
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Институт биоорганической химии им. академиков и Российской академии наук,
лаборатория сравнительной и функциональной геномики
Московская гимназия на Юго-западе № 000
Определение нарушений структуры протоонкогена AML1 в лейкоцитарных клетках больных острым лимфобластным лейкозом
Карнаева Анастасия
Джафарова Ася
Научный руководитель:
д. б.н.
Москва 2013
СОДЕРЖАНИЕ
Введение 3
1. Обзор литературы 4
Список цитируемой литературы 7
2. Материалы и методы
2.1. Материалы и реактивы 9
2.2. Методы
2.2.1. Полимеразная цепная реакция 12
2.2.2. Электрофорез 12
2.2.3. Лигирование 13
2.2.4. Трансформация 13
2.2.5. Выделение плазмидной ДНК 13
2.2.6. Приготовление ночной культуры 14
2.2.7. Получение ДНК для скрининга 14
2.2.8. Секвенирование ДНК 15
3. Результаты и их обсуждение 16
3.1.Проведение ПЦР-анализа наличия транслокаций t(12;21) 16
3.2. Клонирование фрагментов кДНК TEL/AML1 18
3.3. Проведение анализа первичных структур кДНК TEL/AML1 21
3.4. Заключение 23
Выводы 24
Благодарности 25
Введение
Лейкозы – обширная группа онкологических заболеваний кроветворной системы, зарождающиеся в костном мозге. При лейкозах недифференцированные предшественники кровяных клеток бесконтрольно делятся на более ранних стадиях гемопоэза, хотя в норме неспециализированные клетки не размножаются до тех пор, пока не прошли дифференцировку. При острых лейкозах происходит наиболее интенсивное деление (пролиферация) таких клеток в костном мозге и их массированный выход в кровоток. Эти клетки не способны функционировать так, как нормальные зрелые клетки крови. Уменьшение числа и утрата функций нормальных клеток приводят к анемии, подавлению иммунитета (и, как следствие, к инфекционным заболеваниям), внутренним кровотечениям, а циркуляция в организме недифференцированных клеток приводит к появлению лейкозных инфильтратов в различных органах.
Детские формы лейкоза делятся на острый лимфобластный (ОЛЛ) и острый миелобластный (ОМЛ) лейкозы. Предполагается существование многих генетических причин возникновения ОЛЛ. Одними из наиболее характерных для данного заболевания являются хромосомные транслокации, наблюдаемые в опухолевых клетках. Мы исследуем одну из подобных генетических причин заболевания. Мы рассматриваем транслокацию t(12;21)(p13;q22). Она характерна примерно для 25% случаев детского В-клеточного ОЛЛ (В-ОЛЛ). Она приводит к образованию химерного гена TEL/AML1. Ген AML1 кодирует ключевой транскрипционный фактор, регулирующий экспрессию целой группы генов кроветворения. Основными мишенями фактора AML1 служат гены, кодирующие белки дифференцировки лимфоидных клеток. Продукт гена TEL действует как транскрипционный регулятор; белок TEL важен в ангиогенезе и в гемопоэзе. Следовательно, любые хромосомные перестройки, в которых участвует этот ген, могут приводить к нарушению процесса дифференцировки.
Цель нашей работы заключается в определении первичной структуры транскриптов химерных генов TEL/AML1 в образцах лимфобластных клеток, полученных от нескольких пациентов с ОЛЛ.
Для достижения поставленной цели нам было необходимо решить следующие экспериментальные задачи:
1) Проведение ПЦР-анализа наличия транслокаций t(12;21) в образцах лимфобластных клеток пациентов, больных острым лимфобластным лейкозом;
2) Клонирование фрагментов транскриптов в E. coli; и секвенирование рекомбинантных клонов
3) Проведение анализа первичных структур кДНК TEL/AML1.
Обзор литературы
Химерный ген TEL/AML1 – генетический маркер острого лимфобластного лейкоза
Одним из признаков острого лимфобластного лейкоза (ОЛЛ) является t(12;21) транслокация, которая приводит к слиянию генов TEL и AML1.
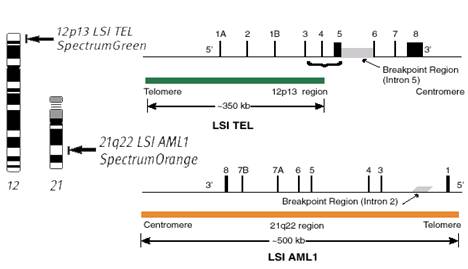
Структура обоих генов и общая схема данной транслокации представлена на рисунке 1. Ген AML1 (находится в 21 хромосоме), длиной 150 т. п.о., включает в себя 8 экзонов. Ген TEL (в 12 хромосоме) состоит из 240 т. п.о, включает 8 экзонов. В гене TEL место разрыва находится между пятым и шестым экзоном, в гене AML1 – между вторым и третьим. В результате транслокации гены «сшиваются» на месте разрыва.
Рисунок 1. Общая схема транслокации t(12;21)
![]()
 - указание локусов хромосом, содержащие рассматриваемые гены.
- указание локусов хромосом, содержащие рассматриваемые гены.
12, 21 – номера хромосом.
1А, 2, 1В и т. д. – номера экзонов.
![]() - область разрыва хромосомы.
- область разрыва хромосомы.
![]() - ген TEL.
- ген TEL.
![]() - ген AML1.
- ген AML1.
Такие ненормальные хромосомы встречаются в 25% случаях детского В-ОЛЛ. Таким образом, присутствие таких хромосом можно считать признаком заболевания, поэтому в клинике при первичной диагностике проверяют наличие таких транслокаций с помощью цитогенетического анализа. Пример такого анализа представлен на рисунке 2 .
Хромосомы делящихся клеток окрашиваются методом FISH (Флюоресцентная гибридизация in situ), При флуоресцентной гибридизации in situ используют ДНК-зонды, которые связываются с комплементарными участками хромосомы. В нижнем левом углу картинки приведен пример нормальной клетки, у которой разные зонды находятся на разных хромосомах. Белые стрелки указывают на ненормальные хромосомы, у которых гены TEL и AML1 расположены на одной такой хромосоме.
Рисунок 2. Пример результатов цитогенетического анализа
![]()
![]()
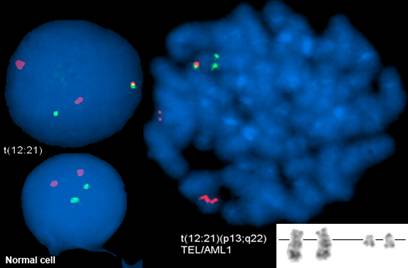
Красная флуоресцентная метка находится на зонде, содержащий ген TEL.
Зеленая флуоресцентная метка находится на зонде, содержащий ген AML1.
Стрелки указывают на продукты транслокации.
Рассмотрим более подробно некоторые свойства обоих генов.
AML1 (RUNX1) относится к семейству белков RUNX, к которому также относят транскрипционные факторы RUNX2 и RUNX3 (они же AML2 и 3 соответственно). Подобная “двойственность” в названиях неслучайна и связана, в первую очередь, с историей изучения данного семейства белков. Первым был идентифицирован ген, участвующий в хромосомных перестройках, ассоциированных с острым миелоидным лейкозом человека. Отсюда происходит его первое название, AML1, акроним английского Acute Myeloid Leukemia. В ходе дальнейшей работы установили, что продукт этого гена – транскрипционный фактор, содержащий ДНК - связывающий домен runt, отсюда появилось второе название, RUNX1,акроним английского Runt related transcription factor 1. Два других фактора идентифицировали позже, что позволило добавить соответствующие порядковые номера к их названиям. Эволюционно консервативные факторы RUNX принимают участие в регуляции таких важнейших процессов в организме млекопитающих, как гемопоэз (RUNX1), остеогенез (RUNX2), нейрогенез, тимопоэз и регуляция пролиферации клеток эпителия желудка (RUNX3). Каждый из белков RUNX кодируется только одним геном, но в результате альтернативного сплайсинга образуются его различные изоформы. Экспрессия каждого из трех генов RUNX у позвоночных регулируется двумя промоторами: дистальным P1 и проксимальным P2. Однако до сих пор не известно, действует ли каждый из факторов на свой промотор, поддерживая стабильный уровень экспрессии гена или подавляя активность промотора при высокой концентрации белка, или каждый из факторов связывается с промоторами генов других представителей семейства RUNX, подавляя их экспрессию. Транскрипты, синтезированные с промоторов P1 и P2, подвергаются альтернативному сплайсингу, что приводит к синтезу разнообразных изоформ белков RUNX в разных клетках на разных стадиях цитокинеза. В настоящий момент большое внимание уделяется генам RUNX, так как с нарушением их структуры и регуляции экспрессии связано множество патологий. В частности, хромосомные перестройки, затрагивающие ген RUNX1, обнаружены у 25% больных острым миелоидным и В-лимфобластным лейкозом. У 3% больных лейкозом находят мутации в этом гене, которые приводят к замене аминокислотных остатков или к сдвигу рамки считывания, а, следовательно, к изменению структуры белка, в том числе ДНК-связывающего и трансактивационного доменов.
Ген RUNX1 (AML1) содержит 8 экзонов. Экзоны 2, 3 и 4 гена RUNX1 кодируют ДНК-связывающий домен (runt-домен), а шестой экзон кодирует трансактивационный домен (ТА-домен). В результате альтернативного сплайсинга образуются мРНК, кодирующие различные изоформы RUNX1. На сегодняшний день известно четыре варианта этого транскрипционного фактора: RUNX1a, RUNX1b, RUNX1c и RUNX1ΔN.
На работу фактора RUNX1 большое влияние оказывают белки, вовлеченные в регуляцию клеточного цикла. RUNX1 содержит три сайта, фосфорилируемых циклин-зависимыми киназами. Транскрипционный фактор RUNX1 принимает участие в регуляции таких важнейших процессов, как гемопоэз, ангиогенез, нейрогенез и работа скелетных мышц. На сегодняшний день известно большое число генов, в регуляции экспрессии которых принимает участие белок RUNX1. В промоторной области всех этих генов находится сайт связывания RUNX1. Обычно именно этот сайт определяет тканевую специфичность экспрессии того или иного гена, однако, в составе гетерологичного промотора не изменяет тканевую специфичность экспрессии контролируемого им гена, свидетельствуя о том, что RUNX1, скорее всего, выполняет функцию “организатора” транскрипции, привлекая тканеспецифичные факторы к сайтам их связывания в промоторах [1,2]. Нарушение работы RUNX1 часто наблюдается при патологии кроветворной системы, поэтому в первую очередь изучали участие этого фактора в процессах гемопоэза. RUNX1 активен в лимфоидных, миелоидных и эритроидных клетках. Он контролирует экспрессию генов цитокинов, клеточных рецепторов, регуляторов клеточного цикла. В В-лимфоцитах RUNX1 активирует экспрессию генов, кодирующих IgA1 и тирозинкиназу BLK, стимулирует транскрипцию генов, кодирующих α-, β-, γ-, δ-цепи Т-клеточных рецепторов, IL-3, гранзим В, CD3, ГМ-КСФ и цитокин MIP-1α, в Т-лимфоцитах [3-5]. В незрелых миелоидных клетках RUNX1 активирует гены миелопероксидазы и эластазы нейтрофилов. RUNX1 может не только активировать транскрипцию, но выступать и в качестве фактора негативной регуляции. Однако ингибитором чаще служит не полноразмерный белок RUNX1, а его укороченная изоформа RUNX1a. В то же время, этот белок в разных клеточных линиях может вести себя по-разному: активировать транскрипцию целевого гена в одних клетках и подавлять ее в других, что показано на примере регуляции гена транскрипционного фактора PU.1. В промоторной области этого гена находятся три сайта связывания RUNX1. При этом в миелоидных клетках и В-лимфоцитах RUNX1 стимулирует экспрессию гена PU.1. Нарушение регуляции работы гена PU.1 приводит к дедифференцировке предшественников миелоидных и лимфоидных клеток крови, их неконтролируемой пролиферации, что, в свою очередь, ведет к развитию лейкоза [1]. Процессы гемопоэза и ангиогенеза тесно связаны, однако на сегодняшний день мало что известно о физическом и функциональном взаимодействии транскрипционных факторов, вовлеченных в эти процессы. Получены экспериментальные данные, позволяющие предположить, что фактор RUNX1 связывает процессы гемопоэза и ангиогенеза.
Несмотря на то, что продукт гена RUNX1 контролирует экспрессию большого числа генов, о регуляции экспрессии самого гена на сегодняшний день известно немного. Установлено, что экспрессия этого гена характеризуется выраженной тканевой специфичностью. RUNX1 экспрессируется только в клетках кроветворной системы [6].
На сегодняшний день известны многочисленные повреждения гена RUNX1, в том числе точечные мутации в кодирующей области, приводящие к аминокислотным заменам, делеции и дупликации локуса q22 хромосомы 21 человека, а также хромосомные транслокации. Последние вызывают наибольший интерес, так как они чаще всего коррелируют с развитием различных лейкозов. Описано более 30 хромосомных транслокаций, затрагивающих ген RUNX1, некоторые хромосомные перестройки могут приводить к развитию различных лейкозов. Среди хромосомных перестроек с участием гена RUNX1 лучше всего изучены транслокации t(8;21) и t(3;21).
Ген TEL находится в 12 хромосоме, относится к семейству факторов транскрипции ETS. Белки-ETS контролируют экспрессию генов, которые влияют на ряд биологических процессов, включая клеточную пролиферацию, дифференциацию, трансформацию и апоптоз. Так же они участвуют и в белок - белковых взаимодействиях. TEL принадлежит к маленькому классу ETS-транскрипционных репрессоров, действия которых контролируются митогенными (вызывающими митоз) сигналами клеточного цикла.
Хромосомные перестройки с участием TEL приводят к различным миелоидным и лимфоидным лейкозам и к возникновению солидных опухолей. Продукты перестроек можно разделить на две группы: транскрипционные факторы и тиразинкиназы. TEL представляет собой ингибитор увеличения числа злокачественных клеток [7].
В результате транслокации гены «сшиваются» на месте разрыва. В гене TEL место разрыва находится между пятым и шестым интроном. В гене AML1– между вторым и третьим. Биологические функции химерного гена в лейкозе остаются неизвестными.
Список цитируемой литературы
1. , , Транскрипционный фактор RUNX1, Молекулярная биология. 2012, том 46, №6, с. 846-859.
2. , , Наноколонии и диагностика онкологических заболеваний, ассоциированных с хромосомными транслокациями, Успехи биологической химии. 2010, том 50, с. 387–446.
3. Liu H., Holm M., Xie X. Q., Wolf-Watz M., Grundstrom T. AML1/Runx1 recruits calcineurin to regulate granulocyte macrophage colony-stimulating factor by Ets1 activation. J. Biol. Chem. 20, 29398–29408.
4. Redondo J. M., Pfohl J. L., Hernandez_Munain C.,Wang S., Speck N. A., Krangel M. S. Indistinguishable nuclear factor binding to functional coresites of the T-cell receptor delta and murine leukemia virus enhancers. Mol. Cell Biol. 1992. 12, 4817–4823.
5. Bristow C. A., Shore P. Transcriptional regulation of the human MIP"1alpha promoter by RUNX1 and MOZ. Nucl. Acids Res. 2003. 31, 2735–2744.
6. Levanon D., Negreanu V., Bernstein Y., Bar-Am I.,Avivi L., Groner Y. AML1, AML2, and AML3, the human members of the runt domain gene-family: cDNA structure, expression, and chromosomal localization. Genomics. 1994. 23, 425–432.
7. Wendy A. G. Stams. Genetic and biological diversity in TEL-AML1 positive ALL. Clin Cancer Res. 2005. 11, .
2. Материалы и Методы
2.1. Материалы и реактивы:
В работе использовали следующие материалы и наборы реактивов:
2.1.1. Реактивы для полимеразной цепной реакции:
Дистиллированная вода (mQ) – бидистиллированная и деионизованная на установке Milli-Q/Milli-ρ (Millipore Corporation, USA);
Буферные растворы:
Десятикратный буферный раствор с Mg2+ для ПЦР /10 x PCR buffer / (Евроген, Россия)
Смесь нуклеотидтрифосфатов /dNTP/ 2,5 мМ каждого из четырех дезоксирибонуклеотид трифосфатов (Евроген, Россия)
Термоустойчивые ДНК-полимеразы: HS-TaqPol – 20х; TaqPol – 5ед/мкл (Евроген, Россия)
Агароза - LE 2 agarose (Хеликон, Россия)
Препараты первых цепей кДНК – препараты получены сотрудниками ЛСФГ с образцов тотальной РНК, выделяемой из клеток лейкоцитарной фракции крови больных ОЛЛ. Синтез кДНК проводят с олиго-dT затравочного праймера; содержание кДНК в использованных нами препаратах составляло 10-30 нг/мкл
Олигонуклеотидные праймеры для ПЦР синтезированы по заказу на
Таблица 1. Структуры использованных в работе олигонуклеотидных праймеров
название | Структура (5’ – 3’) | Рабочая концентрация |
TEL/AML For | GCCCATCAACCTCTCTCATCG | 10 пмоль/мкл |
TEL/AML Rev | GCCTCGCTCATCTTGCCTG | 10 пмоль/мкл |
М13-dir | GTATAGAAAGAAGTAGACATAGGAGACTC | 20 пмоль/мкл |
М13-rev | AAAAAGACACAGAGACAAAGTATAGAG | 20 пмоль/мкл |
2.1.2. Среды и растворы.
LB (на 1 л ): Триптон 10 г
Дрожжевой экстракт 5 г
NaCl 5 г
Трис-HCl (2M) 5 мл
Среду автоклавировали 1 ч при 0,5 атм.
LB-агар: перед автоклавированием в среду добавлен агар (Difco) до концентрации 1,5%.
IPTG (изопропил-тио-галактазид) – 2М водный раствор;
Х-Gal (5-Br-4-Cl-3-индолил-β-D-галактозид) – 50мг/мл раствор в диметилформамиде
2.1.3. Реактивы для гель-электрофореза:
1хТрис-ацетатный буфер (ТАЕ; 40 мМ Трис-ацетат, рН 7,8-8,0; 1 мМ ЭДТА)
Концентрированный раствор бромистого этидия (10 мг/мл)
Маркеры молекулярных длин ДНК - 1 kb или 100 DNA ladder – (Сибэнзим, Россия)
Буфер для нанесения проб (10 мМ Трис-HCL, рН 7,8; 0,025% бромфеноловый синий; 0,025% ксиленцианол; 30% глицерин; 25 мМ ЭДТА)
2.1.4. Набор реактивов и ферментов для клонирования ПЦР продуктов - pGEM-T Easy Vector System (Promega, USA):
Двухкратный буферный раствор /2X Rapid Ligation Buffer, T4 DNA Ligase/ (60mM Tris-HCl(pH 7.8); 20mM MgCl2; 20mM DTT; 2mM ATP; 10% polyethylene glycol)
Вектор / pGEM-T Easy Vector /
2.1.5. Система очистки плазмидной ДНК “Wizard Plus SV Minipreps DNA Purification System” (Promega, USA):
Cell Resuspension Solution (50mM Tris-HCl (pH 7.5); 10mM EDTA, 100μg/ml RNase A)
Cell Lysis Solution (0.2M NaOH; 1% SDS)
Neutralization Solution (4.06M guanidine hydrochloride; 0.759M potassium acetate; 2.12M glacialacetic acid)
Column Wash Solution (162/8mM potassium acetate; 22.6mM Tris-HCl (pH 7.5); 0.109mM EDTA (pH 8.0))
Minipreps Spin Columns
Collection Tubes (2ml)
Alkaline Protease Solution
Nuclease-Free Water
2.2. Методы
2.2.1. Полимеразная цепная реакция:
Для проведения полимеразной цепной реакции (ПЦР) в пробирке на 0.5 мл готовили смесь, общим объемом 20 мкл:
ДНК-матрица 10нг-50 нг
праймеры (прямой и обратный) по 10 пмоль
смесь дезоксинуклеотидтрифосфатов 2.5 мМ
буфер для Taq-полимеразы х 10 х 1
Taq-полимераза 0.5 ед.
H2O до 18 мкл
В первую очередь, в пробирку мы добавляли дистиллированную воду и буфер для ПЦР. Количество воды мы высчитывали путем вычитания из общего объема всех остальных компонентов реакционной смеси.
Затем в пробирку добавляли dNTP и смесь праймеров. Мы использовали два вида праймеров - М13 и TEL/AML. М13 праймеры мы брали для скрининга бактериалных рекомбинантных клонов. После этого все компоненты смеси хорошо размешивали. И в самую последнюю очередь в пробирку добавляли Taq-полимеразу и матричную кДНК.
В конце всю пробирку мы еще раз тщательно перемешивали пипетированием для равномерного распределения всех компонентов смеси.
Пробирка с готовой реакционной смесью помещали в амплификатор DNA Engine. У нашего амплификатора крышка так же нагревалась, поэтому мы не добавляли масла в пробирку с реакционной смесью для того, чтобы избежать испарения смеси.
Профиль цикла амплификации для TEL/AML - праймеров:
предварительный прогрев 20 сек при 95оС
денатурация ДНК 20 сек при 95оС
отжиг олигонуклеотидов 20 сек при 62оС
достройка цепей ДНК 30 сек при 72оС
Реакция включает 42 цикла амплификации (повторение стадий 2-4)
Профиль цикла амплификации для М13-праймеров:
предварительный прогрев 20 сек при 95оС
денатурация ДНК 20 сек при 95оС
отжиг олигонуклеотидов 20 сек при 56оС
достройка цепей ДНК 30 сек при 72оС
Реакция включает 42 цикла амплификации (повторение стадий 2-4)
После проведения ПЦР мы анализировали полученные продукты при помощи электрофореза.
2.2.2. Электрофорез.
Приготовление агарозного геля:
В 100 мл электрофорезного буфера мы добавляли 0,8 г порошка агарозы.
Колбу с раствором мы нагревали в микроволновой печи до тех пор, пока агароза полностью не растворялась в буфере.
Затем добавляли 4мкл раствора бромистого этидия и остужали раствор примерно до 50°C
В подложку для заливки геля с гребенками мы выливали раствор и ждали, пока застынет гель.
После того, как гель полностью затвердеет, мы удаляли гребенки и помещали гель в электрофорезную камеру(гель должен быть полностью покрыт буфером)
Продукты ПЦР мы смешивали с буфером для нанесения проб в соотношении 10:1
В первую лунку в геле при помощи автоматической пипетки мы вводили маркер, в следующие – ПЦР - продукты с буфером.
Затем электрофорезную камеру подключали к источнику питания «Эльф-4» (ДНК-Технология, Россия) мощностью 155В на 20-30 минут.
Через 20-30 минут мы аккуратно вытаскивали гель и помещали его на ультрафиолетовый детектор с CCD камерой и снимали полученные результаты.
2.2.3 Лигирование:
В 1,5мл пробирку добавляли 5мкл двукратного буфера, 0.5мкл вектора рGEM-T, 0.1мкл (3 ед. Вейса) Т4 ДНК лигазы и 3.5мкл свежего ПЦР - продукта.
Смесь перемешивали пипетированием и инкубировали 12-14 часов при 4 °С.
По окончании реакции фермент инактивировали прогревом при 65°С в течение 10 мин.
2.2.4 Трансформация:
1) В пробирку с 100мкл суспензией компетентных клеток мы добавляли 2мкл лигазной смеси, хорошо перемешивали и инкубировали полученную смесь на ледяной бане в течение 40 минут.
2) После инкубирования мы помещали пробирку в суховоздушный термостат «Термит» (ДНК Технология, Россия) и инкубировали при температуре 42°С 90 секунд.
3) Сразу же переносили пробирку в лед на 2 минуты.
4) После этого в пробирку мы добавляли 950мкл питательной среды LB и подращивали клетки в течении часа при температуре 37оС с перемешиванием.
5) Для каждой лигированной реакционной смеси с питательной средой мы подготавливали по 2 чашки петри со агаризованной средой LB, содержащей 150мкг/мл ампициллина, Среда застывала при комнатной температуре. На поверхность агара в каждую чашку Петри добавляли 10мкл IPTG и 15мкл X-Gal, равномерно распределяли по поверхности и втирали оба раствора. В каждую чашку Петри с LB-агаром вносили от 100мкл до 600мкл суспензии подращенных трансформированных клеток. Колонии выращивались в течение 16-24 часов при температуре 37оС.
2.2.5 Выделение плазмидной ДНК с использованием набора Wizard SV Minipreps:
1) 2 мл ночной культуры E. coli переносили в 1,5мл пробирку и центрифугировали 10 минут при 4000 об/мин.; супернатант удаляли
2) К клеточному осадку добавляли 250мкл Cell Resuspension Solution и полностью перемешивали вортексированием до образования однородной клеточной суспензии.
3) Затем добавляли к клеточной суспензии 250мкл Cell Lysis Solution и перемешивали, переворачивая пробирку 4 раза до тех пор, пока содержимое пробирки не посветлело. Инкубировали (примерно 1-5 минут).
4) Добавляли 350мкл Neutralization Solution и сразу же перемешивали инвертированием.
5) Центрифугировали бактериальный лизат на максимальной скорости (10-12 тыс. об/мин) в течение 10 минут при комнатной температуре.
6) Далее мы аккуратно наносили 800мкл очищенного лизата на Spin Column, поставленную в пустую 2,0мл пробирку (Collection Tube), для связывания плазмидной ДНК с носителем.
7) Затем мы центрифугировали всю систему на максимальной скорости в течение 1 минуты при комнатной температуре. После этого вытаскивали Spine Column из Collection Tube, фильтрат выливали из Collection Tube и вставляли Spine Column в Collection Tube.
8) Добавляли 750мкл Column Wash Solution, предварительно разбавив его 95% этанолом, в Spine Column.
9) Центрифугировали на максимальной скорости в течение 1 минуты при комнатной температуре. После этого вытаскивали Spine Column из Collection Tube, фильтрат выливали из Collection Tube и вставляли Spine Column в Collection Tube.
10) Повторяли промывку, используя 250мкл Column Wash Solution.
11) Переносили Spine Column в чистую 1.5 миллилитровую пробирку.
12) Элюировали ДНК плазмиды, добавляя 100мкл Nuclease-Free Water в Spine Column. Центрифугировали на максимальной скорости в течение 1 минуты при комнатной температуре.
13) После этого убирали Spine Column из пробирки и выбрасывали, оставляли только пробирку с выделенной ДНК. Пробирку закрывали и хранили при температуре -20оС.
2.2.6 Приготовление ночной культуры трансформированных клонов
1) Брали небольшую часть выбранной единичной колонии зубочисткой.
2) Переносили зубочистку в стеклянную пробирку с 2 мл жидкой LB-среды.
3) Выращивали бактериальный клон в течение 12-14 часов при 37°С на термостатируемой качалке.
2.2.7. Получения ДНК для скрининга рекомбинантных клонов
1) Небольшую часть выбранной единичной колонии переносили зубочисткой в пробирку объемом 1,5 мл с 10 мкл дистиллированной воды.
2) Перенесенные бактериальные клетки разрушали нагреванием в кипящей водяной бане в течение 5-6 минут.
3) Полученный раствор использовали в качестве матрицы для ПЦР при скрининге рекомбинантных клонов.
2.2.8. Секвенирование ДНК
Очищенную ДНК рекомбинантных плазмид отдавали в сервис для определения нуклеотидных последовательностей плазмидных вставок на автоматическом секвенаторе ABI 373 (Apply Biosystems)
3. Результаты
3.1 Проведение ПЦР-анализа наличия транслокаций t(12;21)
Транслокация t(12;21) характерна для больных В-клеточным острым лимфобластным лейкозом (В-ОЛЛ). В результате транслокации образуется химерный, но транскрипционно активный ген TEL/AML1.
Так как ставить ПЦР-анализ с геномной ДНК очень трудно из-за большой длины интронов, мы проводили анализы с кДНК лимфоцитов, полученных от больных В-ОЛЛ (кДНК – одноцепочечная копия зрелой мРНК).
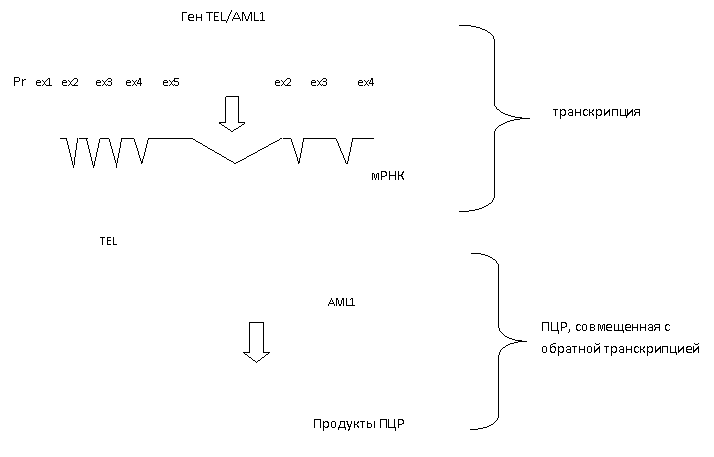
Нами была проведена серия ПЦР-реакций. С помощью них мы выявляли наличие транскрипта химерного гена, образовавшегося в результате транслокации. Один из праймеров идентичен участку, находящегося на конце 5 экзона гена TEL. Второй праймер комплементарен участку 3 экзона гена AML1 (см. рис. 1).
Положительный результат ПЦР-реакций свидетельствует об образовании мРНК TEL/AML.
Рисунок 3. Общая стратегия ПЦР-анализа
![]()
![]()
Pr – промоторный участок гена.
Сплошной черной линией обозначена геномная ДНК. Синие, белые и голубые прямоугольники обозначают экзоны.
ex1, ex2 и т. д – номера экзонов.

![]()


 - первичный транскрипт.
- первичный транскрипт.
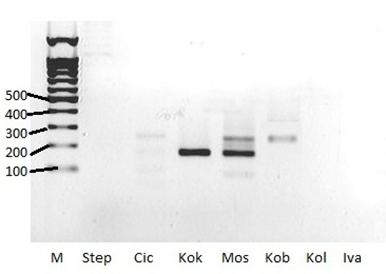
Рисунок 4. Результаты ПЦР-анализа
![]()
![]()
![]()
![]()
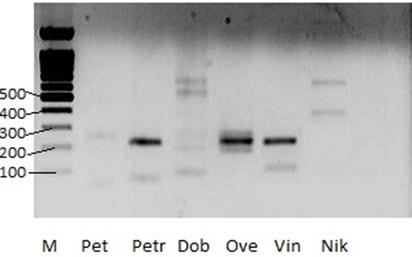
М-маркер длин 1bp (СибЭнзим, Россия).
Длины маркерных фрагментов (bp) указаны слева.
Ярко-черные полосы в центре рисунка – ПЦР-продукты.
![]() - фигурными стречками указаны положения ПЦР-продуктов.
- фигурными стречками указаны положения ПЦР-продуктов.
Условные обозначения образцов кДНК приведены под каждой дорожкой.
На рисунке 2 показаны результаты ПЦР - анализа. В образцах Step, Cic, Kol, Iva, Pet, Nik и Dob ПЦР - продукт не был обнаружен (на их дорожках нет характерных черных полос). А на дорожках образцов Kok, Kob, Vin, Ove и Petr есть черные полосы, находящиеся примерно все на одной высоте, в районе 200 пар нуклеотидов. Но полосы Kob, Petr, Ove, Vin (210 пар нуклеотидов) находятся выше фрагмента «200 п. о.» маркера длин, Kok – ниже (171 пара нуклеотидов), а образец Mos имеет 2 полосы (210 и 171 пар нуклеотидов). Это значит, что длина ПЦР - продуктов у одних образцов одна, а у других – другая, что говорит о разных местах разрыва при слиянии фрагментов генов TEL и AML1 в химерный ген или разных вариантов сплайсинга транскриптов этого гена.
Наши результаты совпадают с результатами цитогенетического анализа, проведенного в клинике и установившего эту транслокацию t(12;21) у пациентов Kok, Mos, Kob, Vin, Ove и Petr.
3.2 Клонирование фрагментов кДНК TEL/AML1
Мы провели клонирование фрагментов кДНК TEL/AML1 для того, чтобы определить нуклеотидную последовательность этого гена. Длина ПЦР - продукта (171 или 210 пар нуклеотидов) может зависеть не только от точки разрыва, но и от типа сплайсинга.
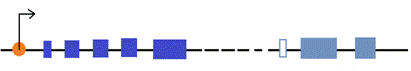
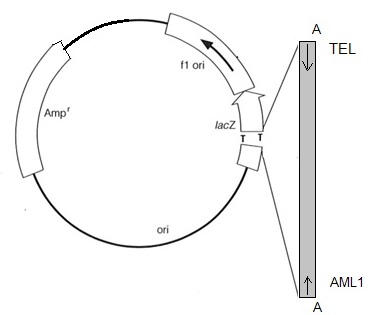
Для клонирования ПЦР - продуктов мы использовали специальную векторную систему pGEM - T (рис. 5). Так как ПЦР - продукты содержат на концах добавочные аденины (А на рис. 5), то для повышения эффективности их встраивания на концы векторной плазмиды введены комплементарные тимидины (Т на рис. 5). ПЦР - продукты мы внедряли в плазмиду pGEM-T (процедура «лигирование» в разделе Методы), а затем рекомбинантные плазмиды мы помещали в клетки E. coli (процедура «трансформация» в разделе Методы).
В ДНК плазмиды есть ген Ampr. Он кодирует ампицилазу, которая расщепляет ампицилин в среде, что не позволяет вырасти колониям бактерий без плазмид.
Так же в ДНК плазмиды находится ген lacZ, котрый кодирует β-галактозидазу, расщепляющую хромогенный субстрат XGal. Расщепление XGal приводит к синему окрашиванию колоний. Если же в бактерию попала плазмида со вставкой, то структура гена lacZ нарушается, в следствие чего β-галактозидаза не синтезируется, и колония не имеет цвета. На рис. 6 приведен внешний вид нескольких чашек Петри в выросшими колониями рекомбинантных клонов.
Рисунок 5. Схема клонирования
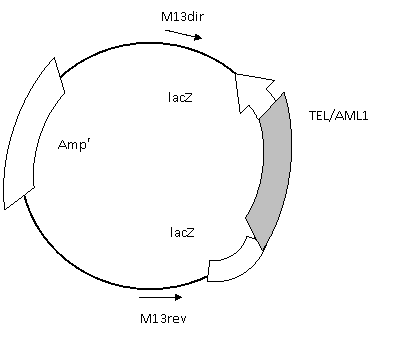 |
Аmpr – ген устойчивости к ампецилину.
lacZ – ген β-галактозидазы.
Ori – точка начала репликации в бактериальной системе E.coli
f1 ori – точка начала репликации бактериофага f1
M13dir – прямой прамер.
M13rev – обратные праймер.
Рисунок 6. Чашки Петри с колониями рекомбинантных клонов
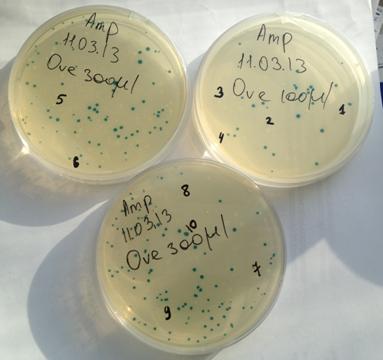
1-8 – белые колонии
9,10 - сини колонии
Для скрининга ДНК рекомбинантных клонов мы использовали праймеры М13. Они коплементарны участкам ДНК плазмидного вектора, расположенным вблизи от сайта клонирования вставки. Поэтому, если в исходной плазмиде (векторе) нет вставки, то в результате ПЦР будет образовываться продукт длиной 220 п. о. (участок плазмидной ДНК). Если же в ДНК есть вставка, то длина продукта равно 220 п. о. + 171/210 п. о. (зависит от длины вставки). На рис. 7 в качестве примера представлены результаты ПЦР-скрининга четырех независимых клонов.
Рисунок 7. Скрининг
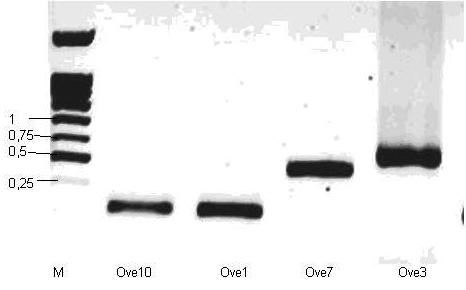
М – маркер длин 1Kb(СибЭнзим, Россия).
Ove10-синяя колония без вставки
Ove1- белая колония без вставки
Ove3 – белая колония со вставкой
Ove7 – белая колония
3.3 Проведение анализа первичных структур кДНК TEL/AML1.
Очищенную ДНК рекомбинантных плазмид отдавали в сервис для определения нуклеотидных последовательностей плазмидных вставок на автоматическом секвенаторе ABI 373 (Apply Biosystems).
С полученными при секвенировании нуклеотидными последовательностями мы проводили множественные выравнивания (программа BioEdit). Мы выяснили, что у всех пациентов одинаковые нуклеотидные последовательности транскрипта химерного гена TEL/AML1, кроме пациента Mos. У него мы обнаружили два разных типа кДНК химерного гена. Один включает в себя второй экзон гена AML1, а другой не имеет этого экзона (рис.1). Результаты секвенирования фрагмента кДНК из лимфобластов пациента Mos подтверждают предположение о наличии двух типов транскриптов TEL/AML1, сделанное нами по результатам ПЦР - анализа (рис.4).
Рисунок 8. Результаты секвенирования
pGEM TELfr TEL exon5
 160
160
TEL/AML GCCCATCAACCTCTCTCATCGGGAAGACCTGGCTTACATGAACCACATCATGGTCTCTGTCTCCCCGCCTGAAGAGCACGCCATGCCCATTGGGAGAATA
MOS-1 ggatt GCCCATCAACCTCTCTCATCGGGAAGACCTGGCTTACATGAACCACATCATGGTCTCTGTCTCCCCGCCTGAAGAGCACGCCATGCCCATTGGGAGAATA
MOS10 ggattGCCCATCAACCTCTCTCATCGGGAAGACCTGGCTTACATGAACCACATCATGGTCTCTGTCTCCCCGCCTGAAGAGCACGCCATGCCCATTGGGAGAATA
Kob-1 ggattGCCCATCAACCTCTCTCATCGGGAAGACCTGGCTTACATGAACCACATCATGGTCTCTGTCTCCCCGCCTGAAGAGCACGCCATGCCCATTGGGAGAATA
Kok-2 ggattGCCCATCAACCTCTCTCATCGGGAAGACCTGGCTTACATGAACCACATCATGGTCTCTGTCTCCCCGCCTGAAGAGCACGCCATGCCCATTGGGAGAATA
Ove-3 ggattGCCCATCAACCTCTCTCATCGGGAAGACCTGGCTTACATGAACCACATCATGGTCTCTGTCTCCCCGCCTGAAGAGCACGCCATGCCCATTGGGAGAATA

AML exon2 AML exon 3 AMLrev
TEL/AML GCAGAATGCATACTTGGAATGAATCCTTCTAGAGACGTCCACGATGCCAGCACGAGCCGCCGCTTCACGCCGCCTTCCACCGCGCTGAGCCCAGGCAAGAT
MOS-1 GCAG --ATGCCAGCACGAGCCGCCGCTTCACGCCGCCTTCCACCGCGCTGAGCCCAGGCAAGAT
MOS10 GCAGAATGCATACTTGGAATGAATCCTTCTAGAGACGTCCACGATGCCAGCACGAGCCGCCGCTTCACGCCGCCTTCCACCGCGCTGAGCCCAGGCAAGAT
Kob-1 GCAGAATGCATACTTGGAATGAATCCTTCTAGAGACGTCCACGATGCCAGCACGAGCCGCCGCTTCACGCCGCCTTCCACCGCGCTGAGCCCAGGCAAGAT
Kok-2 GCAGAATGCATACTTGGAATGAATCCTTCTAGAGACGTCCACGATGCCAGCACGAGCCGCCGCTTCACGCCGCCTTCCACCGCGCTGAGCCCAGGCAAGAT
Ove-3 GCAGAATGCATACTTGGAATGAATCCTTCTAGAGACGTCCACGATGCCAGCACGAGCCGCCGCTTCACGCCGCCTTCCACCGCGCTGAGCCCAGGCAAGAT
pGEM
 210
210
TEL/AML GAGCGAGGC
MOS-1 GAGCGAGGCaatcacta
MOS10 GAGCGAGGCaatcacta
Kob-1 GAGCGAGGCaatcacta
Kok-2 GAGCGAGGCaatcacta
Ove-3 GAGCGAGGCaatcacta
Зеленым шрифтом обозначена нуклеотидная последоватедьность праймера «TELfr», шрифтом синего цвета – последовательность праймера AMLrev, строчными буквами – фрагменты нуклеотидной последовательности плазмиды pGEM-T
Вертикальными линиями отмечены границы между экзонами, нуклеотидные последовательности 2-го экзона гена AML1 выделена прямоугольником
В результате сравнительного анализа нуклеотидных последовательностей ПЦР - фрагментов мы подтвердили наличие двух типов транскриптов химерного гена, установили экзонную структуру обоих типов транскриптов и показали отсутствие индивидуальных мутаций в исследуемом фрагменте химерного гена TEL/AML1 в лимфобластах 4-х неродственных пациентов с ОЛЛ.
3.4 Заключение.
Мы провели ПЦР-анализы наличия транслокаций t(12;21)(p13;q22) в образцах лимфобластных клеток пациентов, больных острым лимфобластным лейкозом, затем клонировали фрагменты транскриптов в E. coli и секвенировали несколько рекомбинантных клонов - Kok, Mos, Kob, Ove (табл. 2). По результатам секвенирование мы определили нуклеотидную последовательность транскрипта химерного гена. У пациентов Kok, Kob и Ove на стыке пятого экзона гена TEL и третьего экзона гена AML1, находится второй экзон гена AML1. У пациента Mos мы наблюдали два варианта транскрипта химерного гена. В одном случае в химерном гене присутствует второй экзон гена AML1, а в другом отсутствует.
Таблица 2.
Пациенты | Наличие t(12;21)* | Структура кДНК TEL/AML1 | |
TELex5/ex2-ex3AML1 | TELex5/ex3AML1 | ||
Step | - | Не определяли | |
Cic | - | Не определяли | |
Kok | + | + | - |
Mos | + | + | + |
Kob | + | + | - |
Kol | - | Не определяли | |
Iva | - | Не определяли | |
Pet | - | Не определяли | |
Petr | + | Не определяли | |
Dob | - | Не определяли | |
Ove | + | + | - |
Vin | + | Не определяли | |
Nik | - | Не определяли |
![]() * - по результатам ПЦР-анализа образцов кДНК
* - по результатам ПЦР-анализа образцов кДНК
«+» - во второй колонке обозначает наличие транслокации; в третьей колонке обозначает наличие определенной структуры кДНК TEL/AML1.
«- » - во второй колонке обозначает отсутствие транслокации; в третьей колонке обозначает отсутствие определенной структуры кДНК TEL/AML1.
ВЫВОДЫ
1. По результатам проведенного ПЦР-анализа панели образцов кДНК из клеток лейкоцитарной фракции крови больных ОЛЛ подтверждено наличие транслокации t(12;21)(p13;q22) у 6 из 13 обследованных пациентов. Доказана транскрипционная активность химерного гена TEL/AML1 в шести образцах опухолевых лимфобластных клеток.
2. Выполнено клонирование и секвенирование фрагментов кДНК химерного гена TEL/AML1 четырех пациентов.
3. Проведен сравнительный анализ нуклеотидных последовательностей фрагментов кДНК. Показана идентичная структура транскриптов гена TEL/AML1 у трех пациентов (образцы Kok, Kob и Ove). Установлено одновременное образование двух изоформ транскрипта гена TEL/AML1, отличающихся по наличию/отсутствию 2-го экзона AML1, у одного из обследованных пациентов (образец Mos).
Благодарности
Мы выражаем благодарность за организацию практики и за предоставление возможности проведении работы в лаборатории сравнительной и функциональной геномики ИБХ РАН, за чуткое руководство и огромную проделанную с нами работу.
Мы так же благодарим за предоставленные образцы кДНК для работы и Е. Богачеву и за советы и помощь в проделывании опытов.
(И рецензента )
