зависящие от возраста составляющие смертности. Уравнение, описывающее такую линейно-степенную MMC, имеет вид:
(15)
При этом величины п (табл. 9) увеличиваются (особенно у дрозофил) по сравнению с данными табл. 8.
Для оценки особенностей геропротекторного эффекта ряда воздействий полезным может оказаться уравнение мультивариационной теории радиационного поражения, которое имеет вид:
NfN0=I - [1-exp (-kt)]p, (16)
где k — константа, численно равная смертности при единичной интенсивности повреждения (одно повреждение за единицу времени) ; P — летальное число повреждений.
Обработка данных смертности дрозофил, инкубированных при 18, 21, 27 и 31 0C, показала, что P соответственно равно,,и, а константа k — 0.11, 0.09, 0.22 и 0.72 (Atlan et al., 1976).
Рассмотренные MMC традиционно наиболее известны и изучены, но, к сожалению, они не самые лучшие. Ряд авторов считает, что организацию биологических систем можно удовлетворительно описать лишь с помощью не менее трех констант. Подобный анализ смертности мышей также показал,, что три константы — минимальное число для описания возрастной динамики смертности (Atlan et al., 1976). Обращает на себя внимание также то, что указанные выше MMC являются феноменологическими и в большинстве случаев константы соответствующих уравнений не имеют четкой биологической интерпретации. Содержательный анализ этих и ряда других MMC проведен и H. С. Гавриловой (1986).
В последние годы появились самые разные подходы к решению такой важной задачи, как поиск типа распределений смертности
7 В В Фролькис, X К Мурадян 97
и создание MMC. Весьма плодотворными оказались попытки использования отдельных положений теории информации и надежности (Murthy et al., 1981; Smith, White, 1982; Woodbury, Maten, 1983; Witten, 1984). Современные методы моделирования и обработки результатов сделали возможным и необходимым отказ от анализа смертности с помощью 2—3-константных MMC и применение более четко формализованных и многоконстантных моделей. Каждая константа в такой модели должна количественно оценить влияние определенной функциональной системы или определенным образом сгруппированных факторов старения и смертности, что позволит получить более дифференцированную оценку действия тех или иных геропротекторов. В качестве примера наиболее простой MMC можно представить разработанную нами модель, позволяющую провести оценку вклада повреждающих процессов и противоборствующих с ними процессов витаукта. В основу этой MMC положены три допущения.
1. Вероятность смерти в единицу времени (Rt) пропорциональна отношению скорости повреждающих процессов (Vn) и процессов витаукта (V8):
Rt=k- VJV^ (17)
где k — константа размерности, численно равная R1 при Vn= V8.
2. Скорость повреждающих процессов прямо пропорциональна концентрации повреждающих агентов. Подобное допущение не противоречит логике цепного характера распространения повреждений, когда каждое новое повреждение увеличивает вероятность образования других повреждений:
Vn=UcJcLt^k1PnI (18)
где /гп и Cn—соответственно константа скорости повреждающих процессов и концентрация повреждающих агентов.
3. Скорость витаукта также подчиняется аналогичной кинетике (т. е. каждый новый акт восстановления увеличивает вероятность новых восстановлений). Однако в отличие от Cn соответствующая константа процессов витаукта (св) из-за постоянного повреждения самих восстановительных процессов с возрастом снижается обратно пропорционально времени (c'B — ca/t). Тогда аналогично (18)
VB = dcJdt = kicB = kBcJt, (19)
где къ и св—соответственно константа и концентрация агентов, участвующих в процессе витаукта.
Разделив переменные, решив дифференциальные уравнения (18) и (19) и подставив их значения в (17), получим:
Rt = ^exp(knt-ka\nt), (20)
98
где с°п и с\—соответственно концентрации повреждающих и восстанавливающих агентов в момент времени t = 0 и t=\. Поскольку стоящее перед экспонентой выражение состоит только из величин, не зависящих от времени, то для изучаемой популяции его можно считать также константой. Обозначив это выражение ехр (/го) и после соответствующей подстановки прологарифмировав (20), получим окончательное выражение предлагаемой MMC:
\nRt=ko + k4 — kBint, (21)
где k0, kn и kB—положительные и специфические константы, характеризующие вклад в общую смертность не изменяющихся с возрастом факторов (&о), повреждающих факторов (kn) и факторов витаукта (kB). Нетрудно заметить, что последняя группа факторов противодействует, снижает эффект первых двух и в то же время сопряжена с ними, т. е. по мере увеличения вклада повреждающих факторов (kj) при увеличении / растет и значение kB\nt. При этом до определенного момента темпы возрастных изменений процессов витаукта (kB\nt) должны опережать, а на более поздних этапах уступать темпам возрастных изменений повреждающих процессов (kj). Иначе говоря, при t = kjkn (2\) имеет минимум и неограниченно растет по обе стороны от этой точки. Такая модель может описать смертность не только после достижения зрелости, как это делают указанные выше экспоненциальные и степенные MMC, но и на ранних этапах онтогенеза. В то же время бесконечный рост смертности при очень больших и малых значениях t указывает на ограниченность такой линейнологарифмической MMC на «хвостах» распределения.
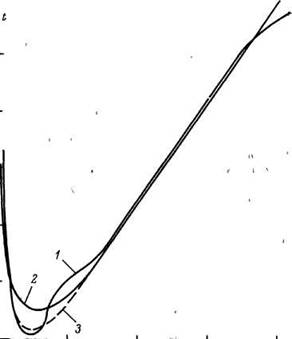
Какими бы ни были логические допущения и аппарат формализации MMC, основной критерий их справедливости — соответствие с экспериментальными данными. Результаты сравнительной оценки предсказанной нашей модели и фактической смертности мужского населения СССР по данным 1959 г. приведены на рис. 18. Из представленных графиков видно, что, за исключением возрастных периодов 5—40 лет и более 90 лет, модель дает величины, близкие к фактической смертности. Поскольку резкий рост смертности в возрасте 15—35 лет, как известно, в основном обусловлен влиянием «небиологических» факторов (несчастные случаи и др.), то мы повторили оценку констант модели, не включив в расчет данные за этот период. Ход модельной кривой в этом случае указывает на значительное улучшение предсказаний модели в области 5—15 лет. Данные аналогичной оценки (без учета смертности в 15—35 лет) смертности мужчин и женщин на основе модельных таблиц ООН приведены на рис. 19, из которого видно, что, за исключением указанного периода, наблюдается удовлетворительное совпадение предсказаний модели и фактических данных.
Величины констант этой MMC для разных видов животных приведены в табл. 10. По мере увеличения ВПЖ наблюдается снижение констант k0 и Zen и увеличение kB. Судя по этим дачным,
т- 99
InR |
 -6
-6
 25
25
50 Годы
75
100
Рис. 18. Возрастная динамика смертности (/) и ход кривой линейно-логарифмической MMC с учетом (2) и без учета (3) данных смертности за период 15—35 лет для мужского населения СССР.
у долгоживущих видов наблюдается снижение повреждающих и повышение восстанавливающих процессов. Но если от дрозофилы к человеку интенсивность процессов витаукта растет в несколько раз, то повреждающих процессов снижается примерно на порядок и более. Эти данные свидетельствуют о том, что эволюционный рост ПЖ сопровождался не столько интенсификацией восстановления образовавшихся повреждений, сколько их профилактикой, предупреждением их образования. Эволюционный рост ВПЖ, очевидно, сопровождался изменением соотношения скоростей повреждающих и противоборствующих с ними процессов витаукта. Интересно с этой точки зрения сопоставление отношения констант k Jkn у животных с разной ВПЖ. Так, у наиболее короткоживущего из исследованных нами видов — дрозофил — эта величина составляла всего 0.03 и увеличивалась на порядок у крыс. У людей kjkn превосходило величину, характерную для крыс и дрозофил, в 34 и 450 раз соответственно, т. е. практически пропорционально ВПЖ.
100
Представляла определенный интерес оценка констант линейнологарифмической MMC на материале смертности мужчин и женщин в разных странах (табл. 11). Из этих данных видно, что единственная константа, по величине которой женщины существенно отличаются от мужчин, — это /гв. По этому показателю, а также по отношению kjkn женщины превосходили мужчин примерно на 10 %, т. е. на столько, на сколько по ПЖ. При этом между константами MMC существовала зависимость вида:
&в=0.188
13.012 kn— 1.295.
(22)
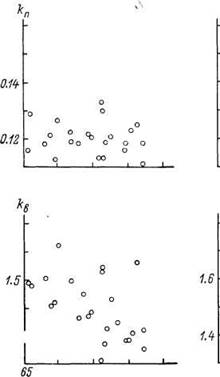
При сопоставлении констант линейно-логарифмической MMC с ожидаемой СПЖ(ППЖ) оказалось, что по мере увеличения ППЖ &ппрактически не изменяется, а kBn ko снижаются (рис. 20).
Таким образом, проведенный краткий обзор MMC указывает на необходимость всестороннего анализа возрастной динамики смертности, как одного из наиболее перспективных путей раскрытия механизмов старения, возможностей продления жизни. Некоторые из имеющихся MMC позволяют количественно охарактеризовать «вклад» отдельных функциональных систем и групп факторов в общую смертность, выявить наиболее уязвимые возрастные звенья. Несколько иные подходы к решению этих задач разработаны при изучении детерминант ИПЖ и БВ, которые будут рассмотрены в следующем разделе.
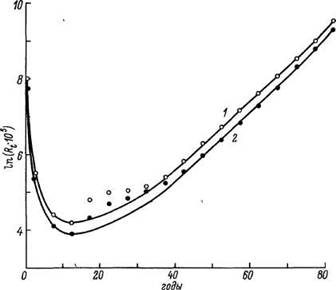
Рис. 19. Возрастная динамика смертности и ход кривой линейно-логарифмической MMC для мужчин (/, светлые кружки) и женщин (2, темные) на основании модельных таблиц смертности ООН.
101
~д
О О
ОСР О
-6
QO0O0O
I О 0„
|
1.3 - |
70 |
75 10 |
о о
о
OO O0O
 CP
CP
75
80 годы
Рис. 20. Зависимость констант линейно-логарифмической MMC от СПЖ мужчин (А) и женщин (Б) в ряде стран мира.
Таблица 10 Видовые отличия констант линейно-логарифмической MMC
Вид | K0 | Kn | кв | KjKn |
Дрозофила | —0.757 | 16.775 | 0 497 | 0 030 |
Крыса | —3.812 | 1 359 | 0.524 | 0.386 |
Морская свинка | —4 469 | 0 496 | 1 605 | 3.236 |
Собака | —6 747 | 0.762 | 1 906 | 2.501 |
Человек | —4.730 | 0.113 | 1.505 | 13319 |
Таблица 11 Константы линейно-логарифмической MMC для разных стран
Мужчины | Женщины | ||||||
Страна | |||||||
«о | Kn | ка | K0 | Kn | |||
Австралия | 5.739 | 0.119 | 1.327 | 5 466 | 0.124 | 1.506 | |
Австрия | 5.946 | 0.119 | 1.336 | 5716 | 0 127 | .585 | |
Англия, Уэльс | 5.720 | 0.134 | 1.536 | 5 499 | 0.135 | 638 | |
Болгария | 6.353 | 0.122 | 1 495 | 6.106 | 0.128 | .670 | |
Венгрия | 6.208 | 0.129 | 1 480 | 5.919 | 0.138 | .697 | |
Греция | 6.061 | 0.125 | 1 562 | 5 862 | 0.139 | .856 | |
Египет | 7.936 | 0.105 | 1.574 | 7.970 | 0 106 | .718 | |
Исландия | 5.750 | 0.109 | 1.257 | 4 805 | 0.133 | .511 | |
Испания | 6.030 | 0.120 | 1.438 | 5.829 | 0 124 | .620 | |
Италия | 5.968 | 0.130 | 1.541 | 5.725 | 0.134 | .723 | |
Канада | 5.862 | 0.113 | 1.269 | 5.592 | 0 121 | 1 491 | |
Новая Зеландия | 5.936 | 0 121 | 1 381 | 5 637 | 0.123 | 1 492 | |
Норвегия | 5.523 | 0 118 | 1 288 | 5 070 | 0 128 | .513 | |
Польша | 6.246 | 0.121 | 1.409 | 5.939 | 0.131 | 680 | |
Португалия | 6.798 | 0.116 | 1.485 | 6.597 | 0 123 | 1.715 | |
Румыния | 6.714 | 0 118 | 1 506 | 6 491 | 0.125 | 678 | |
Финляндия | 5.457 | 0.119 | 1 192 | 5 131 | 0.129 | 1.489 | |
Франция | 5.661 | 0.113 | 1.110 | 5 369 | 0 114 | 1.357 | |
ФРГ | 5 837 | 0.112 | 1.369 | 5 607 | 0.126 | 1 556 | |
Чили | 6.812 | 0.113 | 1.419 | 6 601 | 0.123 | 665 | |
Югославия | 6.630 | 0.127 | 1.623 | 6 539 | 0.134 | 1.853 | |
Япония | 5.411 | 0 119 | 1.339 | 5 262 | 0 123 | 1.493 | |
Модельные табл. ООН | 6.780 | 0.113 | 1.505 | 6 555 | 0.112 | .512 |
Примечание Исходные данные смертности (105- год ) взяты из Ежегодник мировой статистики Демографическая статистика и причины смерти Женева, 1984

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 |
