

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК, 2009, том 79, № 1, с. 57-72
|
Традиционно основой создания новой техники считались точные науки - физика, математика и химия. Они предлагали решения проблем, связанных с достижением предельных характеристик - максимальной дальности, скорости, прочности, точности и т. п., что особенно важно в военной сфере. Однако в последние годы при разработке новой техники инженеры всё чаще обращаются к биологическим наукам. В публикуемой ниже статье речь идёт о применении бионического подхода к созданию технических средств ближней подводной локации, ориентации и связи. Такие разработки более 30 лет проводятся в Институте проблем экологии и эволюции им. РАН.
ЭЛЕКТРИЧЕСКИЕ РЫБЫ - БИОЛОГИЧЕСКИЙ ПРОТОТИП
НОВОЙ ТЕХНИКИ
, ,
Бионическое моделирование требует прежде всего создания взаимосогласованной системы научных представлений и тесного взаимодействия широкого круга специалистов из разных научных областей, в нашем случае - электроэкологии, нейрофизиологии, электродинамики проводящих сред, электроники, океанологии, электрохимии. Конечная цель бионики - техника, и поэтому в задачи бионического моделирования входит адаптация результатов биологических исследований для инженеров. Но прежде чем предлагать инженерам модели, их нужно подвергнуть физико-математическому анализу. Необходимая для этого система физических представлений должна быть тоже проанализирована применительно к классу решаемых задач с учетом имеющихся биологических представлений. В отличие от радиосвязи, электрокоммуникация рыб - неволновая, и многие привычные подходы, распространённые и эффективные в случае волновой связи, здесь следует применять с большой осторожностью, а от
некоторых отказаться. Это же касается и инженерных рекомендаций, например по оптимизации антенн.
Дальность электрокоммуникации у рыб составляет единицы-десятки сантиметров. Привычные дальности технических средств связи - километры. Только детальный физико-математический анализ позволяет предсказать, во сколько раз и при каких условиях можно увеличить масштаб модели так, чтобы она ещё не утратила работоспособность, но уже приобрела практическую значимость.
Одна из самых больших проблем, которую приходится решать в процессе бионического моделирования, - преодоление языковых барьеров, поскольку биологи, физики и инженеры используют разные термины и опираются на разные базовые понятия. Более того, одни и те же термины, например "волна" или "фаза", в разных научных дисциплинах и в инженерной практике могут

57
58
ОЛЬШАНСКИЙ и др.
употребляться с разной степенью строгости и обобщения, что приводит к недоразумениям, порой весьма серьёзным. Бионика берёт за основу биологические модели, поэтому она не ограничивается чисто физическим анализом, а пытается вписать рассматриваемую модель в реальный мир, в его экологию и эволюцию.
ФУНКЦИОНАЛЬНАЯ МОДЕЛЬ СЛАБОЭЛЕКТРИЧЕСКОЙ РЫБЫ
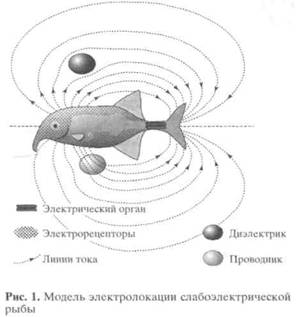
Электрическими называют рыб, имеющих специальные органы для генерации электрических разрядов. У некоторых из них электрические органы занимают большую часть тела и способны генерировать очень мощные разряды, позволяющие добывать пищу и обороняться от врагов. Такие рыбы называются сильноэлектрическими. Существуют и слабоэлектрические рыбы, например клюворылообразные (Mormyri-formes) или гимнотообразные (Gymnotiformes), которые также имеют чётко оформленные электрические органы, но их слабые разряды недостаточны для парализации жертв. Вопрос о функциональном назначении электрических органов таких рыб был поставлен в середине XIX столетия Ч. Дарвиным как серьёзная проблема эволюционной теории, однако до середины XX в., до работ Г. Лиссманна [1,2], этот вопрос оставался без каких-либо вариантов ответа. Лиссманн предположил, что электрические органы являются частью более общей системы, включающей специальные электрорецепторы для восприятия слабых электрических токов. Функциональная модель такой общей системы, предложенная Лиссман-ном, выглядит следующим образом.
Рыба создаёт вокруг себя электрическое поле, напоминающее поле диполя (рис. 1). Её кожу, имеющую высокое сопротивление, пронизывают тысячи каналов с низким сопротивлением, через которые проходят линии тока. В конце каналов находятся сенсорные клетки электрорецепторов, посылающие информацию в мозг. Внешние объекты искажают картину поля, и рыба чувствует искажения, что позволяет ей обнаруживать и различать объекты, то есть осуществлять локационную функцию. Рыба может ориентироваться в пространстве как по рельефу электропроводности (активная электроориентация), так и по особенностям картины фонового электрического поля (пассивная электроориентация). Кроме того, рыба чувствует электрические разряды другой рыбы и может ответить на них изменением ритма своих разрядов, осуществляя таким образом электрокоммуникацию.
В добавление к очень убедительной биологической аргументации Г. Лиссманн совместно с К. Мэйчиным провёл эксперименты по условно рефлекторной методике [3]. На их основе был выполнен подробный физический анализ модели, получены количественные оценки электрочувствительности рыб и определены возможности электролокации такого типа, предложены алгоритмы повышения отношения сигнал/шум, благодаря которым можно приблизиться к физическим пределам восприятия слабых электрических токов. Тройное описание слабоэлектрической рыбы - как биологического объекта, физической модели и инженерной модели - даёт хорошую основу для бионического моделирования.
Привлекая физиков и инженеров к совместным разработкам и объясняя им, как устроены рыбы, биологи должны, помимо разработки общей функциональной модели, ответить на множество конкретных вопросов:
• как устроены электрорецепторы и электрогенераторные ткани;
• каковы пороги чувствительности и формы генерируемых разрядов;
• каковы механизмы обработки электросенсорной информации в мозге и особенности поведения слабоэлектрических рыб;
• в чём сходство и в чём различие электросистем у рыб, принадлежащих к разным видам, и как это связано со степенью родства, с условиями обитания и образом жизни.
Несколько обстоятельств существенно облегчают адаптацию результатов фундаментальных биологических исследований к их использованию в инженерной практике. Во-первых, биоников интересуют в основном функциональные модели и физиологические механизмы электрогенерации, электрорецепции, обработки сенсорной информации и функционирования систем в целом. Язык
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
ЭЛЕКТРИЧЕСКИЕ РЫБЫ - БИОЛОГИЧЕСКИЙ ПРОТОТИП НОВОЙ ТЕХНИКИ 59
физиологии ближе к физико-техническому, чем язык других биологических направлений. Более того, всякий раз, когда есть возможность продемонстрировать или хотя бы предположить наличие морфофункционального соответствия, предложить механизм, поясняющий особенности поведения, или сравнить одинаковые феномены у представителей разных групп электрических рыб, физиологи охотно приводят и комментируют соответствующие морфологические, анатомические, этологические или таксономические сведения, то есть они берут на себя большую часть работы по адаптации биологических знаний к физико-техническому восприятию.
Во-вторых, описание электрической рыбы как бионического прототипа сопряжено с гораздо более мягкими требованиями к достоверности моделей, чем в биологической науке. Биолог обязан тщательно проверить, относятся ли все исходные данные для построения модели к рыбам одной группы или даже одного вида. Модель, построенная биологом, не должна противоречить ни одному известному факту, полученному для данного вида, либо оспаривать достоверность установления этого факта. Биолог должен помнить об эволюционных ограничениях - любой орган, любая функция имели свою эволюционную историю, и модель биолога не должна противоречить ей. Инженера больше интересуют сами идеи, предлагавшиеся по ходу исследований, чем доказательства их достоверности применительно к рыбам. Рыба, которую хочет создать бионик, не имеет видового названия. Биологические идеи - это кубики, из которых бионик будет складывать свои композиции, и он вовсе не обязан включать все эти кубики в конечное решение. В любой момент времени инженер вправе заменить любой узел на другой, взятый из другой отрасли, изготовленный по другой технологии и имеющий другую функцию. Химеры и конвергентная эволюция в технике вполне допустимы. Даже отвергнутые биологами гипотезы могут оказаться технически плодотворными, а достоверность и эффективность их применения будут определяться другими критериями.
Не секрет, что далеко не все специалисты считают бионику плодотворным подходом к созданию новой техники. Многие учёные высоко ценят верность своей специализации и скептически относятся к призывам преодолеть разделение научных дисциплин ради успеха прикладных разработок. Тем не менее совместная деятельность биологов и физиков может быть чрезвычайно эффективной для науки и техники. Свидетельство тому - роль электрических рыб в истории формирование базовых идей и понятий общей теории электричества. Напомним основные этапы этой истории.
В гг. Дж. Уолш и Г. Кавендиш выполнили серию экспериментов с морскими нарко-выми скатами (Narkidae) и пресноводными электрическими угрями (Electrophorus electricus). К тому времени биологи, испытавшие на себе сходство ощущений от воздействия лейденской банки и этих рыб, полагали, что природа разрядов электрическая. Но физики возражали, по их мнению, проводящая среда полностью экранирует электрические источники, а значит, электрических рыб быть не может, как и иных электрических явлений в воде. Чтобы разрешить противоречие между доводами биологов и физиков, Кавендишу фактически пришлось открыть электростатическую аналогию, согласно которой токи в проводящей среде являются источниками электрического поля в той же мере, что и заряды в диэлектрической. Кавендиш изготовил физическую модель электрической рыбы, с помощью которой показал, что в водной среде происходит ослабление токов и напряжений, однако не до полного нуля [4]. По мнению историков науки, эти исследования Уолша и Кавендиша имели существенное значение для становления общей теории электричества и породили электродинамику проводящих сред [5, 6].
Другой пример - спор Л. Гальвани и А. Вольты. Один из главных пунктов разногласий - принципиальны ли различия между электрическими органами электрических рыб и обычными нервно-мышечными комплексами? Гальвани утверждал, что различия не принципиальны. Его эксперименты показали, что электричество свойственно не только электрическим рыбам, но и всему живому. Любая мышца, любой нерв чувствительны к электрическим стимулам и способны генерировать электрические импульсы. Вольта, повторивший опыты Гальвани, стоял на противоположной точке зрения: электрические органы принципиально отличны от обычных мышц и нервов. Созданная им модель электрической рыбы -вольтов столб - стала одним из величайших технических изобретений человечества и фактическим началом электротехники, а опыты Гальвани привели к возникновению электрофизиологии.
Бионический характер своих исследований Вольта не скрывал, напротив, он всячески подчёркивал связь собственного изобретения с его биологическим прототипом: "Этот прибор, более сходный, по существу, с естественным электрическим органом электрического ската или электрического угря и т. п., чем с лейденской банкой и известными электрическими батареями, я назову искусственным электрическим органом" [7, с. 74]. Следует отметить, что физические представления того времени не позволяли объяснить принцип действия вольтова столба и аналогия с биологиче-
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
60
ОЛЬШАНСКИЙ и др.
|
ским прототипом, по сути, была единственным основанием для изобретения Вольты.
В XIX столетии электрическая теория всё более резко отходит от своего медико-биологического прошлого. После того как М. Фарадей, который ещё интересовался животным электричеством и ставил эксперименты с электрическими угрями [8], фактически завершил феноменологию макроэлектродинамики, Дж. Максвелл придал ей стройное математическое описание. Ссылки на биологию стали казаться излишними, но в первых экспериментах по радиосвязи в качестве приёмника ещё выступала лягушачья лапка.
СПЕЦИФИКА ПОДВОДНОЙ СВЯЗИ НА ГАЛЬВАНИЧЕСКИХ ТОКАХ
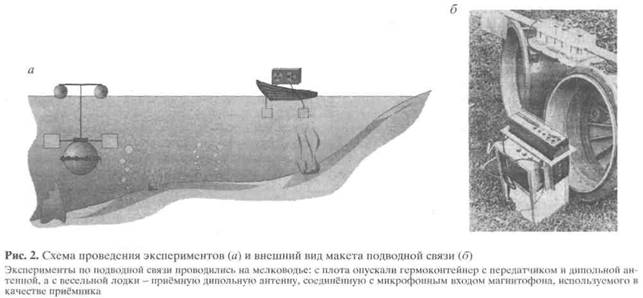
После публикаций Лиссманна появились сообщения о попытках связи на гальванических токах в открытом море. Для приёма и передачи электрических сигналов было достаточно простых ди-польных антенн из двух металлических электродов и усилителей от бытовой полупроводниковой аудиотехники. Сообщалось о достижении дальности связи в сотни метров и даже километров при потребляемых мощностях порядка единиц-долей ватта [9-11]. В конце 1960-х годов экспериментальные исследования возможностей подводной коммуникации на гальванических токах начались в Институте эволюционной морфологии и экологии животных АН СССР (ныне Институт проблем экологии и эволюции РАН - ИПЭЭ РАН). Под руководством был создан действующий макет системы подводной речевой связи (рис. 2), получивший название "Гимнарх" в честь открытий Лиссманна, первоначально сделанных на нильской щуке Gymnarchus niloticus.
Принятые речевые сигналы были различимы при удалении приёмника от передатчика на сотни метров - единицы километров. Следует отметить, что такие дальности возможны только на мелководье до глубин нескольких метров. С ростом глубины места, а также глубины погружения передатчика и приёмника предельная дальность снижается и на произвольно больших глубинах составляет десятки метров. Но даже с этой оговоркой практическая значимость подобных устройств подводной связи была очевидной и нуждалась в теоретическом обосновании, поскольку было широко распространено мнение, что беспроводная электромагнитная связь в морской воде невозможна вообще.
Специфика подводной связи на гальванических токах состоит в первую очередь в том, что эта связь неволновая. Водная (проводящая) среда отличается от воздушной (диэлектрической) тем, что в воде не могут распространяться радиоволны. Это определяется характерным временем экранирования свободных зарядов. Время экранирования для воздуха около 300 с. Соответственно уже на частотах выше единиц герца экранирование зарядов за один период несущественно, то есть эта среда - диэлектрическая. Время экранирования для пресной воды - доли микросекунд, а для морской - доли наносекунд. Если на оптических частотах водная среда может рассматриваться как диэлектрическая, то на частотах звукового диапазона или радиочастотах она является проводящей, то есть успевающей полностью экранировать свободные заряды.
Проводящие и диэлектрические среды имеют разные характерные масштабы для распространяющихся электромагнитных волн. В диэлектрических средах характерный пространственный
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1
2009
ЭЛЕКТРИЧЕСКИЕ РЫБЫ - БИОЛОГИЧЕСКИЙ ПРОТОТИП НОВОЙ ТЕХНИКИ 61
масштаб - это длина волны. Элемент среды, размер которого равен длине волны, в равной степени проявляет как индуктивные, так и ёмкостные свойства, то есть является резонатором на данной частоте. Резистивные свойства диэлектрической среды на расстоянии одной длины волны проявляются слабо. На расстояниях от источника поля, заметно превышающих длину волны, в так называемой волновой зоне, поперечные компоненты напряжённости электрического и магнитного полей убывают обратно пропорционально первой степени расстояния, а не третьей степени, как вблизи источника. Продольная компонента затухает обратно пропорционально квадрату расстояния и в волновой зоне становится пренебрежимо малой по сравнению с поперечными. Малое затухание поперечных компонент электромагнитного поля в волновой зоне определило вектор развития радиотехнической методологии, возникшей для анализа и практического использования поперечных электромагнитных волн.
В проводящей среде её ёмкостные свойства практически не заметны на фоне резистивных. Характерным размером становится толщина скин-слоя - расстояние, на котором индуктивные и резистивные свойства среды проявляются соизмеримым образом. Элемент среды размером в один скин-слой представляет собой однозвенный низкочастотный фильтр. С ростом расстояния число звеньев этого фильтра увеличивается, и он всё сильнее (экспоненциально) гасит верхние частоты. Статическое затухание, обратно пропорциональное кубу расстояния, становится не худшим случаем, как в диэлектриках, а лучшим. Экспоненциальное затухание в проводящих средах на расстояниях, заметно превышающих толщину скин-слоя, фатально. Его нельзя компенсировать увеличением ни мощности, ни размера антенн. Скажем, для частоты 4 кГц в морской воде толщина скин-слоя равна 4 м. Это означает, что на расстоянии 100 м ослабление составит десятки миллиардов раз. Для частоты 400 кГц такое затухание должно иметь место на расстоянии в 10 м.
Из-за невозможности волнового распространения в воде радиофизики предпочитают анализировать не задачи прямой подводной связи, такие как электрокоммуникация у рыб, а ситуации, когда большая часть трассы связи приходится на воздушную среду. В этом случае возможно формирование и распространение радиоволн [12-16]. Речь идёт, например, о системах глобальной связи с погруженными подводными лодками или о системах управления торпедами, плывущими на относительно малых глубинах. Анализ таких задач позволяет применять стандартный радиофизический подход и рассматривать создание волн в воздухе, распространение их вдоль водной границы и проникновение под воду. Даже если антенны полностью зарыты в землю или погружены в воду,
они остаются радиофизическими - предназначенными для генерации и приёма радиоволн.
При анализе прямой подводной связи следует с большой осторожностью пользоваться некоторыми привычными понятиями волновой физики, такими как поток вектора Пойнтинга, точечный электрод и резонанс. В противном случае стереотипы радиофизической методологии приводят к неверной оценке возможностей гальванической связи (см., например, [17]). Так, широко распространённая физическая модель - диполь Герца, состоящая из двух маленьких шариков, разнесение которых значительно превышает их радиусы, - неадекватная модель для анализа возможностей связи на гальванических токах. И размеры электродов, и промежуток между ними должны быть соизмеримы с длиной антенны. Именно такие диполи применялись уже в первых моделях сова, созданных с учётом знаний о строении электрических органов рыб [18, 19]. В дальнейшем сотрудники ИПЭЭ РАН выполнили подробный физический анализ гальванической связи, предложили методики конкретных расчётов, провели многочисленные эксперименты в море, разработали и опробовали макеты связных устройств [20, 21]. В статье мы остановимся только на двух изученных проблемах - диаграмме направленности и оценке эффективности и оптимизации антенн для гальванической связи.
В отличие от поперечных радиоволн, при связи на гальванических токах продольная и поперечная компоненты поля соизмеримы, причём имеют разную зависимость амплитуды и фазы от частоты. Особенно заметно это проявляется на расстояниях, соизмеримых с толщиной скин-слоя, которые наиболее интересны, если мы хотим добиться максимальной дальности и информативности связи. В результате поле от линейного диполь-ного источника в большинстве случаев (кроме оси диполя и плоскости симметрии диполя) является эллиптически поляризованным, при этом размер, ориентация и форма каждого эллипса определяются расстоянием от источника и углом между направлением на него и осью диполя (рис. 3).
Зная структуру поля, можно предсказать, как будет меняться сигнал при изменении ориентации приёмной и передающей антенн - максимальный сигнал совпадёт с ориентацией приёмной антенны вдоль большой оси эллипса поляризации. На рисунке 3, в частности, видно, почему далеко не всегда справедливо утверждение одного из разработчиков устройств гальванической связи Н. МакЛеода: "Если приёмная антенна расположена под прямым углом к передающей, то наблюдается узкий ноль сигнала... чтобы достичь максимума дальности связи следует добиваться параллельности диполей" [10, с. 21]. Это так только для двух частных случаев: когда угол между осью
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
62
ОЛЬШАНСКИЙ и др.

передающего диполя и направлением на точку приёма равен 0 или 90°. Если же этот угол составляет 50-60°, а расстояние между приёмником и передатчиком сравнительно невелико, то ситуация обратная: параллельность антенн даёт минимум сигнала, а перпендикулярность - максимум. В случае же произвольных углов и расстояний в связи с эллиптической поляризацией электромагнитного поля всё обстоит гораздо сложнее, в частности ноль сигнала может отсутствовать вообще.
В выбранном диапазоне частот всё, что мы можем сделать для повышения дальности связи, - это добиться максимума дипольного момента Il, где I - ток в диполе, а l - эффективная длина диполя. Коэффициент, связывающий мощность (Р) в антенне с дипольным моментом, можно использовать в качестве критерия эффективности антенны: Il = √αeP. При этом речь идёт не о волновой мощности, а об используемом в электротехнике произведении квадрата силы тока на модуль импеданса (Р = I2Z). Отсюда αе = l2/Ze, то есть эффективность электрических антенн характеризуется отношением квадрата эффективной длины диполя к модулю импеданса. Чем больше разнесение электродов и чем ниже их сопротивление, тем эффективнее антенна, то есть тем больше величина сигнала, создаваемого антенной на удалении от источника при фиксированной мощности в источнике. Чтобы уменьшить импеданс, размеры электродов надо увеличивать. Однако если увеличение размера электродов производится за счёт уменьшения промежутка между ними, то это приводит к падению эффективной длины диполя. Оптимум, как правило, достигается, когда продольный размер электрода примерно равен величине промежутка между электродами. На низких частотах могут существенно сказываться сопротивление и ёмкость двойного слоя "проводник-вода". Для уменьшения их вклада в полный импе-
данс выбирают "правильный" материал электродов, например нержавеющую сталь или угольные ткани, а также полируют или увеличивают площадь поверхности электрода.
В начале 1980-х годов в Дальневосточном филиале НПО "Промрыболовство" была разработана система локальной связи между аквалангистами (руководитель группы разработчиков ёв) при консультативном участии сотрудников ИПЭЭ РАН [21, 22]. Было изготовлено пять комплектов, которые после успешных испытаний были переданы в опытную эксплуатацию. На акватории Тихого океана, на Чёрном и Карском морях аквалангисты использовали эту систему локальной связи при проведении исследовательских и судоремонтных работ, подводных киносъёмок в археологической экспедиции, а также в хозяйствах марикультуры. Дальность связи на глубинах до 10—15 м составляла 100-150 м. При увеличении длины антенны надводной станции до 300 м дальность односторонней связи "берег-вода" достигала 2 км. Связь осуществлялась на частотах речевого диапазона Гц. К преимуществам речевой связи на токах проводимости перед акустической связью относятся: лучшая разборчивость речи, отсутствие реверберации, сохранение связи при наличии между аквалангистами водорослей, подводных скал, аэрированных слоев воды. Малая зависимость (при дальности до 30 м) речевой связи от взаимной ориентации отмечалась во всех экспериментах и испытаниях. Уже в ближайшее время можно ожидать появления более эффективных устройств подводной токовой связи как для аквалангистов, так и для автономных подводных устройств, построенных с учётом возможностей современной электроники и новых материалов.
КОНТРОЛЬ ПЕРЕКОСОВ ТРАЛА
Дальность современных технических средств связи заметно превышает расстояния, на которых люди могут разговаривать без помощи техники. Поэтому вопрос о достижении больших дальностей представлялся главным для оценки практической значимости этого вида связи. При выработке инженерных рекомендаций мы исходили из необходимости излучать как можно более сильные и принимать как можно более слабые электрические сигналы. Соответственно, разрабатывая устройства связи, мы в гораздо большей степени опирались на физическую, а не бионическую методологию.
Вообще говоря, выполненный нами физический анализ, как и опыт регистрации слабых электрических и магнитных сигналов в морской и пресной воде, оказался полезен и для электроэкологических исследований. Так, с помощью аппаратуры, созданной в ходе выполнения приклад-
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
ЭЛЕКТРИЧЕСКИЕ РЫБЫ - БИОЛОГИЧЕСКИЙ ПРОТОТИП НОВОЙ ТЕХНИКИ 63
ных работ, были впервые зарегистрированы электрические разряды слабоэлектрических сомов непосредственно в месте их обитания - в озере Чамо (Эфиопия) [23]. Однако сама задача обеспечения максимальной дальности и информативности связи и такие её аспекты, как структура поля на расстояниях, превышающих толщину скин-слоя, или возможность использования поверхностной волны и длинных береговых антенн, большого интереса для исследований электрокоммуникации рыб не представляла. Скажем, мы выполнили множество экспериментов по регистрации электрических разрядов при парных взаимодействиях сомов Clarias macrocephalus, измеряя амплитуды электрических разрядов непосредственно на теле рыбы и на некотором расстоянии от неё [24]. Сопоставив эти результаты с данными об электрочувствительности сомов, можно было бы прийти к заключению о дальностях коммуникации больше 1 м. Однако реальные поведенческие ситуации, в которых наблюдался обмен разрядами, соответствуют только случаям, когда рыбы находятся почти вплотную друг к другу, преимущественно на расстоянии 1—4 см.
Современные модели электрических рыб строятся на основе детальных представлений о физиологических характеристиках и морфологических особенностях электрорецепторов, электрических органов и центральных структур, участвующих в обработке электросенсорной информации и управлении электрическими органами. Электро-ориентационные системы рыб представляют собой сложный многоуровневый комплекс, включающий множество электрических клеток, десятки тысяч рецепторов, сотни тысяч нейронов. Казалось бы, бионики должны стремиться к более полному использованию всех имеющихся сведений об электрических рыбах, например сразу начинать с оснащения подводных аппаратов многоэлектродными системами Р - и Т-рецепторов [25]. Но если уж рассматривать бионику как биологическую метафору, то надо честно следовать эволюционному подходу: начать с совсем простой модели и постепенно шаг за шагом наращивать её сложность. При этом надо всякий раз иметь перед глазами какую-нибудь конкретную цель. Поскольку самой близкой нам практической областью в конце 1980-х годов было рыбное хозяйство и промышленное рыболовство, то мы обратились к специалистам рыбной отрасли с просьбой назвать проблемы, наиболее актуальные и наименее удовлетворительно решённые. Первой была названа задача контроля перекосов трала.
Отечественные промысловые тралы имеют длину до 200 м, ширину до 77 м. Буксировка трала за рыболовным судном осуществляется с помощью ваеров - длинных (до 2 км) стальных тросов, каждый из которых закреплён на отдельном барабане траловой лебёдки, так что в процессе тра-

ления можно регулировать взаимную длину вытравленной части ваеров. Достоверно известно, что небольшие изменения взаимной длины (перетравки), вызывающие перекос орудий лова, могут резко снижать эффективность лова.
Мы решили, что с позиций бионики трал можно рассматривать как размещённую в безграничной проводящей среде большую прозрачную для тока оболочку с собственной координатной сеткой, в узлах которой можно расположить источники и приёмники электрического поля. В качестве основы для решения задачи контроля перекосов трала была взята простейшая четырёхэлектродная функциональная модель, предложенная в основополагающей статье Лиссманна "О функции и эволюции электрических органов рыб" [2] для иллюстрации принципа электролокации (рис. 4).
Согласно Лиссманну, основное назначение че-тырёхэлектродной модели - измерять нарушения симметрии. Эти нарушения могут возникать вследствие либо наличия каких-нибудь объектов вблизи электродов, либо смещений самих электродов относительно друг друга. Соответственно, для регистрации перекосов достаточно закрепить в точках, расположенных вдоль продольной оси трала, электроды, связанные с измерителем потенциала, а поперёк оси трала - электроды, соединённые с генератором тока. Пока регистрирующие электроды симметричны относительно излучающих, регистрируемое напряжение равно
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
64
ОЛЬШАНСКИЙ и др.

нулю, при перекосе регистрирующий электрод будет удаляться от одного из излучающих электродов и приближаться к другому. Соответственно, потенциал регистрирующего электрода будет определяться степенью перекоса, а фаза - знаком перекоса (вправо, влево).
Расчёты показывают, что если характерный размер электродов порядка 5 см, расстояния между соседними электродами 2 м, а разность потенциалов на выходе генератора тока 1 В, то изменение угла на 0.01° вызывает изменение регистрируемой разности потенциалов на 3 мкВ. Такая (и даже гораздо меньшая) разность потенциалов вполне может быть измерена без использования больших мощностей, громоздких антенных конструкций или сверхмалошумящих усилителей. При обработке сигналов целесообразно применять синхронное детектирование, теоретически обоснованное для слабоэлектрических рыб [2] и позже подтверждённое экспериментально для клюворылообразных [26].
Во многих случаях электроды следует размещать не вдоль и поперёк оси трала, а вдоль нитей, из которых связано сетное полотно. Это позволяет измерить усреднённый угол ромбов ячеи. При перекосах трала, например, влево левые ромбы ячеи сжимаются, а правые - вытягиваются. Соответственно, целесообразно разместить на трале два контролирующих устройства - слева и справа от продольной оси - и сопоставлять измеренные ими углы ромбов. При размещении трёх электродов на одной линии и жёстком закреплением расстояния между двумя из них можно контролировать растяжения фалов и нитей.
Созданные нами макеты системы контроля перекосов орудий лова (рис. 5) были испытаны в октябре 1985 г. на большом морозильном траулере "Мыс Бабушкина" при пелагических и донных тралениях в Татарском проливе и на сейнере "Ор-жев" Дальневосточной базы флота в октябре 1989 г. Испытания подтвердили высокую чувствительность этой системы к перекосам, наглядность отображаемой информации и её пригодность для управления тралом.
Общение с представителями рыбной отрасли как в России, так и за рубежом показывает, что заинтересованность в таких системах не только сохранилась, но даже возросла. Особенно актуальными эти средства контроля должны стать в связи с введением Европейским союзом ряда запретов на некоторые типы ячеи сетного полотна тралов. Тем не менее перспективы массового выпуска таких приборов в нашей стране остаются более чем туманными. Проблема упирается в традиционные трудности внедрения академических разработок в промышленность.
Новая система отношений между бюджетными и коммерческими структурами сегодня ориентирована на права собственности. Между тем авторские права и права собственности, возникающие на стадии оформления патентов и при получении начального финансирования на общие исследования, не учитывают интересов разработчиков действующих макетов и опытных образцов, которые только и могут довести идею "до ума". В результате при существующей ныне в Российской академии наук системе начисления баллов для оценки результативности научной деятельности сотрудников выгоднее бросить разработку сразу после получения патента и публикации общих идей, чем доводить её до товарного вида. Реформа академии, предполагающая повышение зарплаты только научным сотрудникам, делает ситуацию с инженерами и технологами ещё сложнее. Пока эти общие проблемы не решены, трудно ждать, что Академия наук будет эффективно выполнять функции инкубатора новой наукоёмкой продукции.
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
ЭЛЕКТРИЧЕСКИЕ РЫБЫ - БИОЛОГИЧЕСКИЙ ПРОТОТИП НОВОЙ ТЕХНИКИ 65
Если продолжить бионическую аналогию и сопоставить судьбу технических инноваций с проблемой возникновения новых биологических структур, таких как электрические органы и электрорецепторы, то при внедрении электро-ориентационных устройств мы сталкиваемся с той же проблемой, которая в литературе по слабоэлектрическим рыбам известна как загадка Дарвина, или проблема переходных форм. Как возникли электрические рыбы? Как на протяжении многих поколений складывалась эта сложная взаимоувязанная система рецепторов, электроци-тов и структур мозга, пока она не обрела конечную эффективность? Как переходная форма смогла преодолеть барьер выживания, оказаться конкурентоспособной по отношению к зрелым формам, вписаться в экосистему? К сожалению, в физико-технических вузах будущих инженеров не знакомят даже с общими идеями эволюционной теории и экологии. В результате от руководителей, которые формируют инновационную политику в стране, можно услышать, что всякая научно-исследовательская разработка должна заканчиваться опытно-конструкторской, а последняя - серийным устройством. Такой подход полностью противоположен идее естественного отбора. Нельзя гарантировать каждому сперматозоиду право стать человеком - земли не хватит. Вместе с тем низкая выживаемость молоди может быть следствием не только естественных процессов, но и проявлением неблагоприятной экологии, требующей корректировки среды. В переводе на технический язык это означает, что мы должны уметь анализировать, почему разработки, казавшиеся перспективными, остались не востребованы и почему бионика, популярная в конце 1960-х годов к 1980-м полностью сдала свои позиции в нашей стране. Одна из возможных причин - отсутствие стимулов и оценок для деятельности, заполняющей промежуток между уже известными научными идеями и работающими промышленными образцами.
ЗАДАЧА ПОСТРОЕНИЯ
ЭЛЕКТРОЛИТИЧЕСКОГО
ИНКЛИНОМЕТРИЧЕСКОГО СЕНСОРА
Инклинометрические сенсоры предназначены для измерения углового положения объектов относительно вектора силы тяжести. Они нужны в океанологии и океанографии, медицине, геофизике, топографии, компьютерных системах, строительстве, спорте, сельском хозяйстве, военной технике. Только в США с 1969 г. на различные варианты инклинометрических сенсоров выдано около 400 патентов. В России до 1993 г. ежегодно выдавалось от пяти до 15 авторских свидетельств, а после изменения патентного законодательства один-три. В мире выпускаются сотни модифика-

ции инклинометрических сенсоров и готовых инклинометров на их основе. Число компаний-производителей инклинометрических сенсоров исчисляется десятками, а общие тиражи изделий -миллионами.
Помогает ли инженерам при создании таких сенсоров знание результатов исследования электрических рыб? Прежде чем ответить на вопрос, рассмотрим современные модели слабоэлектрических рыб.
Две группы электрических рыб в наибольшей степени используют электрические поля в информационных целях: это южноамериканские гимнотообразные, например Apteronotus albifrons, генерирующие разряды волнового типа (рис. 6, а), и африканские клюворылообразные, например Gnathonemus petersil, генерирующий разряды пульсирующего типа (рис. 6, б). Хотя функциональная модель Лиссманна справедлива для всех представителей названных отрядов, разработаны и более детальные модели, учитывающие видовую принадлежность и тип разрядов рыбы, в частности, модели В. Хейлигенберга для волновых гимнотообразных [27, 28] и К. Белла для пульсирующих клюворылообразных [26, 29].
У этих моделей много общего, поскольку много общего в строении, физиологии и функционировании электросистем у гимнотообразных и клюворылообразных. Каждая особь, относящаяся к этим двум отрядам, имеет электрорецепторы двух морфологических типов - ампулярные и бугорковые. Они заметно различаются по своим
5 ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
66 ОЛЬШАНСКИЙ и др.
функциональным характеристикам. Более чувствительные ампулярные предназначены для восприятия низкочастотных стимулов длительностью порядка десятков миллисекунд и более. К таким стимулам относятся, в частности, неспециализированные проявления электрической активности водных организмов, например элек-тропневмограммы - электрические токи, сопровождающие дыхание рыб. Бугорковые рецепторы предназначены для восприятия специализированных электрических разрядов - своих собственных для целей локации и чужих для целей коммуникации. Бугорковые рецепторы делятся на несколько функциональных типов, причём у каждой рыбы есть кодировщики амплитуды и кодировщики времени. Число электрорецепторов у одной особи исчисляется десятками тысяч.
Одним из основных различий между моделью Хейлигенберга, построенной для гимнотообраз-ных, и моделью Белла, построенной для клюворыл ообразных, является отсутствие в первом случае и наличие во втором специальной связи между командным ядром, управляющим запуском электрического разряда, и электросенсорной зоной, осуществляющей первичную обработку нервных импульсов, которые поступают от электрорецепторов. У клюворылообразных между командным центром и электросенсорной долей существует прямая связь, по которой поступают импульсы, блокирующие или, напротив, усиливающие ответы рецепторов разных типов во время собственных разрядов. Таким образом, задачи локации и коммуникации разносятся во времени. У гимнотообразных нет прямой центральной связи между командным ядром и электросенсорной долей боковой линии. При этом происходит трансформация исходного сигнала в образы на амплитудно-фазовой плоскости, которые строятся на соматотопических структурах электросенсорной зоны мозга.
Современные модели электрогенерации у рыб также заметно отличаются от простых диполь-ных источников поля. В первых моделях Лиссманна электрические органы описывались как диполи, причём особое внимание обращалось на механизмы, обеспечивающие синхронность возбуждения электроцитов. Но позже было показано, что по крайней мере у некоторых электрических рыб, в первую очередь гимнотообразных, электрогенераторные структуры возбуждаются поблочно, или посегментно. За счёт посегментно-го срабатывания форма разряда разная в разных точках поверхности рыбы, и вектор напряжённости поля за время генерации одного разряда может заметно изменить направление. Это приводит к возникновению фазовых сдвигов, что особенно важно для модели Хейлигенберга, согласно которой воспринимаемый электрический стимул
трансформируется в образ на амплитудно-фазовой плоскости.
Итак, современные модели электрических рыб полифункциональны, приёмные системы включают тысячи разнесённых приёмников, источники имеют сложную структуру, в мозге осуществляется параллельная обработка образов. И хотя по частотным характеристикам электрорецепция ближе к акустике, её чаще называют "электровидение", а систему электрорецепторов со времен Лиссманна именуют "электрической сетчаткой". При этом электрические образы строятся с учётом информации от всех сенсорных систем, включая зрение и проприорецепцию [26].
Вернёмся к практической задаче разработки инклинометрического сенсора. Он содержит нечто, что смещается при изменении углового положения сенсора относительно вектора силы тяжести, например маятник, пружинку или пузырёк воздуха в жидкости, а также некое устройство, преобразующее это смещение в электрический сигнал. Наиболее распространены электролитические инклинометры, имеющие полость, заполненную проводящей жидкостью с пузырьком или маятником и систему электродов. Внутри полости возбуждают электрическое поле и по величине сопротивления между электродами или по величине потенциала на регистрирующем электроде определяют угол отклонения пузырька либо маятника.
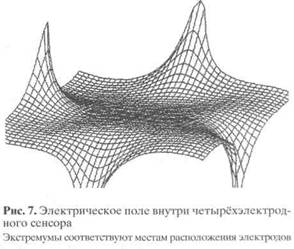
Если сравнивать существующие и заявленные в патентах конструкции инклинометрических сенсоров, то конкретные недостатки одних решений устранены в других, однако два главных недостатка всё ещё не преодолены. Первый - нелинейная зависимость выходного сигнала от угла наклона и заметное ухудшение точности при расширении диапазона измеряемых углов. Этот недостаток сказывается не только в применениях, требующих полного диапазона углов - от нуля до 360°, но и при создании любых векторных сенсоров, если в их состав входят два или три одномерных сенсора, оси чувствительности которых взаимно ортогональны. Производитель должен в этом случае обеспечить при изготовлении высокую точность ортогональности, что для чувствительности порядка 0.01° сделать довольно трудно. Нелинейность зависимости электрического сигнала от угла обусловлена неоднородностью электрического поля, создаваемого в полости сенсора (рис. 7). Для преодоления этого недостатка нелинейность либо компенсируют, либо вводят дополнительные электронные узлы с корректирующей характеристикой или математические поправки, учитываемые процессором при переводе потенциала в угол.
Второй недостаток состоит в том, что в существующих сенсорах угол наклона преобразуется в
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
ЭЛЕКТРИЧЕСКИЕ РЫБЫ - БИОЛОГИЧЕСКИЙ ПРОТОТИП НОВОЙ ТЕХНИКИ 67
величину, зависящую не только от этого угла. Определяем ли мы, как изменяются сопротивление в случае проводящих жидкостей или ёмкость в случае диэлектрических жидкостей, либо измеряем амплитуду потенциала на регистрирующем электроде - всегда измеряемая величина зависит от геометрии жидкости внутри полости и изменений её физических свойств, которые в свою очередь зависят от температуры и давления в сенсоре. Разумеется, все эти зависимости можно в какой-то степени учесть и компенсировать, но в идеальном варианте хотелось бы, чтобы их не было изначально.
Для построения инклинометрического сенсора, лишённого названных недостатков, возьмём в качестве базовой модель Хейлигенберга для рыб с волновым типом разряда. Согласно этой модели, воспринимаемые электрические поля трансформируются в образы на амплитудно-фазовой плоскости. При этом электрические рыбы демонстрируют удивительно высокое разрешение по фазе: они чувствуют микросекундные задержки ритма генерации при среднем периоде собственных разрядов в десятки миллисекунд [30]. С точки зрения современной техники микросекундное (и даже пикосекундное) разрешение интервала времени не кажется фантастическим, но у рыбы все нервные процессы происходят с характерными временами около единиц миллисекунд, поэтому чувствительность к микросекундным изменениям впечатляет.
Напомним, что в волновой физике временные задержки, или набеги фазы, определяются скоростью распространения прямого и отражённого сигналов от источника до цели, а затем от цели к приёмнику. При статической локации в проводящей среде этими задержками распространения можно пренебречь. В инклинометрах фаза сигнала на приёмном электроде либо совпадает с фазой сигнала на токовом электроде, либо отличается на 180°. Откуда берётся набег фазы у электрических рыб?
Два ответа на этот вопрос были предложены в гг. Первый - полный импеданс как самой рыбы, так и лоцируемых ею биологических объектов имеет соизмеримую резистивную и ёмкостную компоненты. При изменении положения объекта относительно источника действительная и мнимая части импеданса меняются по-разному и, соответственно, меняется фаза сигнала в точках регистрации. Таким образом, в простой четы-рёхэлектродной модели мы имеем дело не с активным четырёхплечим мостом и четырьмя сопротивлениями, а с мостом переменного тока и комплексными импедансными нагрузками. Второй ответ на вопрос о причинах набега фазы - суперпозиция собственных полей рыбы с близкими по частоте разрядами других рыб. Именно бие-
ния, возникающие при суперпозиции электрических разрядов от двух близко расположенных волновых электрических рыб, позволили Хейли-генбергу создать и экспериментально исследовать модель, описывающую реакции избегания помех, и установить роль различных структур мозга в обработке электросенсорной информации. Но если речь идёт о локации пассивных объектов, не создающих собственных электрических полей, то говорить о фазовых набегах за счёт суперпозиции не приходится.
В последние годы активно обсуждается третья причина фазовых набегов - сложное устройство электрического органа рыб, который состоит из множества электрических клеток - электроцитов. Каждый электроцит имеет отдельную иннервацию, и в зависимости от того, насколько одновременно поступают нервные импульсы, электроциты генерируют разряд либо синхронно, либо с задержками относительно друг друга [31, 32]. В публикациях 70-х годов прошлого века обсуждались механизмы, обеспечивающие одновременность срабатывания электроцитов, но отмечались "несовершенства" устройства электрических органов. В 90-х годах этим "несовершенствам" было дано функциональное объяснение: разные фазы разряда формируются разными сегментами электрического органа [33, 34], размеры электрического органа соизмеримы с размером рыбы и расстоянием до мишени. В процессе разряда различные сегменты срабатывают последовательно, и рыба как бы сканирует окружающее её пространство. Поскольку лоцируемый объект находится на разном удалении от разных сегментов электрического органа, объект в зависимости от своего местоположения по-разному искажает разные фазы сигнала. Следовательно, информация о фазе искажения становится не менее важной, чем информация об амплитудных изменениях. При этом фазовая информация в гораздо
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
5*
68
ОЛЬШАНСКИЙ и др.
|
меньшей степени подвержена влияниям параметров внешней среды, например изменениям электропроводности воды.
Все эти идеи могут быть привлечены для создания новой концепции построения электролитических инклинометрических сенсоров, которая предусматривает следующие шаги:
• обеспечение максимальной симметрии;
• создание распределённого источника тока, который обеспечивает пространственное сканирование области возможного нахождения объекта (пузырька газа или металлического маятника в электролите, заполняющем полость сенсора) и вызывает соответствующие изменения не только амплитуды, но и фазы регистрируемого сигнала;
• трансформирование искажения поля в образ на амплитудно-фазовой плоскости и разделение фазы и амплитуды, при этом точностные характеристики должны улучшаться за счёт пространственной и временной интеграции регистрируемых сигналов.
Наиболее очевидный путь обеспечения симметрии - выполнение полости в виде цилиндра и симметричное расположение электродов относительно его оси. Этот принцип широко используется в инклинометрах. Для построения сегментированного источника нужно взять несколько электродов и последовательно подключать их к источнику тока. Чем больше электродов, тем более плавно меняется фаза. Очевидно, что, в отличие от электрических рыб, мы можем использовать полный диапазон фаз в 360° и равномерно распределить фазы между всеми токовыми электродами. В этом случае в полости сенсора поле будет вращаться относительно оси симметрии. Если бы внутри полости не было неоднородностей (пузырька или маятника), то вследствие симметрии в центре полости мы бы всегда имели нулевой потенциал. Наличие неоднородности приводит к тому, что симметрия нарушается и потенциал регистрирующего электрода становится отличным от нуля, причём фаза потенциала
соответствует угловому положению неоднородности.
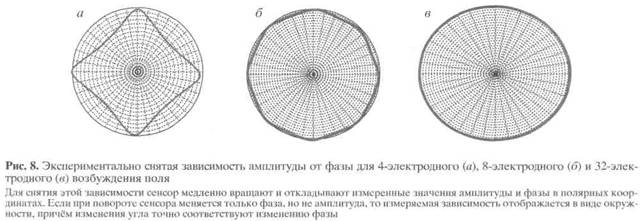
Развиваемый подход позволяет сформулировать понятие идеального инклинометрического сенсора. У такого сенсора, если его вращать вокруг оси симметрии, на регистрирующем электроде меняется только фаза, но не амплитуда потенциала. Наклоны идеального инклинометрического сенсора в плоскости, проходящей через его ось симметрии, могут приводить к изменению амплитуды, но не фазы напряжения на регистрирующем электроде. Чем больше число электродов, тем ближе к идеалу зависимость амплитуды и фазы от угла (рис. 8).
Однако значительное увеличение числа электродов означает качественное усложнение электронной части макетов. В макетах с двумя или четырьмя токовыми электродами мы использовали встроенные цифроаналоговые преобразователи дешёвых микроконтроллеров. Их быстродействия, как и разрядности, вполне достаточно для традиционных технических решений. Если же мы применяем 32 электрода и, соответственно, генерируем 32 управляющих сигнала, например 32 синусоиды, то мы должны иметь 32-канальный цифроаналоговый преобразователь. Увеличение числа электродов сопряжено с уменьшением размеров каждого из них. Чем меньше электрод, тем сильнее влияние электрохимических процессов на границе "электрод-электролит" на результаты измерения, в том числе на стабильность фазы сигнала. Чтобы ослабить это влияние, надо применять достаточно высокие рабочие частоты - в десятки килогерц. Пусть, например, мы хотим обеспечить рабочую частоту около 30 кГц (период около 32 мкс). Если каждый период синусоиды мы формируем по 32 точкам, то каждую точку мы должны выставить за 1 мкс. Если у нас 32 токовых электрода и если мы выставляем напряжения на них последовательно, то на обслуживание одного токового электрода у нас остается только 30 не, причём за это время нужно также провести
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
ЭЛЕКТРИЧЕСКИЕ РЫБЫ - БИОЛОГИЧЕСКИЙ ПРОТОТИП НОВОЙ ТЕХНИКИ 69
|
измерение сигнала и его предварительную обработку на регистрирующем электроде. В такой ситуации целесообразно перейти от последовательных устройств к параллельным, например к программируемым логическим интегральным схемам, выпускаемым с использованием нанотехнологий. В наших макетах мы применяли базовые матричные
микросхемы Spartan-З фирмы "Xilinx", изготовленные по технологии 90 нм.
Какова конечная цель, оправдывающая эти сложности? Она очевидна - борьба за динамический диапазон в 7, а в перспективе в 9 десятичных порядков. Современные инклинометры имеют
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
70
ОЛЬШАНСКИЙ и др.
|
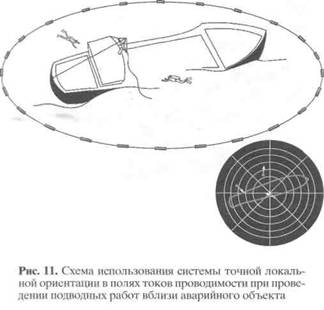
ную систему последовательно возбуждаемых электродов разместить в открытой водной среде, то можно создать вращающееся локальное поле гальванических токов, например, в месте проведения аварийно-спасательных или подводных археологических работ (рис. 11), чтобы обеспечить точную ориентацию подводных пловцов и маркировку позиций найденных объектов. При этом каждый элемент такой системы может быть простым и дешёвым.
Подавая на излучающие электроды ориента-ционной системы единицы вольт и регистрируя на приёмных электродах единицы нановольт (тепловые шумы составляют доли нановольта) мы можем расширить динамический диапазон до 9 десятичных порядков по амплитуде. Используя частоты в десятки герц и измеряя фазу с точностью до долей наносекунды, мы также имеем около 9 порядков. Таким образом, можно рассчитывать на высокую точность определения положения в системах токовой электроориентации.
точность около 0.01°, то есть динамический диапазон в 4-5 десятичных порядков. Мы хотим выйти на точности не хуже долей угловой секунды в полном диапазоне углов - от нуля до 360°. Это означает, что мы должны обеспечить соответствующие точности измерения фазы.
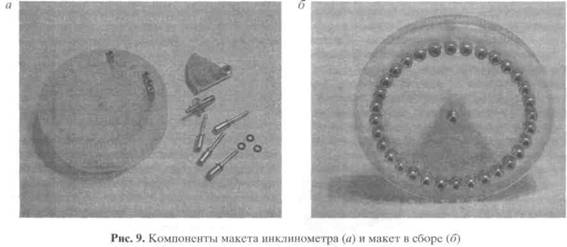
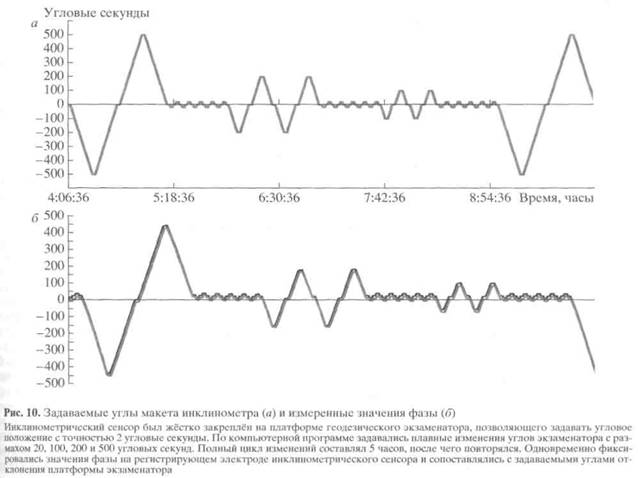
Механическая часть сенсора также должна изготавливаться с применением нанотехнологии. В наших макетах использовались маятники, закреплённые на оси на подшипниках (рис. 9). Керамические шарики этих подшипников имеют шероховатость порядка 3.0-3.6 нм, шероховатость обойм подшипника в 10 раз больше. Для наружной обоймы, имеющей радиус около 8 мм, отклонение на один градус соответствует 70 мкм, а на одну угловую секунду - 20 нм, то есть меньше, чем шероховатость обоймы. Из-за этого в экспериментах, в которых мы задавали малые угловые отклонения (единицы-десятки угловых секунд), мы наблюдали отчётливый гистерезис и нарушение соответствия "угол наклона = угол фазы" (рис. 10). Иными словами, после того, как мы научились измерять положение маятника относительно электродной системы с точностью единиц угловых секунд, на первый план вышла проблема механического несоответствия положения маятника направлению вектора силы тяжести. За этими техническими подробностями стоит вопрос о том, какие физические ограничения определяют порог разрешения при электростатической локации и ориентации.
Инклинометрический сенсор мы рассматривали в качестве частного случая ориентационной системы для проводящих сред. Если распределён-
* * *
В истории науки и техники было немало случаев, когда эксперименты биологов требовали кардинальных изменений существовавших физических представлений, некоторые из них приведены в начале статьи. Но и тогда, когда обсуждаемые в биологии модели апеллируют к сложившимся представлениям, в бионический анализ вовлекаются вопросы, которые обычно остаются в стороне при чисто физическом рассмотрении, например, вопросы цели, целостности, эволюции и экологии.
При физическом анализе цель возможного применения рассматриваемой модели вносится извне или может вообще не обсуждаться. Для электроэкологов назначение электрического органа или ампулярных рецепторов - объект исследования. При этом биологическая структура могла первоначально иметь одну функцию, а потом заменить её на другую. Скажем, ампулярный орган мог на ранних стадиях эволюции возникнуть как механорецептор, а потом "переквалифицироваться" в электрорецептор. У физической модели может не быть предыстории или будущего развития. В биологии объединённая электрорецепторно-электрогенераторная система рассматривается в эволюционном аспекте. Физическая модель может представлять собой одно элементарное целое, любая биологическая система состоит из взаимоувязанных частей и сама является частью более общей системы. Физическая модель может быть представлена в воображаемом пустом пространстве или очень упрощённой среде, биологическая всегда относится к конкретной группе животных, обитающих в реальном мире в конкретной экологической нише, имеющей свою специфику. Все эти вопросы близки инженерам, которых ин-
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
ЭЛЕКТРИЧЕСКИЕ РЫБЫ - БИОЛОГИЧЕСКИЙ ПРОТОТИП НОВОЙ ТЕХНИКИ 71
тересуют и возможная смена функционального назначения, и принципы стыковки частей в целое, и рекомендации по совершенствованию на основе предыстории разработок, и привязка разработки к условиям её будущей эксплуатации.
Настаивая на перспективности бионической методологии, мы исходим из того, что в физиологических моделях заложен огромный запас идей и возможностей по сравнению с теми техническими макетами, которые мы реально сегодня можем сделать. Приведённые в статье примеры технических разработок, очевидно, проще электро-ориентационных систем слабоэлектрических рыб, но, тем не менее, даже в таком виде наши разработки имеют явную практическую значимость. Большое количество рецепторов, электроцитов и нейронов, участвующих в обработке электросенсорной информации, обеспечивают биологическим системам качественные преимущества по сравнению с их малоэлектродными аналогами. Даже на простой модели инклинометрического сенсора отчётливо видно, как много дают увеличение числа электродов и параллельная обработка информации. Между тем 32 электрода инклинометра - это не десятки тысяч рецепторов "электрического глаза" рыбы. Технические модели по крайней мере на три порядка уступают своим биологическим прототипам и по чувствительности - электрорецепторы воспринимают токи порядка долей наноампера, а в наших моделях пока надёжно используются только микроамперы. С первых экспериментальных исследований электрорецепции стало ясно, что возможность достижения физических пределов электровосприятия сопряжена с особенностями организации сенсоров на молекулярном уровне - с тем, что рыба умеет "считать" электроны и ионы поштучно.
Соответственно, перспективы создания многоэлектродных комплексов, гораздо более приближённых к натуральным системам электровидения рыб, чем описанные выше, связаны в первую очередь с прогрессом в области электроники и нанотехнологий. Живые аналоги могут служить путеводной звездой, в какой-то степени позволяя "подсмотреть ответ" или воспользоваться рыбой как готовым "наноизделием", подобно тому как во времена Гальвани лягушки выступали в роли гальванометра, заменяя не существовавшие тогда микроамперметры и приближая время их создания.
Работа выполнена при поддержке гранта РФФИ и программы "Ведущие научные школы".
ЛИТЕРАТУРА
1. Lissmann H. W. Continuous electric signals from the tail offish, Gymnarchus niloticus // Nature. 1951. V. 167. P. 201.
2. Lissmann H. W. On the function and evolution of electric organs in fish // Journ. Exp. Biol. 1958. V. 35. P. 156-191.
3. Lissmann H. W., Machin K. E. The mechanism of object location in Gymnarchus niloticus and similar fish // Journ. Exp. Biol. 1958. V. 35. P. 451-486.
4. Cavendish H. An account of some attempts to imitate the effects of the Torpedo by electricity // Philos. Trans. Roy. Soc. 1776. V. 66. P. 196-225.
5. Piccolino M., Bresadola. Drawing a spark from darkness: John Walsh and electric fish // Trends in Neuro-sciences. 2002. V. 25. P. 51-57.
6. Piccolino M. The bicentennial of the Voltaic battery (): the artificial electric organ // Trends in Neurosciences. 2000. V. 23. P. 147-151.
7. Об электричестве, возбуждаемом простым соприкосновением различных проводящих веществ. Письмо к сэру И. Банксу от 01.01.01 г. // ММ. Радовский. Гальвани и Вольта. М.-Л.: ГЭИ, 1941.
8. Заключение о характере направления электрической силы у электрического угря // Экспериментальные исследования по электричеству. Т. 2. М.-Л.: ОГИЗ, 1951. С. 28.
9. Conover E. A. Experimental conduction current signal system // IEEE Int. mun. Digest. Minneapolis: Minn, 1967.
10. MacLeod N. Electric diver communication: Non acoustic system can operate in noisi environments // Sea Technol. 1977. May. P. 21,48.
11. Bogie I. S. Conduction and magnetic signalling in the sea. A background rev. // Radio and Electron. Eng. 1972. V. 42. P. 447-^152.
12. KraichmanM. B. Handbook of electromagnetic propagation in conducting media // Wash. (D. C.): US Gov. print, off., 1970.
13. Rowe H. E. Extremely low frequency (ELF) communication to submarines //IEEE mun. 1974. V. 22. P. 371-385.
14. Burrows ML. ELF communication antennas. Peregri-nus, 1978. P. 80.
15. Bannister P. R. New simplified formulas for ELF subsur-face-tosubsurface propagation // IEEE Journ. Ocean. Eng. 1984. V. 9. P. 154-163.
16. Антенны в материальных средах. В 2 т. М.: Мир, 1984.
17. Остроумов ГЛ. На каком расстоянии морские животные могли бы общаться с помощью электромагнитных волн // Дифракция и распространение радиоволн. 1968. Вып. 8. С. 3-24.
18. , , Бионическое моделирование электрических коммуникационных и локационных систем рыб // Вестник АН СССР. 1981. № 1.
19. , Бондарчук AM., Введение в электроэкологию. М.: Наука, 1982.
20. Бионическое моделирование электросистем слабоэлектрических рыб. М.: Наука, 1990.
21. , П. Разработка систем локальной подводной связи на гальванических токах // Нелинейный мир. 2007. № 12.
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
72
ОЛЬШАНСКИЙ и др.
22. Переговорное устройство для легководолазов // Спортсмен-подводник. 1982. Вып. 68. С. 30-33.
23. , С, и др. Наблюдения электрической активности силуриевых сомов (Siluriformes) в оз. Чамо (Эфиопия) // Вопросы ихтиологии. 2001. Т. 41. С. 542-549.
24. , С, Насека A.M., Нгуен Тхи Нга. Электрические разряды кларие-вых сомов, культивируемых в Южном Вьетнаме // Вопросы ихтиологии. 2002. Т. 42. С. 549-557.
25. Nelson M. E. Target Detection, Image Analysis and Modeling // Electroreception / Eds. Bullock Т. Н., Hopkins CD., Popper A. N., Fay R. R. N. Y.: Springer, 2005.
26. Интеграция периферических и центральных путей в электросенсорных долях мозга мормиридовых рыб // Сенсорные системы. 1994. Е. 8. С. 233-244.
27. Heiligenberg W. Jamming avoidance responses model systems for neuroethology // Electroreception / Eds. Bullock Т. Н., Heiligenberg W. N. Y.: Wiley, 1986.
28. Heiligenberg, W. Neural Nets in Electric Fish. Cambridge, MA: MIT Press., 1991.
29. Bell C. C. Electroreception in mormyrid fish: central physiology // Electroreception / Eds. Bullock Т. Н., Heiligenberg W. N. Y.: Wiley, 1986.
30. Rose G. J., Heiligenberg W. Temporal hyperacuity in the electric sense of fish//Nature. 1985b. V. 318. P. 178-180.
31. Электрогенераторные системы рыб: эволюция и механизмы адаптации. М.: Наука, 1982.
32. Барон В Д. Возможная роль электрорецепции в поведении морских слабоэлектрических скатов Raja clavata (Rajidae) // Сенсорные системы. 1994. Т. 8. С. 147-161.
33. Schuster S. Changes in electric organ discharge after pausing the electromotor system of Gymnotus carapo // Journ. Exp Biol. 2000. V. 203. P. .
34. Schuster S., Otto N. Sensitivity to novel feedback at different phases of a gymnotid electric organ discharge // Joum. Exp. Biol. 2002. V. 205. P. .
![]()
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ НАУК том 79 № 1 2009
