

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Глава 4. Природная динамика сообществ мышевидных грызунов в лесных экосистемах Каспийско-Балтийского водораздела
Одним из основных подходов изучения структуры сообществ является концепция гильдии - группы видов, использующих совокупность ресурсов сходным образом (Джиллер, 1988). В данной главе рассматриваются особенности структуры и динамики гильдий мышевидных грызунов в коренных ельниках разного генезиса, и обсуждаются их реакции на ветровальные разрушения зрелых древостоев.
Динамика сообществ мышевидных грызунов в коренных ельниках. В неморальных ельниках зарегистрировано 13 видов мышевидных грызунов. Основа сообщества сформирована одним доминантным видом (рыжая полевка - 96,2%), четырьмя обычными и постоянно встречающимися с невысокой долей участия в составе гильдий: красная (1,2%), темная (0,7%) и подземная (0,4%) полевки, малая лесная мышь (0,9%). Остальные виды регистрируются крайне нерегулярно, в единичных экземплярах, и могут быть отнесены к категориям редких и случайных. В сообществах бореальных ельников отмечено 7 видов мышевидных грызунов. Наблюдается содоминирование рыжей (74,6%) и красной (20,5%) полевок. К числу обычных и регулярно присутствующих в отловах с относительно невысокой долей видов принадлежит темная полевка (2,7%). «Краевой эффект» за счет проникновения чужеродных видов в коренных ельниках выражен чрезвычайно слабо, поэтому сообщества грызунов удаленных и близко расположенных к антропогенным территориям участков лесного массива не имеют достоверных различий.
За рассматриваемый период времени в сообществах коренных ельников на фоне циклических колебаний наблюдались четко выраженные тренды увеличения суммарного обилия грызунов (рис. 1).
|
Рис.1. Динамика суммарной численности сообществ мышевидных грызунов в коренных ельниках ЦЛГПБЗ (R2 – величина достоверности аппроксимации) |
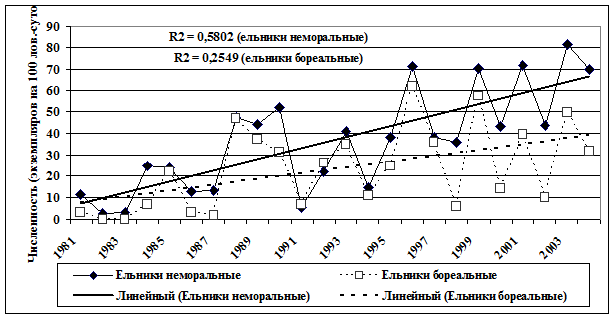
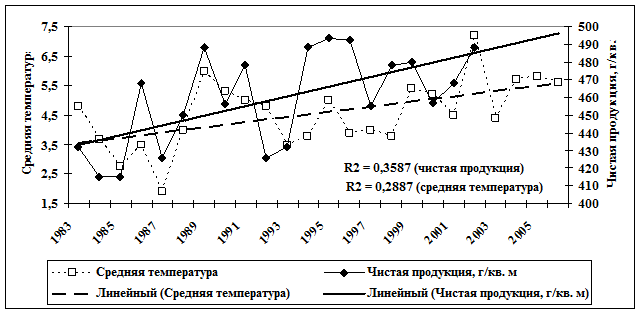
В большей степени это характерно для неморальных ельников и связано с характером динамики фоновых видов. В целом рост численности гильдий грызунов хорошо согласуется с общим ходом динамики среднегодовой температуры и чистой продукции в коренных лесных экосистемах (рис. 2).
|
Рис. 2. Динамика среднегодовой температуры и чистой продукции в естественных лесных экосистемах (R2 – величина достоверности аппроксимации) |
С 1992 г. в коренных зеленомошных ельниках происходят процессы «бореализации» сообществ грызунов (Истомин, 1998; 2005). Доля участия восточно-палеарктических видов в составе гильдий по сравнению с периодом гг. возросла почти в 10 раз (рис. 3).
| Рис. 3. Доля особей восточно-палеарктических видов в составе гильдий грызунов в разные периоды исследований в коренных ельниках |
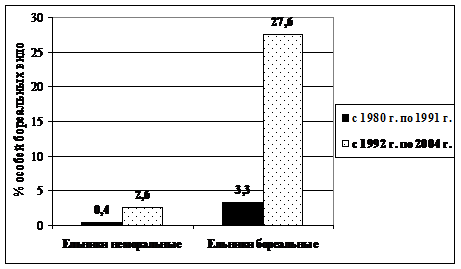
Прежде всего, это произошло за счет увеличения численности красной полевки. Структура гильдий грызунов бореальных еловых лесов в сравниваемые периоды времени отличается весьма существенным образом (IChS=0,28, Р=0,999). Аналогичные процессы отмечены и для гильдий землероек (Истомин, 1998). В неморальных ельниках после 1994 г. красная полевка также стала довольно регулярно встречаться в отловах почти на всех пробных площадях. Кроме этого, наблюдалось увеличение численности западно-палеарктических видов, находящихся здесь почти на краю ареала: подземной полевки, малой лесной мыши. В значительной степени возрастание роли этих типичных неморальных видов, ранее очень редких, может быть обусловлено потеплением зимних месяцев и ветровальными процессами, которые вызывают формирование в древостое «окон» различного размера.
Формирование и динамика разнообразия сообществ до сих пор принадлежат к числу наиболее дискуссионных вопросов. В исследованный нами промежуток времени разнообразие гильдий грызунов демонстрировало устойчивую тенденцию роста в неморальных и бореальных ельниках (рис. 4).
|
|
Рис. 4. Некоторые показатели разнообразия и летняя суммарная численность сообществ мышевидных грызунов в коренных неморальных (слева) и бореальных (справа) ельниках в разные периоды исследований |
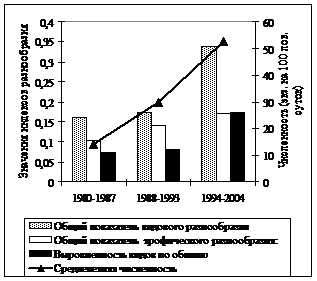
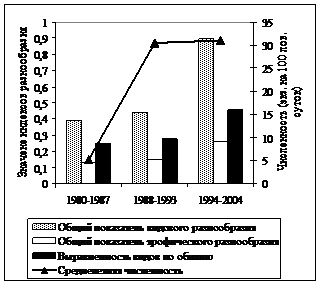
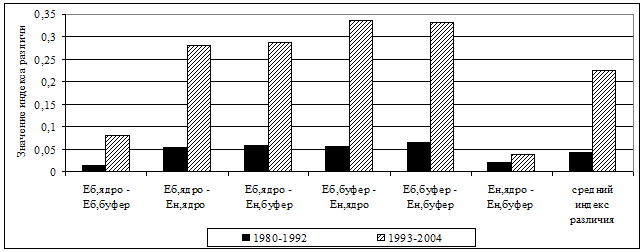
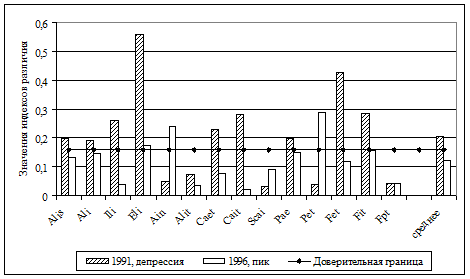
В первую очередь, это связано с более выровненным распределением по обилию типичных лесных видов. Считается, что одним из важных факторов формирования разнообразия сообществ животных является пространственная гетерогенность среды (Уиттекер, 1980; Пианка, 1981; Истомин, 1984; 1992; Одум, 1986; Джиллер, 1988; Бигон и др., 1989; Лукьянова, Лукьянов, 2004 и др.). В нашем случае повышение разнообразия сообществ, по-видимому, также во многом определяется увеличением структурной сложности среды в результате ветровальных процессов, усыханий еловых древостоев с их последующим разрушением, что привело к формированию чрезвычайно мозаичного почвенно-растительного покрова. Отмеченные тенденции обуславливают выраженную в последние годы гетероморфность сообществ грызунов. Тогда как в 70-80-е гг. прошлого века группировки мелких млекопитающих были гораздо более однородными на всей исследованной территории. Значения индексов различия между сравниваемыми типами местообитаний в период гг. в среднем в 5 раз выше, чем в гг. (рис. 5). Это свидетельствует также об увеличении бета-разнообразия сообществ мышевидных грызунов в лесном массиве в целом.
|
Рис. 5. Степень различия структуры сообществ мышевидных грызунов в коренных ельниках ЦЛГПБЗ в различные периоды наблюдений. Еб - ельники бореальные; Ен – ельники неморальные; ядро – участки, удаленные от антропогенных территорий; буфер – участки с удалением не более 1 км от антропогенных территорий. |
Таким образом, в последние 10-15 лет произошли значимые изменения структуры, разнообразия и обилия сообществ мелких млекопитающих в естественных лесах Каспийско-Балтийского водораздела, что привело к увеличению зональной контрастности населения мелких млекопитающих в коренных ельниках различного генезиса.
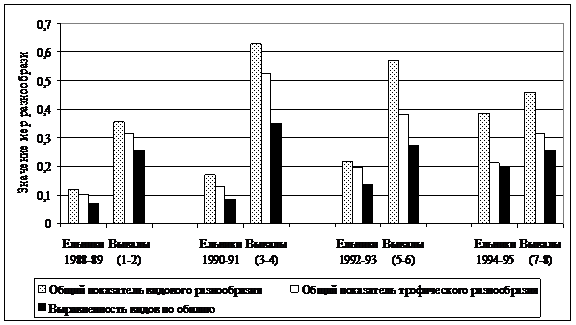
Влияние массовых ветровалов на сообщества мышевидных грызунов. Ветровалы характерны для всей лесной зоны и принадлежат к числу мощных факторов, вызывающих экологическую дестабилизацию среды. Работ, посвященных изучению воздействия ветровалов на популяции и сообщества животных, в том числе мелких млекопитающих, сравнительно немного (Истомин, 1992, 1995; Зюсько и др., 2001; Кузнецова, 2003; Лукьянова и др., 2004). В августе 1987 г. на исследуемой территории произошел массовый вывал древостоя, в результате которого 66% лесной площади оказались поврежденными, 25% подверглись сильной степени разрушения. Воздействие ветровалов на сообщества грызунов осуществляется через изменение экологической структурированности среды и ее ресурсных возможностей. Реакции мышевидных грызунов на дестабилизацию среды, вызванную ветровалами, видоспецифичны и определяются в большей степени экологическими особенностями и минимальными требованиями животных к условиям среды. На ранних стадиях демутации ветровалов снижается степень доминирования рыжей полевки и повсеместно возрастает роль полевки-экономки, которая активно заселяет участки крупных вывалов уже в первый год после ветровального воздействия. В зависимости от конкретного сочетания различных факторов на некоторых свежих вывалах довольно многочисленные популяционные группировки могут образовывать такие виды, как малая лесная мышь, темная и подземная полевки. Наличие на ветровальных площадях с преобладанием неморальной структуры отдельных парцелл бореального характера может приводить к заметному увеличению роли в составе сообщества вывалов красной полевки. Несмотря на весьма ограниченный пул лесных видов грызунов, вариантов формирования эфемерных послеветровальных сообществ с их участием может быть достаточно много, но всегда при сохранении сильного доминирования рыжей полевки. В целом на вывалах значительно возрастают выравненность видов по обилию и общие показатели видового и трофического разнообразия (Истомин, 1992, 2008). Наибольшие показатели видового разнообразия и самую высокую специфичность имеют сообщества на 3-4 годы сукцессии (рис. 6). В целом, режимы ветровальных нарушений, характер распределения и размеры элементов мозаичной структуры ранних сукцессионных стадий играют очень значимую роль в поддержании видового, структурного и генетического разнообразия сообществ мышевидных грызунов в южно-таежных лесах.
|
Рис. 6. Показатели разнообразия сообществ грызунов в коренных ельниках и на разных стадиях послеветровальной демутации (в скобках указаны годы, прошедшие после воздействия ветровала) |
Пространственная структура, динамика популяций и особенности микроэволюционных процессов второстепенных видов самым тесным образом связаны с существованием стадии "окон". Полученные результаты хорошо согласуются с современной лесной Gap-парадигмой, полагающей, что естественные леса представляют собой сукцессионную мозаику пятен, которые находятся на разных стадиях развития и обеспечивают их биоразнообразие и устойчивое существование (Коротков, 1991; Восточноевропейские…, 1994; Смирнова и др., 1999 и др.).
Глава 5. Влияние сплошных рубок и восстановительных сукцессий на сообщества мышевидных грызунов
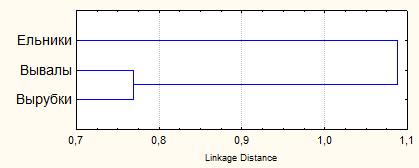
Основная форма антропогенных нарушений лесов региона – сплошные рубки, которые приводят к формированию особой структуры ландшафтов с разновозрастными серийными экосистемами. Изучали влияние рубок и восстановительных сукцессий на сообщества грызунов. Исследованные смены фитоценозов в большей степени соответствуют демутациям (Работнов, 1992; Богданова и др., 2007). В сукцессионной серии всего зарегистрировано 13 видов грызунов. Однако общее число и доля отдельных видов в составе сообществ неодинаковы на различных стадиях сукцессии. Наибольшей специфичностью обладают гильдии грызунов травянистой стадии возобновления 3-7 лет (рис. 7).
| Рис. 7. Дендрограмма сходства видовой структуры сообществ мышевидных грызунов различных стадий послерубочной демутации неморальных ельников |
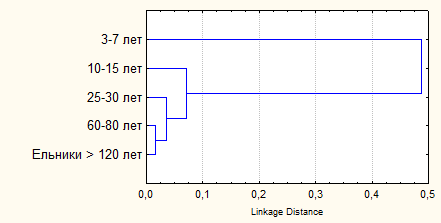
Мера различия в последовательном ряду постепенно снижается и между производными 60-80 лет и собственно неморальными ельниками становится недостоверной (Истомин, 1987, 1992). В это время сообщества ельников, как правило, вступают в устойчивую фазу развития и их принято считать «условно-коренными» (Маслов, 2004). Сообщества грызунов на ранних стадиях зарастания вырубок характеризуются высоким видовым разнообразием, что определяется большей выравненностью распределения видов по обилию. Основными факторами увеличения разнообразия являются: усиление гетерогенности среды и равномерности распределения отдельных «пятен», за счет чего формируется отчетливая сегрегация доминирующих видов; ослабление конкурентной способности рыжей полевки, как типично лесного вида, поэтому в первые годы демутации довольно часто наблюдается активная смена доминирования между тремя видами полевок (рыжей, подземной и экономкой); разделение фоновых видов по временному компоненту ниши (речь идет об асинхронном достижении сезонного максимума численности); действие хищничества, как фактора «разреживания», в результате чего снижается обилие более доступных видов-доминантов.
Таким образом, исследованные формы дестабилизации лесных экосистем (вывалы, вырубки) приводят на ранних стадиях демутации к увеличению разнообразия сообществ грызунов. Полученные результаты вполне соответствуют представлениям о «неравновесных» экосистемах, претерпевающих периодические возмущения, которые характеризуются большим разнообразием, по сравнению с «равновесными», где сильнее выражено доминирование и конкурентное исключение (Abugov, 1982). Аналогичные закономерности отмечены для сообществ мелких млекопитающих при умеренном воздействии некоторых других форм нарушений (Фомин, 1981; Гашев, 2000; Зюсько и др., 2001; Лукьянова, Лукьянов, 2004; Жигарев, 2006).
Глава 6. Природно-очаговые инфекции в коренных и производных лесах Каспийско-Балтийского водораздела
Важными интегральными характеристиками экосистем являются эпизоотические процессы, которые представляют собой постоянное взаимодействие популяций возбудителя и естественных носителей с внешней средой. В данной главе приведены результаты комплексного изучения основных природно-очаговых инфекций региона. Установлена ландшафтно-экологическая приуроченность очагов, выявлен видовой состав носителей, исследованы особенности динамики и построены модели эпизоотий, дана оценка эпидемической ситуации в регионе. Обнаружено, что в лесных экосистемах по сравнению с преобразованными территориями, существуют более устойчивые и активные очаги лептоспирозов (главным образом серогруппы Grippothyphosa) и ГЛПС (рис. 8).
| Рис. 8. Суммарная доля (в %) пораженных лептоспирозами и ГЛПС зверьков в лесных ландшафтах и агролесоландшафтах региона. |
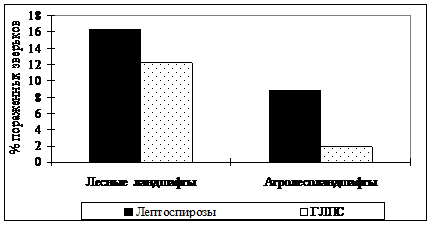
Эпизоотические потенциалы лесных очагов тесным образом связаны с особенностями динамики численности популяций рыжей полевки, которая является основным носителем данных инфекций (Карулин и др. 1993; Истомин и др., 1999). Об этом свидетельствуют высокая численность и степень инфицированности зверьков. Средняя доля особей рыжей полевки с антителами к лептоспирам - 17,5%. Более 80% инфицированных особей имели антитела в титрах 1:100 и выше, 30% - 1:1600 и выше. Наибольшее значение в сохранении возбудителей имеют неморальные ельники и осинники старших возрастных групп. Численность инфицированных особей рыжей полевки в среднем здесь составляла 6,0 экз. на 100 ловушко-суток. Развитие эпизоотий в сезонном аспекте происходит по типу нарастания процесса с быстрой эстафетной передачей возбудителя и резкого увеличения численности вирусоносителей на территории очага к концу лета. При подъеме численности рыжей полевки эпизоотии лептоспироза широко разливаются по территории и в них вовлекаются другие виды. Средняя доля зверьков с лептоспирами у темной и красной полевок, малой лесной мыши, полевки-экономки составила соответственно 27,5%, 17,9%, 14,8%, 10,1%. Специфические и активные очаги лептоспирозов с быстрой сменой основных носителей инфекции формируются на ранних стадиях сплошных вырубок. Антиген к вирусу ГЛПС в лесных экосистемах и на вырубках, кроме рыжих полевок, был обнаружен у красной полевки и экономки.
Глава 7. популяционная динамика ценозообразующих видов полевок в различных экологических условиях
Исследованы особенности динамики численности и фенетического разнообразия популяций ценозообразующих видов с разными пространственными структурами и стратегиями в условиях европейской южной тайги (рыжей и красной полевок, полевки-экономки). Выявлены тренды, периодические высокочастотные и низкочастотные колебания многолетних временных рядов численности модельных видов. Произведен расчет их примерного вклада в общее варьирование численности и построены статистические модели. Описаны неметрические вариации признаков и составлены региональные каталоги фенов. Определены наиболее полиморфные признаки, которые могут иметь индикационную значимость при изучении динамики фенетического разнообразия. Выявлены пространственно-биотопические, временные и внутрипопуляционные особенности формирования фенетической структуры и разнообразия модельных видов в различных экологических условиях.
Популяционная динамика рыжей полевки в естественных и нарушенных лесных экосистемах. Основной модельный вид на онтогенетическом и популяционном уровне исследований - рыжая полевка, которая в лесных массивах формирует континуальные популяции.
Динамика численности. В период гг. динамика численности рыжей полевки в коренных ельниках представляла собой нестационарный процесс (рис. 9).
|
Рис.9. Многолетняя динамика и тренды численности рыжей полевки в неморальных ельниках, на участках массовых ветровалов и на сплошных вырубках |
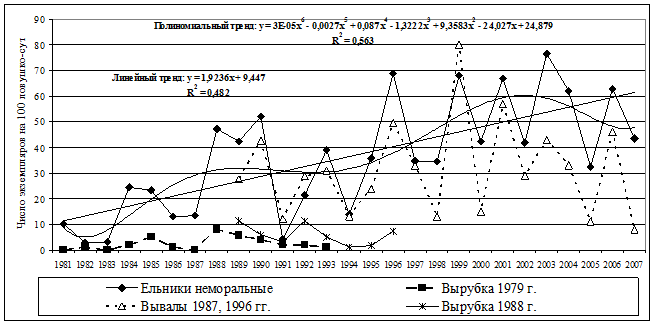
Существуют положительные тренды, на долю которых приходится более 50% варьирования исходных рядов. Достаточно реалистично эти зависимости описываются полиномиальными моделями шестой степени. На фоне увеличения численности наблюдаются периодические ее колебания. Отмечено явное упрощение структуры циклов за данный промежуток времени: от 4-летних с типичным S-образным ростом к трехлетним, а затем к простым двухгодичным. Переходы к новым режимам популяционной динамики произошли после катастрофических ветровалов 1987 и 1996 гг. Одним из триггерных механизмов смены популяционных циклов можно считать подснежное размножение, которое было зарегистрировано зимой гг., вероятно, как ответная реакция на значительное и резкое нарушение стационарности экосистем в результате широкомасштабного катастрофического ветровала. Установлено также, что конкретные сочетания климатических характеристик могут оказывать заметное воздействие на уровень численности в различные фазы популяционных циклов (Минаева, Истомин и др., 2001; Истомин, 2005, 2007). В крупном лесном массиве многолетние и сезонные изменения численности в разных типах коренных и производных местообитаний тесно сопряжены (r=0,74). Существенные перестройки общего и сезонного хода динамики численности рыжей полевки происходят на ранних стадиях возобновления вырубок. Средние показатели летней численности вида гораздо ниже (в 6-10 раз), синхронность изменений выражена слабо. В ненарушенных ельниках сезонный пик достигается в середине-конце лета, а на ранних стадиях вырубок – осенью. Это, вероятно, связано с существенным вкладом процесса расселения в динамику численности данного вида на вырубках и притоком особей из лесных экосистем, окружающих вырубленные площади (Истомин, 1999).
Динамика фенотипического разнообразия. Для 40 исследованных элементарных признаков черепа рыжей полевки выявлено 162 фена; для параконида и талонуса - 13 комплексных морфотипов.
Важнейшая характеристика популяций – ее пространственная структура, от которой в значительной степени зависят характер и скорость микроэволюционных событий. Основные представления о типах популяционной структуры и соответствующие теоретические модели были разработаны достаточно давно (Wright, 1951, 1969; Kimura, 1953; Kimura, Weiss, 1964; Malecot, 1967, 1975 и др.). Однако характер эволюционных преобразований в больших непрерывных и в совокупности мелких дискретных популяций до сих пор принадлежит к числу мало изученных вопросов. При исследовании континуальной популяции рыжей полевки нами обнаружено, что в годы высокой численности в диапазоне удаленностей до 4 км пространственные группировки не являются строго изолированными расстоянием. Корреляция между величиной фенетической дистанции и расстоянием по комплексу признаков была недостоверной (r=0,20). Число фенов на отдельных участках варьировало незначительно, доминирующие морфотипы в основном также были одинаковы. Несколько больший уровень различий получен между группировками, населяющими разные типы местообитаний, вне зависимости от расстояния между ними. Наибольшей специфичностью отличались бореальные ельники. Фенетические дистанции между группировками заметно возрастали в годы низкой численности, что вероятно, в первую очередь определяется меньшей интенсивностью генетического обмена между ними в эти периоды (рис. 10).
| Рис. 10. Фенетические дистанции между популяционными группировками рыжей полевки из неморальных и бореальных ельников в годы с низкой и высокой численностью |
Сравнивали популяционно-феногенетические особенности рыжей полевки в крупном сплошном лесном массиве и в системе мелких лесных резерватов, изолированных верховыми болотами. В биотопическом плане, с позиций единовременности и пространственной разобщенности расстоянием субпопуляции «болотных изолятов» и территориальные группировки непрерывной популяции находились в относительно сходных условиях. Однако своеобразие сравниваемых выборок из континуальной и дискретной популяций достаточно велико: из 32 признаков по 18 получены статистически значимые отличия. Обнаружено, что в дискретной популяции заметно выше общий уровень фенотипического разнообразия, но ниже доля редких фенов. Для совокупности популяций «болотных изолятов» отмечено достоверное увеличение флуктуирующей асимметрии для качественных и количественных признаков. Это может свидетельствовать о том, что возрастание общего фенотипического разнообразия происходит, прежде всего, за счет увеличения уровня случайных изменений развития. Однако для некоторых признаков наблюдается независимость изменения величин фенотипического разнообразия и флуктуирующей асимметрии. Поэтому нельзя полностью исключать роль генетической составляющей в повышении общего фенотипического разнообразия. Следовательно, на основании косвенных данных частично подтверждается известный факт увеличения суммарной генетической изменчивости в совокупности подразделенных популяций (Ланде, Бэрроуклафф, 1989). Подобный результат вполне может рассматриваться как одна из форм компенсационных явлений в ответ на видовое обеднение биоты (Чернов, 2005), что наблюдается в мелких лесных резерватах. В целом полученные различия свидетельствуют о разной микроэволюционной ситуации в рассматриваемых популяциях. Ранее нами было показано, что даже очень близкие виды могут быть отличны в своих отношениях к основным характеристикам изолятов: площади, гетерогенности среды, степени изоляции от источников иммиграции (Истомин, 1984, 1990). Поэтому вполне вероятно ожидать неодинаковые направленности и скорости микроэволюционных процессов различных видов в условиях инсуляризации.
Обнаружено, что массовые ветровалы и сплошные вырубки весьма существенным и сходным образом могут изменять фенетическую структуру и популяционное разнообразие типично лесного вида рыжей полевки (рис. 11).
| Рис. 11. Дендрограмма сходства фенетической структуры популяционных группировок рыжей полевки из сравниваемых типов местообитаний |
Между группировками вывалов и вырубок существует гораздо большее сходство, чем при их сравнении с ненарушенными ельниками, несмотря на более существенные изолирующие расстояния. Степень фенетических дистанций между группировками ельников и вывалов значимо повышается в годы низкой численности. Под влиянием рассматриваемых форм нарушения зрелых лесных экосистем происходит некоторое уменьшение общей степени популяционного разнообразия или модифицируется его структура (табл. 2, 3).
Таблица 2. Феногенетическое разнообразие и скоррелированность развития признаков черепа в группировках рыжей полевки из различных местообитаний (серии 1991 г.)
Показатели феногенетического разнообразия и скоррелированность развития признаков | Ельники n=72 | Вывалы n=110 | Вырубки n=94 |
Общее число морф | 57 | 51 | 55 |
Сумма значений общего показателя разнообразия | 44,85 | 39,09 | 43,24 |
Среднее значение общего показателя разнообразия | 2,80 | 2,44 | 2,70 |
Средняя доля редких фенов | 0,198 | 0,201 | 0,195 |
Сумма значений флуктуирующей асимметрии | 177 | 281 | 293 |
Среднее значение флуктуирующей асимметрии | 11,1 | 17,6 | 18,3 |
Доля достоверных коэффициентов корреляций | 84% | 76% | 66% |
Среднее значение коэффициентов корреляций | 0,61 | 0,52 | 0,49 |
Внутрииндивидуальное разнообразие в среднем всегда достоверно выше в группировках, заселяющих вывалы и вырубки, что указывает на снижение стабильности онтогенеза и совершенства его регуляторных процессов у типичного лесного вида в дестабилизированных экосистемах, по сравнению с климаксными равновесными ельниками.
Таблица 3. Феногенетическое разнообразие группировок рыжих полевок из коренных ельников и участков сплошных вырубок (серии гг.; * - различия достоверны)
Показатели | Признаки | Ельники | Вырубки |
Показатель флуктуирующей асимметрии | Параконид М1 * | 14,3 (n=919) | 25,0 (n=44) |
Талонус М3 ** | 12,9 (n=855) | 28,6 (n=49) | |
Число морф | Параконид М1 | 6 (n=990) | 5 (n=224) |
Талонус М3 | 7 (n=878) | 7 (n=235) | |
Среднее значение общего показателя разнообразия | Параконид М1* | 4,26±0,15 | 3,60±0,23 |
Талонус М3 | 4,25±0,26 | 3,70±0,26 | |
Доля редких фенов | Параконид М1 | 0,15±0,03 | 0,14±0,06 |
Талонус М3* | 0,34±0,04 | 0,15±0,06 |
Это дополнительно подтверждается снижением скоррелированности развития линейных признаков черепа (Истомин, 2001, 2007). Несогласованность изменений внутрииндивидуального и популяционного разнообразия косвенно может свидетельствовать об определенном вкладе генетической составляющей в уменьшение общего фенотипического разнообразия популяций типично лесного вида на участках массовых ветровалов и сплошных вырубок.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
