

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Очевидно, что популяций, удовлетворяющих этим условиям в течение длительного времени, в природе не существует. На популяции всегда действуют внешние и внутренние факторы, нарушающие генетическое равновесие. Длительное и направленное изменение генетического состава популяции, ее генофонда получило название элементарного эволюционного явления. Без изменения генофонда популяции невозможен эволюционный процесс.
5.3. Динамические показатели популяции
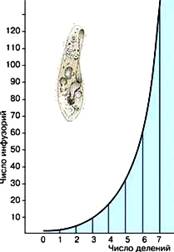 Любая популяция теоретически способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды. Даже самый медленно размножающийся вид способен в короткое время произвести столько особей, что для них не хватит места на земном шаре. Всего за пять поколений, т. е. за 1–1,5 летних месяца, одна единственная тля может оставить более 300 млн. потомков. Если бы все зародыши сохранялись, а все потомство выживало, численность любой популяции через определенные интервалы увеличивалась бы в геометрической прогрессии, и это несмотря на то, что одни производят за всю жизнь всего несколько яиц или детенышей, а другие – тысячи и даже миллионы зародышей. Рост в геометрической прогрессии выражается на графике экспоненциальной кривой. назвал этот процесс «давлением жизни».
Любая популяция теоретически способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды. Даже самый медленно размножающийся вид способен в короткое время произвести столько особей, что для них не хватит места на земном шаре. Всего за пять поколений, т. е. за 1–1,5 летних месяца, одна единственная тля может оставить более 300 млн. потомков. Если бы все зародыши сохранялись, а все потомство выживало, численность любой популяции через определенные интервалы увеличивалась бы в геометрической прогрессии, и это несмотря на то, что одни производят за всю жизнь всего несколько яиц или детенышей, а другие – тысячи и даже миллионы зародышей. Рост в геометрической прогрессии выражается на графике экспоненциальной кривой. назвал этот процесс «давлением жизни».
Фактически во всех живых организмах заложена способность размножаться беспредельно. Эта способность делает жизнь очень мощной силой на Земле. Огромная живая масса организмов поддерживает круговорот веществ на планете, создает горные породы, почвы, регулирует состав вод и атмосферы. Но в природе биотический потенциал популяции никогда не реализуется полностью. Главный ограничитель на пути к беспредельному размножению организмов – нехватка ресурсов, самых необходимых: для растений – минеральных солей, углекислого газа, воды, света; для животных – пищи, воды; для микроорганизмов – разнообразных потребляемых ими соединений. Запасы этих ресурсов не бесконечны, в разных частях планеты они имеют свои пределы, и этим сдерживается размножение видов. Второй ограничитель – влияние неблагоприятных условий, замедляющих рост и размножение организмов, даже если есть необходимые для этого ресурсы. Наконец, гибель зародышей или подрастающих молодых особей от врагов и болезней. Так, тысячи желудей, которые ежегодно производит один большой дуб, оказываются съеденными белками, кабанами, сойками, мышами, насекомыми, или поражаются плесневыми грибками и бактериями, или гибнут на стадии проростков по разным причинам. В результате лишь из считанных желудей вырастают взрослые деревья.
Общие изменения численности популяции складываются за счет четырех явлений: рождаемости, смертности, вселения и выселения особей (иммиграция и эмиграция).
Рождаемость (скорость рождаемости) – число новых особей, появившихся в популяции за единицу времени в результате размножения.
Различают максимальную и фактическую рождаемость. Максимальная рождаемость – максимальная реализация возможности рождения при отсутствии лимитирующих факторов среды. Фактическая рождаемость – реальная реализация возможности рождения.
Различают абсолютную и удельную рождаемость. Абсолютная (общая) рождаемость, или скорость рождаемости – число особей (яиц, семян и т. п.), родившихся (отложенных, продуцированных и т. д.) за некоторый промежуток времени.
Удельная рождаемость – отношение скорости рождаемости к исходной численности. Эта величина зависит от интенсивности размножения особей: для бактерий – час, для фитопланктона – сутки, для насекомых – неделя или месяц, для крупных млекопитающих – год.
Величина рождаемости зависит от многих причин. Большое значение имеет доля особей, способных в данный период к размножению, что определяется соотношением полов и возрастных групп. Важна также частота последовательности генераций. Так, среди насекомых различают моновольтинные и поливольтинные виды. Первые дают одну, вторые – несколько генераций за год. Например, у тлей за сезон насчитывается до 15 партеногенетических поколений. По числу периодов размножения в течение жизни различают моноциклические и полициклические виды. Моноцикличность, или однократное размножение, свойственна обычно видам с короткой продолжительностью жизни в половозрелом состоянии (лососевые рыбы, поденки, майские жуки и многие другие насекомые). Полицикличность характеризуется повторным размножением особей и присуща большинству позвоночных животных и ряду беспозвоночных, например ракообразным.
У растений выделяют монокарпические и поликарпические виды, т. е. с однократным и многократным размножением в течение жизни.
Размножаемость популяции, как правило, не бывает прямо пропорциональна плодовитости. Плодовитость сильно зависит от степени развития заботы о потомстве или обеспеченности яиц питательными материалами. Среди рыб наибольшее количество икринок выметывают виды с пелагической икрой – сельди, тресковые, камбаловые и др. Например, сахалинская сельдь продуцирует 38–46 тыс. мелких, в доли миллиметра, икринок. У лососевых, зарывающих икру в грунт, развивается меньшее число яиц, но более крупных размеров. Средняя плодовитость амурской горбуши 1300–1500 икринок диаметром 4–6 мм. Наиболее крупная икра у акул и химер, до 6–8 см. Яйца этих рыб имеют к тому же плотную защитную оболочку. Количество их очень невелико – несколько штук на одну самку.
У видов, оберегающих и выкармливающих свой молодняк, плодовитость резко понижена. Размер кладки у птиц разных видов варьирует уже не в тысячи раз, как у рыб, а в пределах от одного яйца (у некоторых хищников, пингвинов, чистиков и др.) до 20–25 (максимальное количество яиц у куриных, например, у серой куропатки).
Большая плодовитость вырабатывается у видов также в условиях более высокой смертности, особенно при сильном давлении со стороны хищников. Отбор на плодовитость компенсирует высокую норму гибели в популяциях. Поэтому при высокой плодовитости рост численности популяции может быть весьма низким. В различных популяциях одного вида плодовитость обычно тем выше, чем более неблагоприятны занимаемые ими места обитания. Так, у многих млекопитающих – зайцев, мышей, полевок – число детенышей в помете на границах ареала больше, чем в его центре.
Гипотеза дифференцированной специализации полов () – дифференциация полов происходит по двум основным направлениям эволюции: изменению (мужской пол) и сохранению (женский пол). Чем больше в популяции женских особей, тем лучше сохраняется ее генотипическая структура; чем больше мужских особей, тем больше скорость или величина изменения этой структуры.
Смертность (скорость смертности) – число особей, погибших в популяции за единицу времени (от хищников, болезней, старости и других причин). Смертность – величина обратная рождаемости.
Различают минимальную и фактическую смертность. Минимальная смертность – минимально возможная величина смертности. Фактическая смертность – реальная величина смертности.
Различают абсолютную и удельную смертность. Абсолютная (общая) смертность, или скорость смертности – число особей, погибших за промежуток времени.
Удельная смертность (а) – отношение скорости смертности к исходной численности.
Скорость роста популяции – изменение численности популяции за единицу времени. Скорость роста популяции может быть положительной, нулевой и отрицательной. Она зависит от показателей рождаемости, смертности и миграции (вселения – иммиграции и выселения – эмиграции). Увеличение (прибыль) численности происходит в результате рождаемости и иммиграции особей, а уменьшение (убыль) численности – в результате смертности и эмиграции особей.
Различают абсолютную и удельную скорость роста популяции. Абсолютная (общая) скорость роста – изменение численности популяции за промежуток времени.
Удельная скорость роста – отношение скорости роста к исходной численности.
При отсутствии лимитирующих факторов среды удельная скорость роста равна величине, которая характеризует свойства самой популяции и называется удельной (врожденной) скоростью роста популяции или биотическим потенциалом вида.
Величина биотического потенциала очень различается у разных видов. Например, самка косули способна произвести за жизнь 10–15 козлят, трихина отложить 1,8 тыс. личинок, самка медоносной пчелы – 50 тыс. яиц, рыба-луна – до 3 млрд. икринок.
Однако в природе, в связи с действием лимитирующих факторов, биотический потенциал популяции никогда не реализуется полностью. Его величина обычно складывается как разность между рождаемостью и смертностью в популяции:
r =b - d,
где b – число родившихся, d – число погибших особей в популяции за один и тот же период времени.
Когда b = d, r = 0 и популяция находится в стационарном состоянии. Когда b > d, r > 0, численность популяции увеличивается. Когда b < а, r < 0, численность популяции сокращается. Формула d = b – r позволяет определить смертность, которую трудно измерить непосредственно, а определить r достаточно просто непосредственными наблюдениями.
Скорость роста может быть выражена в виде кривой роста популяции. Существуют две основные модели роста популяции: J-образная и S-образная.
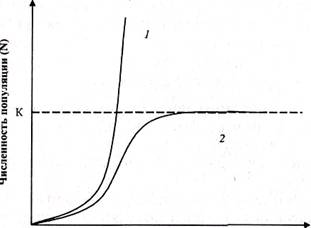 J-образная кривая (1) отражает неограниченный экспоненциальный рост численности популяции, не зависящий от плотности популяции. Такой тип роста возможен пока биотический потенциал популяции (r) реализуется полностью. Это продолжается, пока низка конкуренция за ресурсы. Однако после превышения емкости среды (предельной плотности насыщения, предельной численности) (К), произойдет резкое снижение численности.
J-образная кривая (1) отражает неограниченный экспоненциальный рост численности популяции, не зависящий от плотности популяции. Такой тип роста возможен пока биотический потенциал популяции (r) реализуется полностью. Это продолжается, пока низка конкуренция за ресурсы. Однако после превышения емкости среды (предельной плотности насыщения, предельной численности) (К), произойдет резкое снижение численности.
S-образная (сигмоидная, логистическая) кривая (2) отражает логистический тип роста, зависящего от плотности популяции, при котором скорость роста популяции снижается по мере роста численности (плотности). Скорость роста снижается вплоть до нуля при достижении предельной численности.
5.4. Экологические стратегии выживания популяций
Продолжительность жизни – длительность существования особи. Она зависит от генотипических и фенотипических факторов. Различают физиологическую, максимальную и среднюю продолжительность жизни. Физиологическая продолжительность жизни (ФПЖ) – это продолжительность жизни, которая могла бы быть у особи данного вида, если бы в период всей жизни на нее не оказывали влияние лимитирующие факторы. Она зависит только от физиологических (генетических) возможностей организма и возможна только теоретически. Максимальная продолжительность жизни (МПЖ) – это продолжительность жизни, до которой может дожить лишь малая доля особей в реальных условиях среды. Она варьирует в широких пределах: от нескольких минут у бактерий до нескольких тысячелетий у древесных растений (секвойя). Обычно, чем крупнее растение или животное, тем больше их продолжительность жизни, хотя бывают и исключения (летучие мыши доживают до 30 лет, это дольше, например, жизни медведя). Средняя продолжительность жизни (СПЖ) – это среднее арифметическое продолжительности жизни всех особей популяции. Она значительно колеблется в зависимости от внешних условий, поэтому для сравнения продолжительности жизни разных видов чаще используют генетически детерминированную МПЖ.
Выживаемость – абсолютное число особей (или процент от исходного числа особей), сохранившихся в популяции за определенный промежуток времени.
Z = п / N 100%,
где Z– выживаемость, %; п – число выживших; N – исходная численность популяции.
Выживаемость зависит от ряда причин: возрастного и полового состава популяции, действия тех или иных факторов среды и др. Выживаемость можно выразить в виде таблиц и кривых выживания. Таблицы выживания (демографические таблицы) и кривые выживания отражают, как по мере старения снижается численность особей одного возраста в популяции. Кривые выживания строятся по данным таблиц выживания.
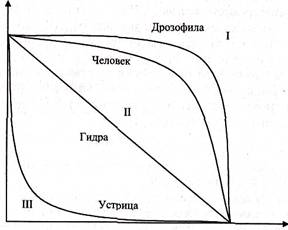 Различают три основных типа кривых выживания. Кривая I типа свойственна организмам, смертность которых на протяжении всей жизни мала, но резко возрастает в ее конце (например, насекомые, погибающие после кладки яиц, люди в развитых странах, некоторые крупные млекопитающие). Кривая II типа характерна для видов, у которых смертность остается примерно постоянной в течение всей жизни (например, птицы, пресмыкающиеся). Кривая III типа отражает массовую гибель особей в начальный период жизни (например, многие рыбы, беспозвоночные, растения и другие организмы, не заботящиеся о потомстве и выживающие за счет огромного количества икринок, личинок, семян и т. п.). Встречаются кривые, сочетающие черты основных типов (например, у людей, живущих в отсталых странах, и некоторых крупных млекопитающих, кривая I вначале имеет резкое падение в связи с большой смертностью сразу после рождения).
Различают три основных типа кривых выживания. Кривая I типа свойственна организмам, смертность которых на протяжении всей жизни мала, но резко возрастает в ее конце (например, насекомые, погибающие после кладки яиц, люди в развитых странах, некоторые крупные млекопитающие). Кривая II типа характерна для видов, у которых смертность остается примерно постоянной в течение всей жизни (например, птицы, пресмыкающиеся). Кривая III типа отражает массовую гибель особей в начальный период жизни (например, многие рыбы, беспозвоночные, растения и другие организмы, не заботящиеся о потомстве и выживающие за счет огромного количества икринок, личинок, семян и т. п.). Встречаются кривые, сочетающие черты основных типов (например, у людей, живущих в отсталых странах, и некоторых крупных млекопитающих, кривая I вначале имеет резкое падение в связи с большой смертностью сразу после рождения).
Комплекс свойств популяции, направленных на повышение вероятности выживания и оставление потомства, называется экологической стратегией выживания. Это общая характеристика роста и размножения. Сюда входят темпы роста особей, время достижения половозрелости, плодовитость, периодичность размножения и т. д.
Так, (1938) различал основные типы стратегий выживания среди растений: виоленты, патиенты и эксплеренты. Виоленты (силовики) – подавляют всех конкурентов, например, деревья, образующие коренные леса. Патиенты – виды, способные выжить в неблагоприятных условиях («тенелюбивые», «солелюбивые» и т. п.). Эксплеренты (наполняющие) – виды, способные быстро появляться там, где нарушены коренные сообщества, – на вырубках и гарях, на отмелях и т. д.
Более подробные классификации выделяют и другие, промежуточные типы. В частности, можно различать еще группу пионерных видов, которые быстро занимают вновь возникающие территории, на которых еще не было никакой растительности. Пионерные виды частично обладают свойствами эксплерентов – низкой конкурентной способностью, но, как и патиенты, обладают высокой выносливостью к физическим условиям среды.
Экологические стратегии популяций отличаются большим разнообразием. Но при этом все их многообразие заключено между двумя типами эволюционного отбора, которые обозначаются константами логистического уравнения: r-стратегия и К-стратегия.
r-стратеги (r-виды, r-популяции) – популяции из быстро размножающихся, но менее конкурентоспособных особей. Имеют J-образную кривую роста численности, не зависящую от плотности популяции. Такие популяции быстро расселяются, но они малоустойчивы, к ним относятся бактерии, тли, однолетние растения и др.
К-стратеги (К-виды, К-популяции) – популяции из медленно размножающихся, но более конкурентоспособных особей. Имеют S-образную кривую роста численности, зависящую от плотности популяции. Такие популяции населяют стабильные местообитания. К ним относятся человек, деревья и др.
Следует отметить, что одну и ту же среду обитания разные популяции могут использовать по-разному, поэтому в одном и том же местообитании могут сосуществовать виды с r- и К-стратегиями. Между этими крайними стратегиями существуют переходы. Ни один из видов не подвержен только r- или только К-отбору.
5.5. Регуляция численности популяции
Любой популяции организмов в конкретных условиях свойствен определенный средний уровень численности, вокруг которого происходят колебания. Отклонения от этого среднего уровня имеют разный размах, но в норме после каждого отклонения численность популяции начинает изменяться с обратным знаком. Выделяют две принципиально разные стороны популяционной динамики: модификацию и регуляцию. Модификация – это случайное отклонение численности, возникающее в результате воздействия самых разнообразных факторов, не связанных с плотностью популяции. Регуляция – это возврат популяции после отклонения к исходному состоянию, совершающийся под влиянием факторов, сила действия которых определяется плотностью популяции.
Модифицирующие факторы, вызывая изменение численности популяций, сами не испытывают влияния этих изменений. Действие их, таким образом, одностороннее. К ним относятся все абиотические влияния среды на организмы, на качество и количество их корма и т. п. Благоприятная погодная обстановка может послужить причиной массовой вспышки размножения вида и перенаселения занимаемой им территории, как, например, в случае стадных саранчовых. Отрицательное воздействие модифицирующих факторов, наоборот, снижает численность популяции иногда до полного ее исчезновения. Влияние модифицирующих факторов, не зависящих от плотности популяции, может вызывать резкий спад ее численности. В сельском хозяйстве чрезвычайно важно прогнозирование погодных условий для возможностей защиты урожая.
Регулирующие факторы не просто изменяют численность популяции, а сглаживают ее колебания, приводя после очередного отклонения от оптимума к прежнему уровню. Это происходит потому, что эффект их воздействия тем сильнее, чем выше плотность популяции. В качестве регулирующих сил выступают межвидовые и внутривидовые отношения организмов. Наиболее эффективные из них – трофические отношения организмов: хищничество, паразитизм, собирательство, пастьба и другие, как прямые, так и косвенные. Прямые связи хищник – жертва наиболее изученные регуляторные механизмы в сообществах. Действие их, таким образом, двустороннее.
Общее число жертв, которых может уничтожить один хищник, сначала растет почти пропорционально росту численности потребляемого вида. Это так называемая функциональная реакция хищников на жертвы. Однако она имеет предел, обусловленный физическими возможностями потребителя. После полного насыщения хищники обычно слабо реагируют на жертву. Новые возможности влиять на популяции жертв возникают при росте численности хищников за счет успешного размножения на хорошей кормовой базе. Это явление получило название численной или количественной реакции хищников на рост численности жертв. Количественная реакция хищников всегда запаздывает по сравнению с ходом размножения жертвы, так как является ответом на изменение кормовой базы.
В лабораторных условиях при совместном содержании хищника и его жертвы отмечаются периодические изменения численности обоих видов, имеющие довольно закономерный характер. В опытах, проведенных нашим отечественным биологом с культурами простейших, было обнаружено следующее. Усиленное размножение хищной инфузории Didinium nasutum закономерно следовало всякий раз за размножением ее жертвы – туфельки Paramecium caudatum. Когда хищник достигал высокой численности, он полностью уничтожал парамеций, после чего наступала гибель самих Didinium, лишенных пищевой базы. В природных условиях такие закономерные циклические изменения численности двух видов, связанных пищевыми отношениями, выявить трудно, так как на их размножение и гибель влияют взаимодействия со множеством других видов, а также абиотические изменения среды. Однако в ряде случаев и в природной обстановке удается заметить регулярные периодические изменения обилия хищников и их жертв.
Можно выделить три основных типа популяционной динамики численности:
I. Стабильный тип – отличается небольшим размахом колебаний (в несколько раз, однако не на несколько порядков величин). Свойствен видам с хорошо выраженными механизмами популяционного гомеостаза, высокой выживаемостью, низкой плодовитостью, большой продолжительностью жизни, сложной возрастной структурой, развитой заботой о потомстве. Целый комплекс эффективно работающих регуляторных механизмов держит такие популяции в определенных пределах плотности. Такова, например, динамика численности крупных млекопитающих и птиц, а также ряда беспозвоночных.
II. Флюктуирующий тип – колебания происходят в значительном интервале плотностей, различающихся на один-два порядка величин. При этом различают три фазы колебательного цикла: нарастания, максимума, разрежения численности. Возврат к стабильному состоянию происходит быстро. Регуляторные механизмы не теряют контроля за численностью популяций, увеличивая свою эффективность вслед за увеличением плотности. Преобладают слабоинерционные меж - и внутривидовые взаимодействия. Такой ход численности широко распространен в разных группах животных.
III. Взрывной тип с вспышками массового размножения – прекращение действия модифицирующих факторов не вызывает быстрого возврата популяции в стабильное состояние. Динамика численности складывается из циклов, в которых различают пять обязательных фаз: нарастания численности, максимума, разреживания, депрессии, восстановления. Для популяций периодически характерны предельно высокий и необычайно низкий уровень численности. Такой ход численности обнаруживается чаще всего у видов с малой продолжительностью жизни, высокой плодовитостью, быстрым оборотом генераций. Он свойствен, например, некоторым насекомым (саранчовые, вредители леса – усачи, короеды, ряд чешуекрылых и пилильщиков и др.), среди млекопитающих отмечен у многих видов мышевидных грызунов.
Тип динамики численности – скорее популяционная, но не видовая характеристика. Популяции одних и тех же видов в различных условиях могут характеризоваться разным ходом динамики численности. Это объясняется преимущественно тем, что среди регуляторных механизмов большую роль играют межвидовые взаимосвязи, которые в пределах ареала вида могут быть разной степени напряженности. Так, многие виды, которые в природных условиях сдерживались врагами, проявляют склонность к вспышкам массового размножения в садах и на полях, где ослаблен биологический контроль.
Ярким примером того, к чему приводит снятие пресса потребителей на популяции жертв, являются масштабные «нашествия» чужеродных видов, попадающих в подходящие для размножения условия на других материках. На родине такие виды испытывают влияние активных регуляторов, занимают соответственные экологические ниши в биоценозах и, как правило, не считаются вредителями в хозяйственной практике человека. В новой среде, не имея специализированных врагов, эти виды за короткие сроки дают взрывы численности, нарушая структуру сложившихся сообществ. С экологических позиций впервые обзор и анализ таких событий был сделан английским зоологом Ч. Элтоном, одним из основоположников популяционной экологии. Вводя понятие «взрыв» применительно к непомерному увеличению численности какого-либо вида, Ч. Элтон объяснял, что он намеренно употребляет это слово, так как оно обозначает «внезапное освобождение сил, проявление которых сдерживалось ранее другими силами». «Нашествия» иноземных видов, занос которых преднамеренно или случайно осуществляется человеком, широко распространены в современном мире и часто вызывают большие осложнения в хозяйственной деятельности. Относительно немногие примеры успешной борьбы с ними связаны, как правило, с интродукцией естественных врагов, т. е. воссозданием системы биоценотической регуляции. Если на «захваченной» видом территории постепенно складывается новый комплекс регуляторов, тогда его численность идет на убыль. Однако для некоторых вселенцев этот процесс происходит достаточно медленно. Таким примером является опасный вредитель картофеля колорадский жук, распространение которого в Европе началось после заноса из Америки в 20-х годах прошлого века с западных берегов Франции. Жук и его личинки имеют ядовитую гемолимфу, спасающую их от насекомоядных птиц и других позвоночных. У колорадского жука на евроазиатской территории до сих пор не сформировался эффективный комплекс потребителей. Потери урожая картофеля от колорадского жука составляют в среднем до 40%.
Одну из простейших математических моделей для системы паразит – хозяин в динамике численности насекомых разработал в 1925 г. статистик А. Лотка. График процесса паразитической инвазии, построенный по его уравнениям, обнаруживает, что в результате взаимодействия двух видов должны возникать осцилляции (колебания) с постоянной амплитудой, которая зависит от соотношения между скоростями увеличения численности двух видов. В это же время математик В. Вольтерра выявил сходные закономерности для системы хищник – жертва, обрабатывая статистические данные рыбного промысла. Один из выведенных им законов – закон периодического цикла – гласит, что процесс уничтожения одного вида другим может привести к периодическим колебаниям численности популяций обоих видов, зависящих только от коэффициентов роста популяций хищника и жертвы и от исходной относительной численности. Лотки и В. Вольтерра позволили выдвинуть идею, что периодический колебательный режим в популяциях может возникнуть в результате межвидовых отношений и без внешнего периодического воздействия. Эта идея оказалась плодотворной для дальнейшего развития теории динамики численности популяций. Однако сама модель являлась не адекватной, т. е. не описывала действительность, так как в природе практически не обнаруживаются подобные непрерывные осцилляции с постоянной амплитудой у пар видов, связанных по типу хищник – жертва или паразит – хозяин. Лотки и В. Вольтерра были чрезвычайно упрощенными, так как исходили из целого ряда нереальных допущений: что изменение численности популяции одного вида немедленно вызывает ответную реакцию популяции другого вида, что «аппетиты» хищника беспредельны, поиски жертв случайны, что плодовитость хищников пропорциональна численности всей популяции жертв.
Гомеостаз популяции – поддержание определенной плотности. Изменение численности зависит от целого ряда факторов среды – абиотических, биотических и антропогенных. Однако всегда можно выделить ключевой фактор, наиболее сильно влияющий на рождаемость, смертность, миграцию особей и т. д.
Популяции многих видов организмов способны к саморегуляции своей численности. Выделяют три механизма торможения роста численности популяций: 1) при возрастании плотности повышается частота контактов между особями, что вызывает у них стрессовое состояние, уменьшающее рождаемость и повышающее смертность; 2) при возрастании плотности усиливается эмиграция в новые местообитания, краевые зоны, где условия менее благоприятны и смертность увеличивается; 3) при возрастании плотности происходят изменения генетического состава популяции, например, быстро размножающиеся особи заменяются медленно размножающимися.
Понимание механизмов регуляции численности популяций чрезвычайно важно для возможности управления этими процессами. Деятельность человека часто сопровождается сокращением численности популяций многих видов. Причины этого в чрезмерном истреблении особей, ухудшении условий жизни вследствие загрязнения окружающей среды, беспокойства животных, особенно в период размножения, сокращение ареала и т. д. В природе нет и не может быть «хороших» и «плохих» видов, все они необходимы для ее нормального развития. В настоящее время остро стоит вопрос сохранения биологического разнообразия. Сокращение генофонда живой природы может привести к трагическим последствиям. Международный союз охраны природы и природных ресурсов (МСОП) издает «Красную книгу», где регистрирует следующие виды: исчезающие, редкие, сокращающиеся, неопределенные и «черный список» безвозвратно исчезнувших видов.
В целях сохранения видов человек использует различные способы регулирования численности популяции: правильное ведение охотничьего хозяйства и промыслов (установление сроков и угодий охоты и отлова рыбы), запрещение охоты на некоторые виды животных, регулирование вырубки леса и др.
В то же время деятельность человека создает условия для появления новых форм организмов или развития старых видов, к сожалению, часто вредных для человека: болезнетворных микроорганизмов, вредителей сельскохозяйственных культур и т. д.
Глава 6. ЭКОЛОГИЯ СООБЩЕСТВ (СИНЭКОЛОГИЯ)
6.1. Понятие о сообществах
Живые организмы находятся между собой и абиотическими условиями среды обитания в определенных отношениях, образуя тем самым, так называемые, экологические системы. Биоценоз – совокупность популяций разных видов, обитающих на определенной территории. Растительный компонент биоценоза называют фитоценозом, животный – зооценозом, микробный – микробоценозом. Ведущим компонентом в биоценозе является фитоценоз. Он определяет, каким будет зооценоз и микробоценоз. Биотоп – определенная территория со свойственными ей абиотическими факторами среды обитания (климат, почва). Экосистема (экологическая система) – система совместно обитающих живых организмов и условий их существования, связанных потоком энергии и круговоротом веществ. Биогеоценоз – это экосистема в границах фитоценоза.
экосистема = биоценоз + биотоп
Термин «биоценоз» был предложен немецким ученым К. Мёбиусом (1877), термин «экосистема» – английским ученым А. Тенсли (1935), а термин «биогеоценоз» – российским ученым (1942). «Экосистема» и «биогеоценоз» – понятия близкие, но не синонимы. Экосистема – понятие более общее. Каждый биогеоценоз – это экосистема, но не каждая экосистема – биогеоценоз. Единая экосистема нашей планеты называется биосферой. Биосфера – экосистема высшего порядка.
По Мёбиусу, возможность видов длительно сосуществовать друг с другом в одном биоценозе представляет результат естественного отбора и сложилась в историческом развитии видов. Дальнейшее изучение закономерностей сложения и развития биоценозов привело к возникновению особого раздела общей экологии – биоценологии.
Масштабы биоценотических группировок организмов очень различны, от сообществ подушек лишайников на стволах деревьев или разлагающегося пня до населения целых ландшафтов: лесов, степей, пустынь и т. п.
Термин «биоценоз» в современной экологической литературе чаще употребляют применительно к населению территориальных участков, которые на суше выделяют по относительно однородной растительности (обычно по границам растительных ассоциаций), например биоценоз ельника-кисличника, биоценоз суходольного луга, сосняка-беломошника, биоценоз ковыльной степи, пшеничного поля и т. д. При этом имеется в виду вся совокупность живых существ – растений, животных, микроорганизмов, приспособленных к совместному обитанию на данной территории. В водной среде различают биоценозы, соответствующие экологическим подразделениям частей водоемов, например биоценозы прибрежных галечных, песчаных или илистых грунтов, абиссальных глубин, пелагические биоценозы крупных круговоротов водных масс и т. п.
По отношению к более мелким сообществам (населению стволов или листвы деревьев, моховых кочек на болотах, нор, муравейников, разлагающихся пней и т. д.) применяют разнообразные термины: «микросообщества», «биоценотические комплексы» и др.
Естественные объединения живых существ имеют собственные законы сложения, функционирования и развития, т. е. представляют собой природные системы.
Важнейшими особенностями систем, относящихся к надорганизменному уровню организации жизни» по классификации немецкого эколога В. Тишлера, являются следующие:
1. Сообщества всегда возникают, складываются из готовых частей (представителей различных видов или целых комплексов видов), имеющихся в окружающей среде.
2. Части сообщества заменяемы. Один вид (или комплекс видов) может занять место другого со сходными экологическими требованиями без ущерба для всей системы.
3. Надорганизменная система существует в основном за счет уравновешивания противоположно направленных сил. Интересы многих видов в биоценозе прямо противоположны.
4. Сообщества основаны на количественной регуляции численности одних видов другими.
5. Размеры надорганизменных систем определяются внешними причинами. Так, биоценоз сосняка-беломошника может занимать небольшой участок среди болот, а может простираться на значительное расстояние на территории с относительно однородными абиотическими условиями.
6.2. Структура биоценоза
Различают видовую, пространственную и экологическую структуры биоценоза.
Видовая структура – число видов, образующих данный биоценоз, и соотношение их численности или массы. То есть видовая структура биоценоза определяется видовым разнообразием и количественным соотношением числа видов или их массы между собой.
Видовое разнообразие – число видов в данном сообществе. Встречаются бедные и богатые видами биоценозы. Видовое разнообразие зависит от возраста сообщества (молодые сообщества беднее, чем зрелые) и от благоприятности основных экологических факторов – температуры, влажности, пищевых ресурсов (биоценозы высоких широт, пустынь и высокогорий бедны видами).
Р. Уиттекер предложил различать следующие типы биоразнообразия: α-разнообразие – видовое разнообразие в данном местообитании; β‑разнообразие – сумма всех видов всех местообитании в данном районе; γ-разнообразие – разнообразие ландшафтов (объединение α- и β-разнообразия).
Законы разнообразия Жаккара – 1) видовое разнообразие территории (γ-разнообразие) прямо пропорционально разнообразию ее экологических условий; 2) видовое богатство сообщества (α-разнообразие) растет одновременно с расширением площади и уменьшается по мере увеличения однородности последней.
Правило географической обусловленности изменений разнообразия Де Кандоля-Уоллеса – по мере продвижения с севера на юг, как правило, наблюдается увеличение видового разнообразия сообществ.
Правило Дарлингтона – уменьшение площади острова в 10 раз, как правило, сокращает число живущих на нем животных вдвое.
Различает бедные и богатые видами биоценозы. В полярно арктические пустынях и северных тундрах при крайнем дефиците тепла, в безводных жарких пустынях, в водоемах, сильно загрязненных сточными водами, – везде, где одни или сразу несколько факторов среды далеко уклоняются от среднего оптимального для жизни уровня, сообщества сильно обеднены. Невелик видовой спектр и в тех биоценозах, которые часто подвергаются каким-либо катастрофическим воздействиям, например ежегодному затоплению при разливах рек или регулярному уничтожению растительного покрова при пахоте, применении гербицидов и других антропогенных вмешательствах. И наоборот, везде, где условия абиотической среды приближаются к оптимальным в среднем для жизни, возникают чрезвычайно богатые видами сообщества. Примерами их могут служить тропические леса, коралловые рифы с их многообразным населением, долины рек в аридных районах и т. д.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 |
